Injury-Induced Innate Immune Response During Segment Regeneration of the Earthworm, Eisenia andrei

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
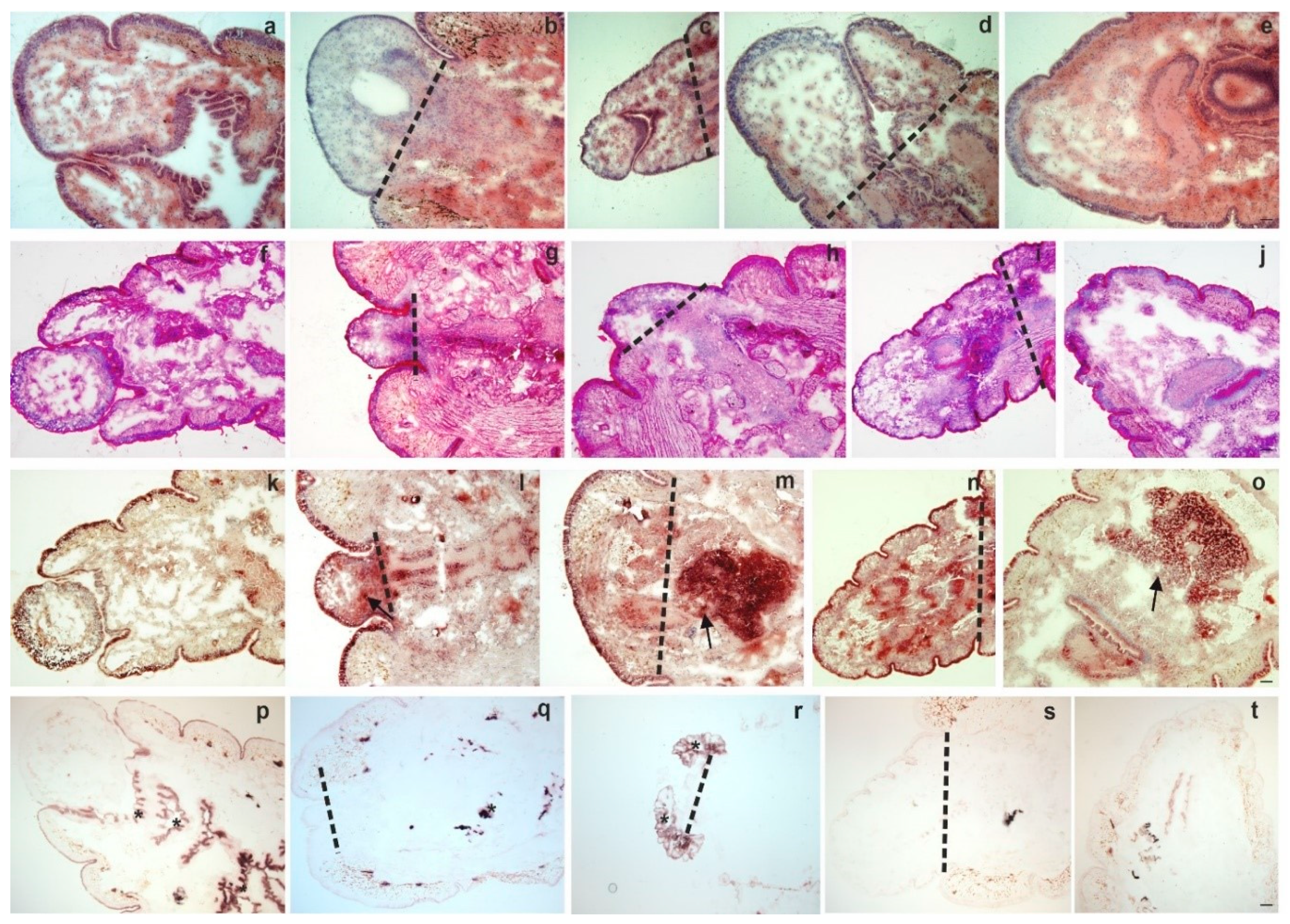
2.1. Morphological Changes During Anterior and Posterior Earthworm Regeneration
2.2. Cell Proliferation and Tissue Remodelling in the Regenerating Blastema
2.3. Coelomocytes are Accumulated in the Regenerating Blastema
2.4. Programmed Cell Death and Coelomocytes in the Course of Regeneration
2.5. Biased Immune-Related Gene Expression during Anterior and Posterior Regeneration
2.6. Lysenin Proteins are Inversely Secreted During Anterior vs. Posterior Regeneration
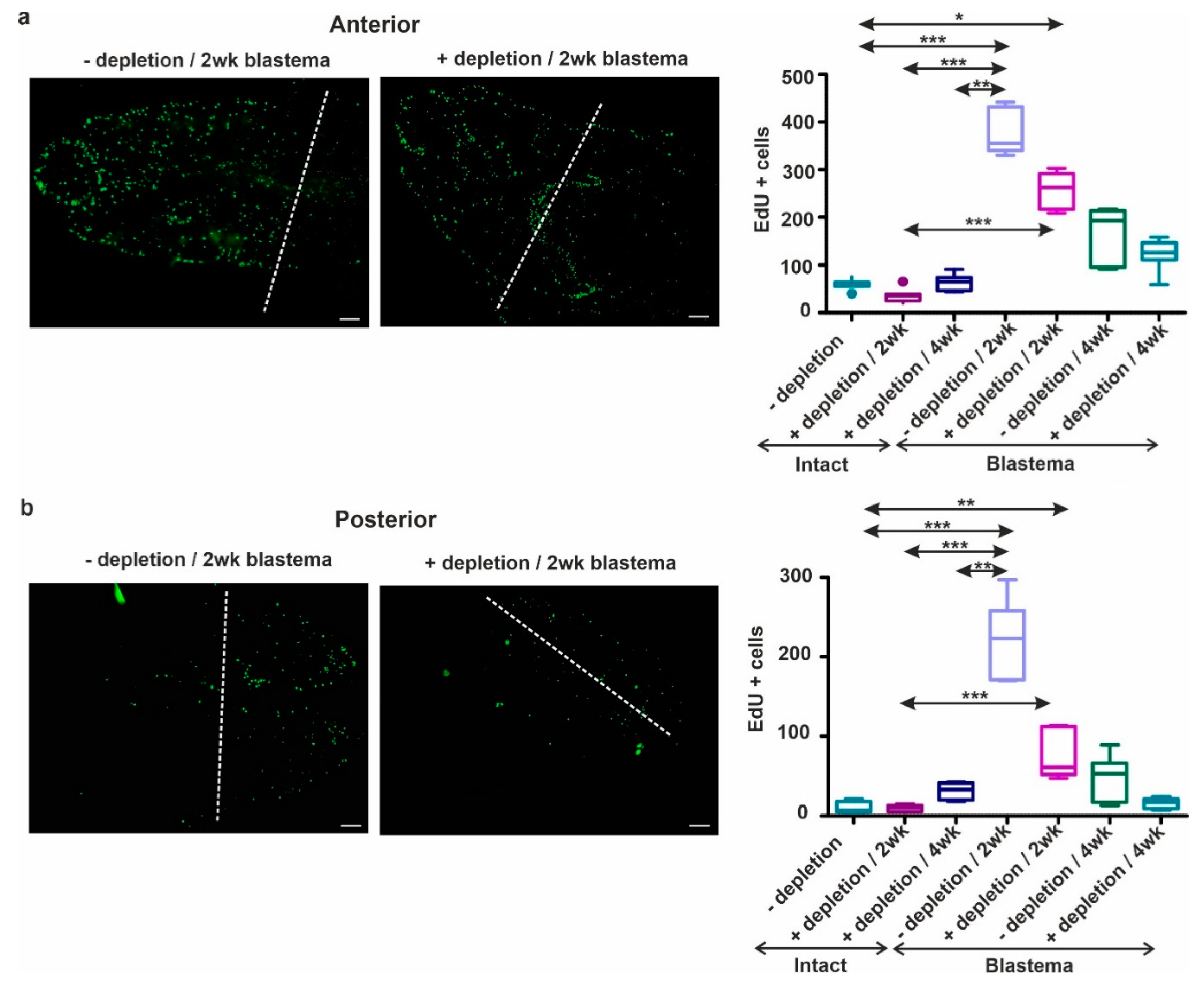
2.7. Coelomocyte Depletion Impairs Cell Proliferation and Blastema Regeneration
3. Materials and Methods
3.1. Experimental Set-Up
3.2. Detection of Cell Proliferation and Programmed Cell Death
3.3. Actin Detection
3.4. Immunofluorescence Staining of Coelomocyte Subsets
3.5. Depletion of Coelomocytes
3.6. Image Analysis
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| α-SMA | smooth muscle α actin |
| ACP | Acid phosphatase |
| ALP | Alkaline phosphatase |
| AMP | Antimicrobial peptide |
| ATP | Adenosine triphosphate |
| A.U. | Arbitrary unit |
| BSA | Bovine serum albumin |
| CC | Coelomic cavity |
| CCF | Coelomic cytolytic factor |
| DAPI | 4′6-diamino-2 phenylindole dihydrochloride |
| EdU | 5-ethynyl-2′-deoxyuridine |
| EFCC | Eisenia fetida coelomocyte clusters |
| FITC | Fluorescein isothiocyanate |
| H&E | Hematoxylin-eosin |
| H2O2 | Hydrogen peroxide |
| IL | Interleukin |
| LBSS | Lumbricus balanced salt solution |
| Lumbr | Lumbricin |
| LuRP | Lumbricin-related peptide |
| LBP/BPI | Lipopolysaccharide-binding protein/bacterial permeability-increasing protein |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| PAS | Periodic Acid and Schiff |
| PRR | Pattern recognition receptor |
| qPCR | Quantitative polymerase chain reaction |
| ROS | Reactive oxygen species |
| SDS-PAGE | Sodium dodecyl sulphate-polyacrylamide gel electrophoresis |
| SR | Scavenger receptor |
| TLR | Toll-like receptor |
| TIR | Toll/interleukin-1 receptor |
| TUNEL | Terminal deoxynucleotidyl transferase-dUTP nick end labeling |
| Wnt | Wingless/integrated |
| wk | week |
References
- Bely, A.E. Evolutionary loss of animal regeneration: Pattern and process. Integr. Comp. Biol. 2010, 50, 515–527. [Google Scholar] [CrossRef] [Green Version]
