
Gap Junctional Communication via Connexin43 between Purkinje Fibers and Working Myocytes Explains the Epicardial Activation Pattern in the Postnatal Mouse Left Ventricle


 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
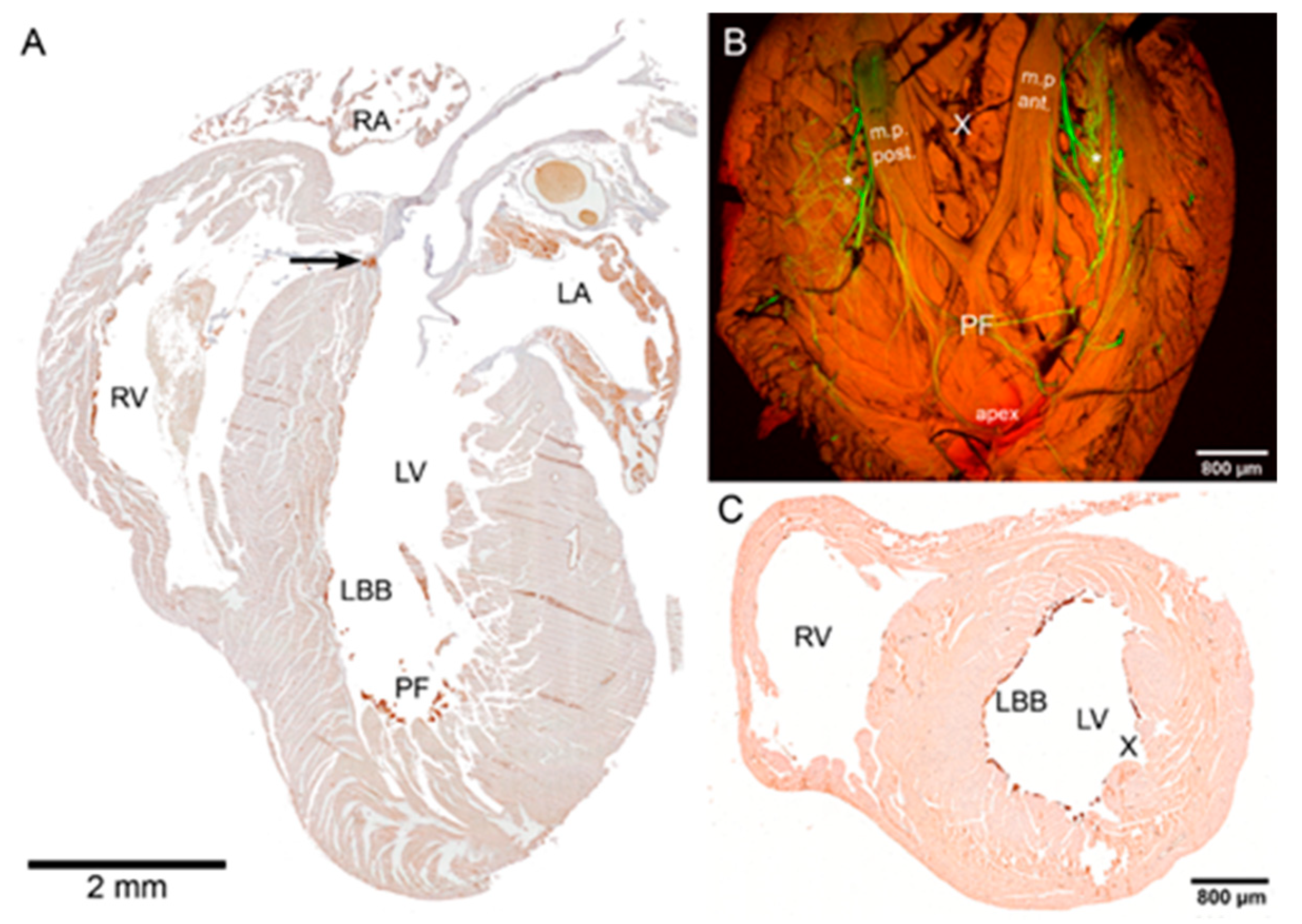
2.1. Cx43 Expression in Purkinje-Myocyte Junctions
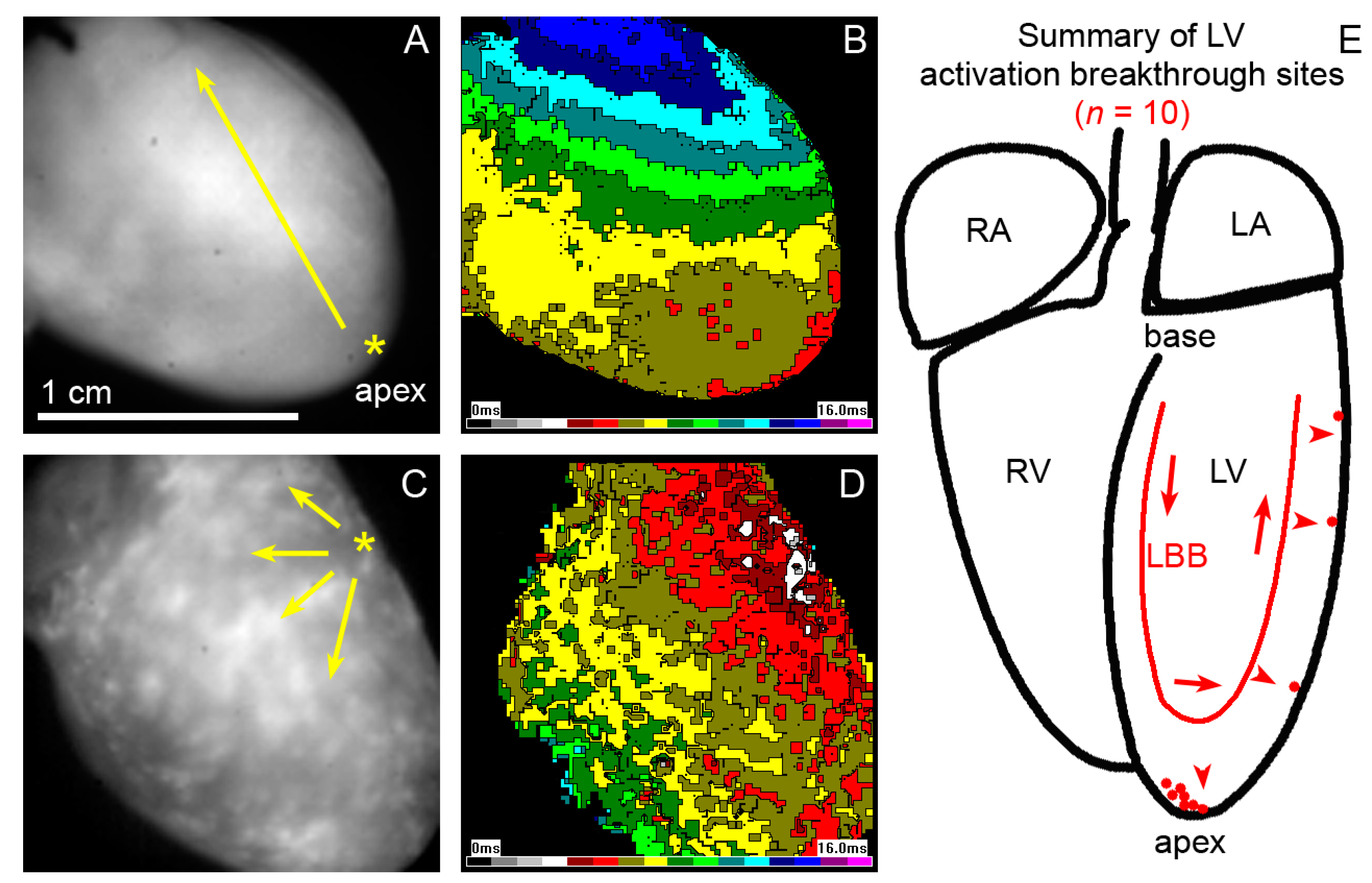
2.2. Epicardial Activation Patterns of the Left Ventricle
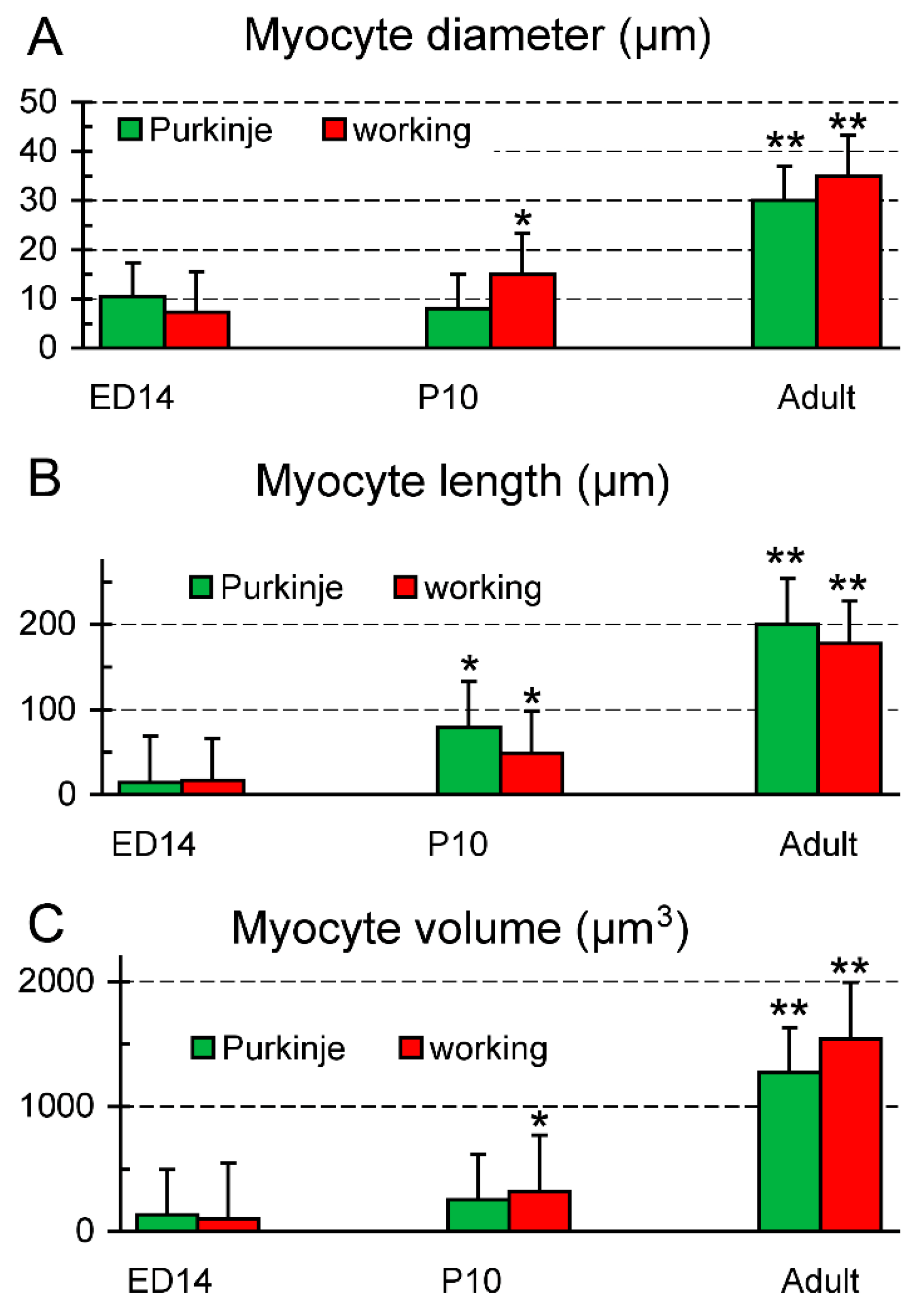
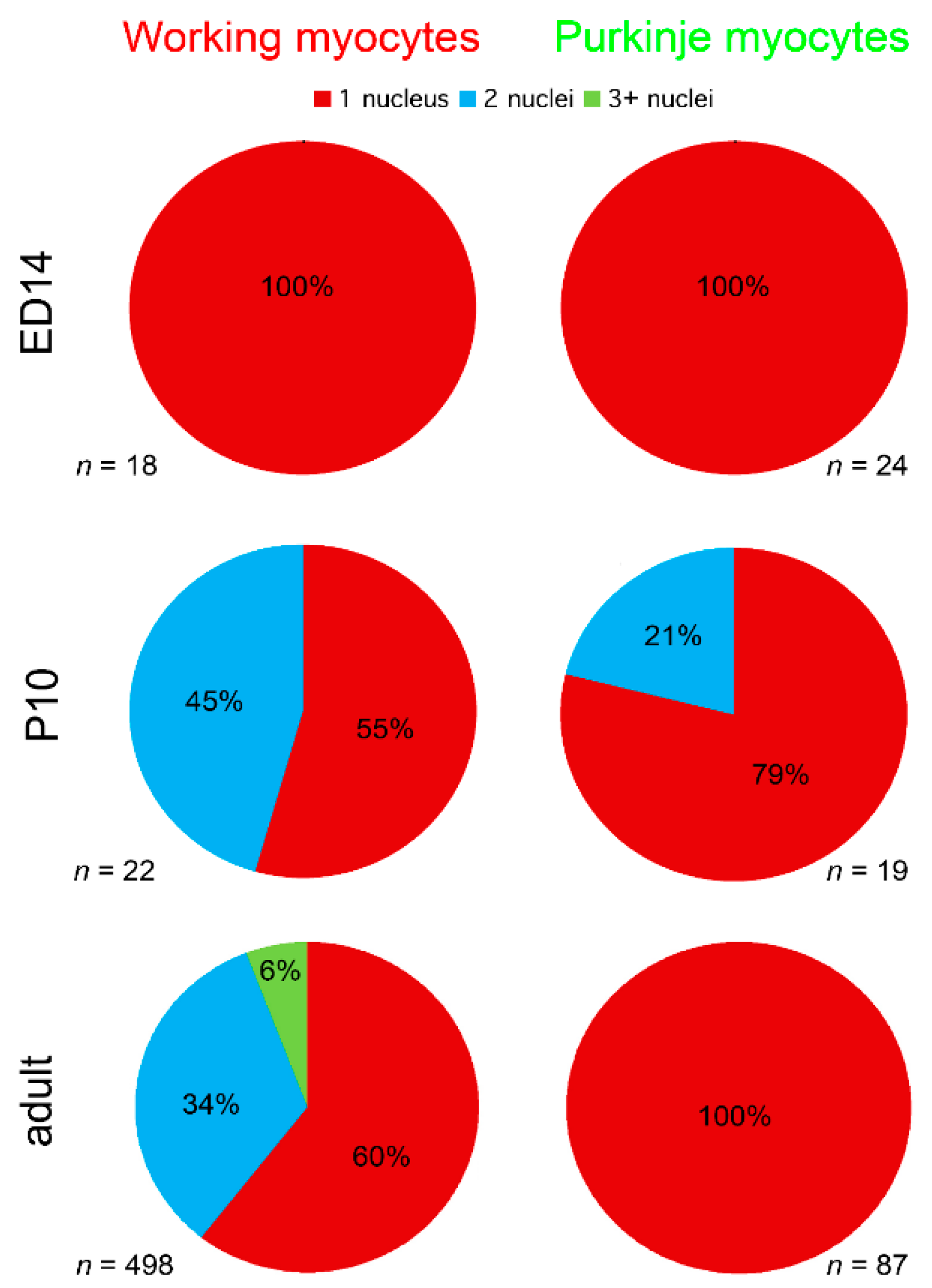
2.3. Cellular Analysis of the PF Network
3. Discussion
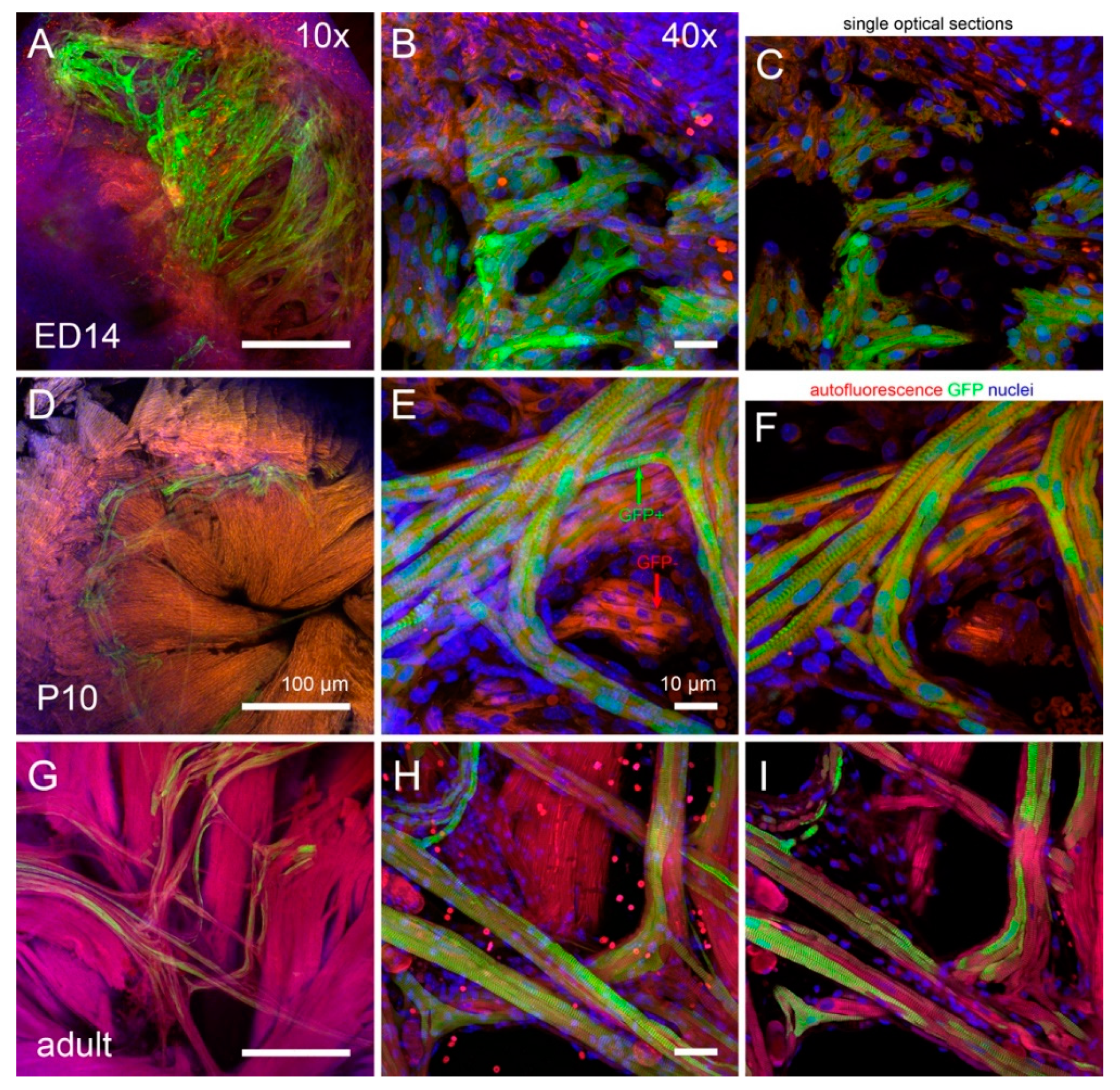
3.1. Maturation of Cx43 Expression
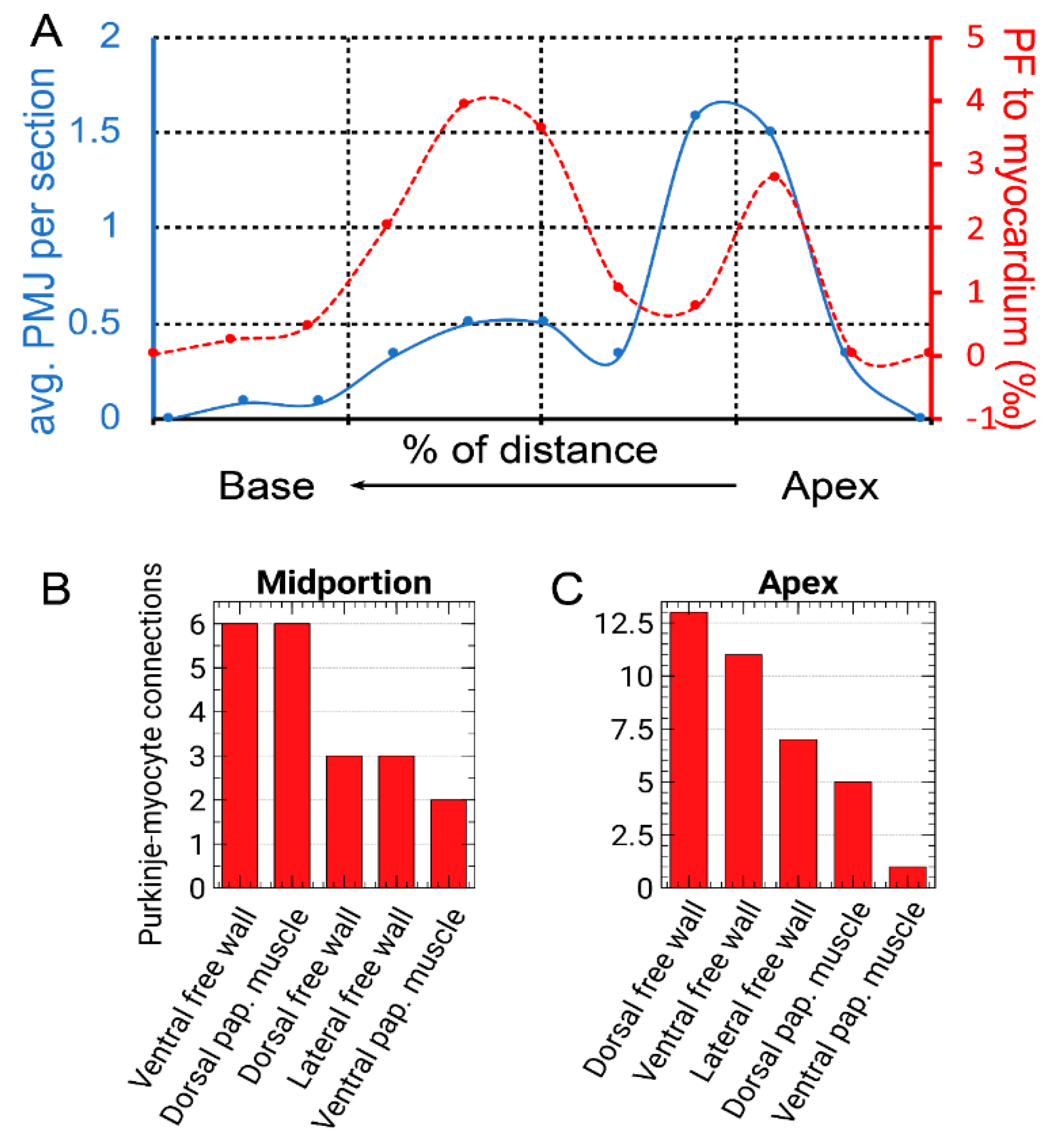
3.2. Distribution of Purkinje-Myocyte Junctions and Its Correlation with the Ventricular Activation Pattern
3.3. Morphological Characteristics of the Purkinje Network
3.4. Conclusions