- Bely, A.E.; Nyberg, K.G. Evolution of animal regeneration: Re-emergence of a field. Trends Ecol. Evol. 2010, 25, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Mescher, A.L.; Neff, A.W. Regenerative capacity and the developing immune system. Adv. Biochem. Eng. Biotechnol. 2005, 93, 39–66. [Google Scholar] [CrossRef]
- Eming, S.A.; Krieg, T.; Davidson, J.M. Inflammation in wound repair: Molecular and cellular mechanisms. J. Investig. Dermatol. 2007, 127, 514–525. [Google Scholar] [CrossRef] [Green Version]
- Engelmann, P.; Bodó, K.; Najbauer, J.; Németh, P. Annelida: Oligochaetes (Segmented Worms): Earthworm immunity, quo vadis? Advances and new paradigms in the omics era. In Advances in Comparative Immunology; Cooper, E.L., Ed.; Springer International Publishing: Heidelberg, Germany, 2018; pp. 135–159. [Google Scholar]
- Julier, Z.; Park, A.J.; Briquez, P.S.; Martino, M.M. Promoting tissue regeneration by modulating the immune system. Acta Biomater. 2017, 53, 13–28. [Google Scholar] [CrossRef]
- Eming, S.A. Evolution of immune pathways in regeneration and repair: Recent concepts and translational perspectives. Semin. Immunol. 2014, 26, 275–276. [Google Scholar] [CrossRef]
- Bely, A.E.; Sikes, J.M. Latent regeneration abilities persist following recent evolutionary loss in asexual annelids. Proc. Natl. Acad. Sci. USA 2010, 107, 1464–1469. [Google Scholar] [CrossRef] [Green Version]
- Baranzini, N.; Pulze, L.; Acquati, F.; Grimaldi, A. Hirudo verbena as an alternative model to dissect the relationship between innate immunity and regeneration. Invertebr. Surv. J. 2020, 17, 90–98. [Google Scholar] [CrossRef]
- Bely, A.E. Distribution of segment regeneration ability in the Annelida. Integr. Comp. Biol. 2006, 46, 508–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varhalmi, E.; Somogyi, I.; Kiszler, G.; Nemeth, J.; Reglodi, D.; Lubics, A.; Kiss, P.; Tamas, A.; Pollak, E.; Molnar, L. Expression of PACAP-like compounds during the caudal regeneration of the earthworm Eisenia fetida. J. Mol. Neurosci. 2008, 36, 166–174. [Google Scholar] [CrossRef]
- Tessmar-Raible, K.; Arendt, D. Emerging systems: Between vertebrates and arthropods, the Lophotrochozoa. Curr. Opin. Genet. Dev. 2003, 13, 331–340. [Google Scholar] [CrossRef]
- Nyberg, K.G.; Conte, M.A.; Kostyun, J.L.; Forde, A.; Bely, A.E. Transcriptome characterization via 454 pyrosequencing of the annelid Pristina leidyi, an emerging model for studying the evolution of regeneration. BMC Genom. 2012, 13, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, E.L.; Kauschke, E.; Cossarizza, A. Digging for innate immunity since Darwin and Metchnikoff. Bioessays 2002, 24, 319–333. [Google Scholar] [CrossRef]
- Engelmann, P.; Hayashi, Y.; Bodó, K.; Ernszt, D.; Somogyi, I.; Steib, A.; Orbán, J.; Pollák, E.; Nyitrai, M.; Németh, P.; et al. Phenotypic and functional characterization of earthworm coelomocyte subsets: Linking light scatter-based cell typing and imaging of the sorted populations. Dev. Comp. Immunol. 2016, 65, 41–52. [Google Scholar] [CrossRef]
- Engelmann, P.; Hayashi, Y.; Bodó, K.; Molnár, L. New aspects of earthworm innate immunity: Novel molecules and old proteins with unexpected functions. In Lessons in Immunity: From Single Cell Organisms to Mammals; Ballarin, L., Cammarata, M., Eds.; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2016; pp. 53–66. [Google Scholar]
- Engelmann, P.; Pálinkás, L.; Cooper, E.L.; Németh, P. Monoclonal antibodies identify four distinct annelid leukocyte markers. Dev. Comp. Immunol. 2005, 29, 599–614. [Google Scholar] [CrossRef] [PubMed]
- Bodó, K.; Ernszt, D.; Németh, P.; Engelmann, P. Distinct immune- and defense-related molecular fingerprints in separated coelomocyte subsets in Eisenia andrei earthworms. Invertebr. Surv. J. 2018, 15, 338–345. [Google Scholar] [CrossRef]
- Prochazkova, P.; Roubalova, R.; Dvorak, J.; Navarro Pacheco, N.I.; Bilej, M. Pattern recognition receptors in annelids. Dev. Comp. Immunol. 2020, 102, 103493. [Google Scholar] [CrossRef]
- Bhambri, A.; Dhaunta, N.; Patel, S.S.; Hardikar, M.; Bhatt, A.; Srikakulam, N.; Shridhar, S.; Vellarikkal, S.; Pandey, R.; Jayarajan, R.; et al. Large scale changes in the transcriptome of Eisenia fetida during regeneration. PLoS ONE 2018, 13, e0204234. [Google Scholar] [CrossRef]
- He, C.; Klionsky, D.J. Regulation mechanisms and signaling pathways of autophagy. Annu. Rev. Genet. 2009, 43, 67–93. [Google Scholar] [CrossRef] [Green Version]
- Kwong, W.H.; Tam, P.P. The pattern of alkaline phosphatase activity in the developing mouse spinal cord. J. Embryol. Exp. Morphol. 1984, 82, 241–251. [Google Scholar]