3.5. Limitations, Future Directions, Perspectives
4. Materials and Methods
4.1. Heart Sampling and Processing
4.2. Quantitative Analysis
4.3. Optical Mapping
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Volgyi, B.; Kovacs-Oller, T.; Atlasz, T.; Wilhelm, M.; Gabriel, R. Gap junctional coupling in the vertebrate retina: Variations on one theme? Prog. Retin. Eye Res. 2013, 34, 1–18. [Google Scholar] [CrossRef]
- Van Kempen, M.J.; Fromaget, C.; Gros, D.; Moorman, A.F.; Lamers, W.H. Spatial distribution of connexin43, the major cardiac gap junction protein, in the developing and adult rat heart. Circ. Res. 1991, 68, 1638–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoagland, D.T.; Santos, W.; Poelzing, S.; Gourdie, R.G. The role of the gap junction perinexus in cardiac conduction: Potential as a novel anti-arrhythmic drug target. Prog. Biophys. Mol. Biol. 2019, 144, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Ek-Vitorin, J.F.; Taffet, S.M.; Delmar, M. Coexpression of connexins 40 and 43 enhances the pH sensitivity of gap junctions: A model for synergistic interactions among connexins. Circ. Res. 2000, 86, E98–E103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rackauskas, M.; Kreuzberg, M.M.; Pranevicius, M.; Willecke, K.; Verselis, V.K.; Bukauskas, F.F. Gating properties of heterotypic gap junction channels formed of connexins 40, 43, and 45. Biophys. J. 2007, 92, 1952–1965. [Google Scholar] [CrossRef] [Green Version]
- Rackauskas, M.; Verselis, V.K.; Bukauskas, F.F. Permeability of homotypic and heterotypic gap junction channels formed of cardiac connexins mCx30.2, Cx40, Cx43, and Cx45. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1729–H1736. [Google Scholar] [CrossRef] [PubMed]
- Desplantez, T. Cardiac Cx43, Cx40 and Cx45 co-assembling: Involvement of connexins epitopes in formation of hemichannels and Gap junction channels. BMC Cell Biol. 2017, 18 (Suppl. 1), 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ek-Vitorin, J.F.; King, T.J.; Heyman, N.S.; Lampe, P.D.; Burt, J.M. Selectivity of connexin 43 channels is regulated through protein kinase C-dependent phosphorylation. Circ. Res. 2006, 98, 1498–1505. [Google Scholar] [CrossRef] [Green Version]
- Solan, J.L.; Lampe, P.D. Spatio-temporal regulation of connexin43 phosphorylation and gap junction dynamics. Biochim. Biophys. Acta Biomembr. 2018, 1860, 83–90. [Google Scholar] [CrossRef]
- Van Veen, A.A.; van Rijen, H.V.; Opthof, T. Cardiac gap junction channels: Modulation of expression and channel properties. Cardiovasc. Res. 2001, 51, 217–229. [Google Scholar] [CrossRef]
- Axelsen, L.N.; Calloe, K.; Holstein-Rathlou, N.H.; Nielsen, M.S. Managing the complexity of communication: Regulation of gap junctions by post-translational modification. Front. Pharm. 2013, 4, 130. [Google Scholar] [CrossRef] [Green Version]
- Axelsen, L.N.; Stahlhut, M.; Mohammed, S.; Larsen, B.D.; Nielsen, M.S.; Holstein-Rathlou, N.H.; Andersen, S.; Jensen, O.N.; Hennan, J.K.; Kjolbye, A.L. Identification of ischemia-regulated phosphorylation sites in connexin43: A possible target for the antiarrhythmic peptide analogue rotigaptide (ZP123). J. Mol. Cell. Cardiol. 2006, 40, 790–798. [Google Scholar] [CrossRef]
- Kohutova, J.; Elsnicova, B.; Holzerova, K.; Neckar, J.; Sebesta, O.; Jezkova, J.; Vecka, M.; Vebr, P.; Hornikova, D.; Szeiffova Bacova, B.; et al. Anti-arrhythmic Cardiac Phenotype Elicited by Chronic Intermittent Hypoxia Is Associated With Alterations in Connexin-43 Expression, Phosphorylation, and Distribution. Front. Endocrinol. 2018, 9, 789. [Google Scholar] [CrossRef]