- Szabó, R.; Ferrier, D.E. The dynamics of alkaline phosphatase activity during operculum regeneration in the polychaete Pomatoceros lamarckii. Int. J. Dev. Biol. 2014, 58, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Rockey, D.C.; Weymouth, N.; Shi, Z. Smooth muscle α actin (Acta2) and myofibroblast function during hepatic wound healing. PLoS ONE 2013, 8, e77166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darby, I.A.; Laverdet, B.; Bonté, F.; Desmoulière, A. Fibroblasts and myofibroblasts in wound healing. Clin. Cosmet. Investig. Dermatol. 2014, 7, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Abnave, P.; Ghigo, E. Role of the immune system in regeneration and its dynamic interplay with adult stem cells. Semin. Cell Dev. Biol. 2019, 87, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Liebmann, E. New light on regeneration of Eisenia foetida (SAV). J. Morphol. 1943, 73, 583–610. [Google Scholar] [CrossRef]
- Cooper, E.L.; Roch, P. Earthworm leukocyte interactions during early stages of graft rejection. J. Exp. Zool. 1984, 232, 67–72. [Google Scholar] [CrossRef]
- Chaves da Silva, P.G.; Corrêa, C.L.; de Carvalho, S.L.; Allodi, S. The crustacean central nervous system in focus: Subacute neurodegeneration induces a specific innate immune response. PLoS ONE 2013, 20, e80896. [Google Scholar] [CrossRef] [Green Version]
- Schikorski, D.; Cuvillier-Hot, V.; Leippe, M.; Boidin-Wichlacz, C.; Slomianny, C.; Macagno, E.; Salzet, M.; Tasiemski, A. Microbial challenge promotes the regenerative process of the injured central nervous system of the medicinal leech by inducing the synthesis of antimicrobial peptides in neurons and microglia. J. Immunol. 2008, 181, 1083–1095. [Google Scholar] [CrossRef] [Green Version]
- Vriz, S.; Reiter, S.; Galliot, B. Cell death: A program to regenerate. Curr. Top. Dev. Biol. 2014, 108, 121–151. [Google Scholar] [CrossRef] [Green Version]
- Galliot, B. Injury-induced asymmetric cell death as a driving force for head regeneration in Hydra. Dev. Genes Evol. 2013, 223, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.S.; Kobayashi, C.; Agata, K.; Ikeo, K.; Gojobori, T. Detection of apoptosis during planarian regeneration by the expression of apoptosis-related genes and TUNEL assay. Gene 2004, 333, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Holstein, T.W.; Watanabe, H.; Ozbek, S. Signaling pathways and axis formation in the lower metazoa. Curr. Top. Dev. Biol. 2011, 97, 137–177. [Google Scholar] [CrossRef] [PubMed]
- Gauron, C.; Rampon, C.; Bouzaffour, M.; Ipendey, E.; Teillon, J.; Volovitch, M.; Vriz, S. Sustained production of ROS triggers compensatory proliferation and is required for regeneration to proceed. Sci. Rep. 2013, 3, 2084. [Google Scholar] [CrossRef] [Green Version]
- Willenborg, S.; Eming, S.A. Macrophages-sensors and effectors coordinating skin damage and repair. J. Dtsch Dermatol. Ges. 2014, 12, 214–221. [Google Scholar] [CrossRef]
- Morales, R.A.; Allende, M.L. Peripheral macrophages promote tissue regeneration in zebrafish by fine-tuning the inflammatory response. Front. Immunol. 2019, 10, 253. [Google Scholar] [CrossRef] [Green Version]
- Okrzesik, J.; Kachamakova-Trojanowska, N.; Jozkowicz, A.; Morgan, A.J.; Plytycz, B. Reversible inhibition of reproduction during regeneration of cerebral ganglia and coelomocytes in the earthworm Dendrobaena veneta. Invertebr. Surv. J. 2013, 10, 151–161. [Google Scholar]
- Molnar, L.; Pollak, E.; Skopek, Z.; Gutt, E.; Kruk, J.; Morgan, A.J.; Plytycz, B. Immune system participates in brain regeneration and restoration of reproduction in the earthworm Dendrobaena veneta. Dev. Comp. Immunol. 2015, 52, 269–279. [Google Scholar] [CrossRef]
- Tadokoro, R.; Sugio, M.; Kutsuna, J.; Tochinai, S.; Takahashi, Y. Early segregation of germ and somatic lineages during gonadal regeneration in the annelid Enchytraeus japonensis. Curr. Biol. 2006, 16, 1012–1017. [Google Scholar] [CrossRef] [Green Version]
- Myohara, M.; Niva, C.C.; Lee, J.M. Molecular approach to annelid regeneration: cDNA subtraction cloning reveals various novel genes that are upregulated during the large-scale regeneration of the oligochaete, Enchytraeus japonensis. Dev. Dyn. 2006, 235, 2051–2070. [Google Scholar] [CrossRef] [PubMed]
- Niva, C.C.; Lee, J.M.; Myohara, M. Glutamine synthetase gene expression during the regeneration of the annelid Enchytraeus japonensis. Dev. Genes Evol. 2008, 218, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeo, M.; Yoshida-Noro, C.; Tochinai, S. Functional analysis of grimp, a novel gene required for mesodermal cell proliferation at an initial stage of regeneration in Enchytraeus japonensis (Enchytraeidae, Oligochaete). Int. J. Dev. Biol. 2010, 54, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Pfeifer, K.; Dorresteijn, A.W.; Fröbius, A.C. Activation of Hox genes during caudal regeneration of the polychaete annelid Platynereis dumerilii. Dev. Genes Evol. 2012, 222, 165–179. [Google Scholar] [CrossRef]
- Pfeifer, K.; Schaub, C.; Wolfstetter, G.; Dorresteijn, A. Identification and characterization of a twist ortholog in the polychaete annelid Platynereis dumerilii reveals mesodermal expression of Pdu-twist. Dev. Genes Evol. 2013, 223, 319–328. [Google Scholar] [CrossRef]
- Huang, X.M.; Tian, Q.N.; Bao, Z.X.; Qin, Y.F.; Chen, S.J.; Lu, P.; Zhang, X.L.; Zhang, Y.Z.; Zhang, S.T. Cloning and identification of microRNAs in earthworm (Eisenia fetida). Biochem Genet. 2012, 50, 1–11. [Google Scholar] [CrossRef]
- Novikova, E.L.; Bakalenko, N.I.; Nesterenko, A.Y.; Kulakova, M.A. Expression of Hox genes during regeneration of nereid polychaete Alitta (Nereis) virens (Annelida, Lophotrochozoa). Evodevo 2013, 4, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özpolat, B.D.; Bely, A.E. Gonad establishment during asexual reproduction in the annelid Pristina leidyi. Dev. Biol. 2015, 405, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Kozin, V.V.; Kostyuchenko, R.P. Vasa, PL10, and Piwi gene expression during caudal regeneration of the polychaete annelid Alitta virens. Dev. Genes Evol. 2015, 225, 129–138. [Google Scholar] [CrossRef]
- De Jong, D.M.; Seaver, E.C. A Stable Thoracic Hox Code and Epimorphosis Characterize Posterior Regeneration in Capitella teleta. PLoS ONE 2016, 11, e0149724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Škanta, F.; Procházková, P.; Roubalová, R.; Dvořák, J.; Bilej, M. LBP/BPI homologue in Eisenia andrei earthworms. Dev. Comp. Immunol. 2016, 54, 1–6. [Google Scholar] [CrossRef]
- Pearson, A.M. Scavenger receptors in innate immunity. Curr. Opin. Immunol. 1996, 8, 20–28. [Google Scholar] [CrossRef]
- Brennan, J.J.; Gilmore, T.D. Evolutionary origins of toll-like receptor signaling. Mol. Biol. Evol. 2018, 35, 1576–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Škanta, F.; Roubalová, R.; Dvořák, J.; Procházková, P.; Bilej, M. Molecular cloning and expression of TLR in the Eisenia andrei earthworm. Dev. Comp. Immunol. 2013, 41, 694–702. [Google Scholar] [CrossRef]
- Chen, L.; DiPietro, L.A. Toll-Like receptor function in acute wounds. Adv. Wound Care 2017, 6, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, S.; Aggarwal, K.; Paquette, N.; Silverman, N. NF-κB/Rel proteins and the humoral immune responses of Drosophila melanogaster. Curr. Top. Microbiol Immunol. 2011, 349, 25–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canton, J.; Neculai, D.; Grinstein, S. Scavenger receptors in homeostasis and immunity. Nat. Rev. Immunol. 2013, 13, 621–634. [Google Scholar] [CrossRef]
- Bodó, K.; Boros, Á.; Rumpler, É.; Molnár, L.; Böröcz, K.; Németh, P.; Engelmann, P. Identification of novel lumbricin homologues in Eisenia andrei earthworms. Dev. Comp. Immunol. 2019, 90, 41–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opper, B.; Bognár, A.; Heidt, D.; Németh, P.; Engelmann, P. Revising lysenin expression of earthworm coelomocytes. Dev. Comp. Immunol. 2013, 39, 214–218. [Google Scholar] [CrossRef]
- Wilkinson, H.N.; Hardman, M.J. Wound healing: Cellular mechanisms and pathological outcomes. Open Biol. 2020, 10, 200223. [Google Scholar] [CrossRef]
- Wenger, Y.; Buzgariu, W.; Reiter, S.; Galliot, B. Injury-induced immune responses in Hydra. Semin. Immunol. 2014, 26, 277–294. [Google Scholar] [CrossRef]
- Peiris, T.H.; Hoyer, K.K.; Oviedo, N.J. Innate immune system and tissue regeneration in planarians: An area ripe for exploration. Semin. Immunol. 2014, 26, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitulo, N.; Dalla Valle, L.; Skobo, T.; Valle, G.; Alibardi, L. Downregulation of lizard immuno-genes in the regenerating tail and myogenes in the scarring limb suggests that tail regeneration occurs in an immuno-privileged organ. Protoplasma 2017, 254, 2127–2141. [Google Scholar] [CrossRef] [PubMed]
- Godwin, J.W.; Pinto, A.R.; Rosenthal, N.A. Macrophages are required for adult salamander limb regeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 9415–9420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, M.W.; Neff, A.W.; Mescher, A.L. The developing Xenopus limb as a model for studies on the balance between inflammation and regeneration. Anat. Rec. 2012, 295, 1552–1561. [Google Scholar] [CrossRef]
- Zattara, E.E.; Bely, A.E. Investment choices in post-embryonic development: Quantifying interactions among growth, regeneration, and asexual reproduction in the annelid Pristina leidyi. J. Exp. Zool. B. Mol. Dev. Evol. 2013, 320, 471–488. [Google Scholar] [CrossRef] [PubMed]
- Bodó, K.; Hayashi, Y.; Gerencsér, G.; László, Z.; Kéri, A.; Galbács, G.; Telek, E.; Mészáros, M.; Deli, A.M.; Kokhanyuk, B.; et al. Species-specific sensitivity of Eisenia earthworms towards nobel metal nanoparticles: A multiparametric in vitro study. Environ. Sci. Nano 2020, 7, 3509–3525. [Google Scholar] [CrossRef]
- Mangoni, M.L.; McDermott, A.M.; Zasloff, M. Antimicrobial peptides and wound healing: Biological and therapeutic considerations. Exp. Dermatol. 2016, 25, 167–173. [Google Scholar] [CrossRef]
- Eming, S.A.; Wynn, T.A.; Martin, P. Inflammation and metabolism in tissue repair and regeneration. Science 2017, 356, 1026–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen-Chi, M.; Laplace-Builhe, B.; Travnickova, J.; Luz-Crawford, P.; Tejedor, G.; Phan, Q.T.; Duroux-Richard, I.; Levraud, J.P.; Kissa, K.; Lutfalla, G.; et al. Identification of polarized macrophage subsets in zebrafish. elife 2015, 4, e07288. [Google Scholar] [CrossRef]
- Nguyen-Chi, M.; Laplace-Builhé, B.; Travnickova, J.; Luz-Crawford, P.; Tejedor, G.; Lutfalla, G.; Kissa, K.; Jorgensen, C.; Djouad, F. TNF signaling and macrophages govern fin regeneration in zebrafish larvae. Cell Death Dis. 2017, 8, e2979. [Google Scholar] [CrossRef]
- Molnár, L.; Engelmann, P.; Somogyi, I.; Mácsik, L.L.; Pollák, E. Cold-stress induced formation of calcium and phosphorous rich chloragocyte granules (chloragosomes) in the earthworm Eisenia fetida. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2012, 163, 199–209. [Google Scholar] [CrossRef]
- Kellermayer, Z.; Vojkovics, D.; Dakah, T.A.; Bodó, K.; Botz, B.; Helyes, Z.; Berta, G.; Kajtár, B.; Schippers, A.; Wagner, N.; et al. IL-22-independent protection from colitis in the absence of Nkx2.3 transcription factor in mice. J. Immunol. 2019, 202, 1833–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelmann, P.; Molnár, L.; Pálinkás, L.; Cooper, E.L.; Németh, P. Earthworm leukocyte populations specifically harbor lysosomal enzymes that may respond to bacterial challenge. Cell Tissue Res. 2004, 316, 391–401. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bodó, K.; Kellermayer, Z.; László, Z.; Boros, Á.; Kokhanyuk, B.; Németh, P.; Engelmann, P. Injury-Induced Innate Immune Response During Segment Regeneration of the Earthworm, Eisenia andrei. Int. J. Mol. Sci. 2021, 22, 2363. https://doi.org/10.3390/ijms22052363
Bodó K, Kellermayer Z, László Z, Boros Á, Kokhanyuk B, Németh P, Engelmann P. Injury-Induced Innate Immune Response During Segment Regeneration of the Earthworm, Eisenia andrei. International Journal of Molecular Sciences. 2021; 22(5):2363. https://doi.org/10.3390/ijms22052363
Chicago/Turabian StyleBodó, Kornélia, Zoltán Kellermayer, Zoltán László, Ákos Boros, Bohdana Kokhanyuk, Péter Németh, and Péter Engelmann. 2021. "Injury-Induced Innate Immune Response During Segment Regeneration of the Earthworm, Eisenia andrei" International Journal of Molecular Sciences 22, no. 5: 2363. https://doi.org/10.3390/ijms22052363
APA StyleBodó, K., Kellermayer, Z., László, Z., Boros, Á., Kokhanyuk, B., Németh, P., & Engelmann, P. (2021). Injury-Induced Innate Immune Response During Segment Regeneration of the Earthworm, Eisenia andrei. International Journal of Molecular Sciences, 22(5), 2363. https://doi.org/10.3390/ijms22052363