- Sedmera, D.; Neckar, J.; Benes, J., Jr.; Pospisilova, J.; Petrak, J.; Sedlacek, K.; Melenovsky, V. Changes in Myocardial Composition and Conduction Properties in Rat Heart Failure Model Induced by Chronic Volume Overload. Front. Physiol. 2016, 7, 367. [Google Scholar] [CrossRef] [Green Version]
- Remo, B.F.; Qu, J.; Volpicelli, F.M.; Giovannone, S.; Shin, D.; Lader, J.; Liu, F.Y.; Zhang, J.; Lent, D.S.; Morley, G.E.; et al. Phosphatase-resistant gap junctions inhibit pathological remodeling and prevent arrhythmias. Circ. Res. 2011, 108, 1459–1466. [Google Scholar] [CrossRef]
- Howarth, F.C.; Chandler, N.J.; Kharche, S.; Tellez, J.O.; Greener, I.D.; Yamanushi, T.T.; Billeter, R.; Boyett, M.R.; Zhang, H.; Dobrzynski, H. Effects of streptozotocin-induced diabetes on connexin43 mRNA and protein expression in ventricular muscle. Mol. Cell Biochem. 2008, 319, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Ogawa, K.; Imanaga, I.; Tribulova, N. Remodeling of connexin 43 in the diabetic rat heart. Mol. Cell Biochem. 2006, 290, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Boulaksil, M.; Winckels, S.K.; Engelen, M.A.; Stein, M.; van Veen, T.A.; Jansen, J.A.; Linnenbank, A.C.; Bierhuizen, M.F.; Groenewegen, W.A.; van Oosterhout, M.F.; et al. Heterogeneous Connexin43 distribution in heart failure is associated with dispersed conduction and enhanced susceptibility to ventricular arrhythmias. Eur. J. Heart Fail. 2010, 12, 913–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostin, S.; Rieger, M.; Dammer, S.; Hein, S.; Richter, M.; Klovekorn, W.P.; Bauer, E.P.; Schaper, J. Gap junction remodeling and altered connexin43 expression in the failing human heart. Mol. Cell Biochem. 2003, 242, 135–144. [Google Scholar] [CrossRef]
- Andelova, K.; Egan Benova, T.; Szeiffova Bacova, B.; Sykora, M.; Prado, N.J.; Diez, E.R.; Hlivak, P.; Tribulova, N. Cardiac Connexin-43 Hemichannels and Pannexin1 Channels: Provocative Antiarrhythmic Targets. Int. J. Mol. Sci. 2020, 22, 260. [Google Scholar] [CrossRef] [PubMed]
- Van der Velden, H.M.; van Kempen, M.J.; Wijffels, M.C.; van Zijverden, M.; Groenewegen, W.A.; Allessie, M.A.; Jongsma, H.J. Altered pattern of connexin40 distribution in persistent atrial fibrillation in the goat. J. Cardiovasc. Electrophysiol. 1998, 9, 596–607. [Google Scholar] [CrossRef]
- Van der Velden, H.M.; Jongsma, H.J. Cardiac gap junctions and connexins: Their role in atrial fibrillation and potential as therapeutic targets. Cardiovasc. Res. 2002, 54, 270–279. [Google Scholar] [CrossRef]
- Franco, D.; Icardo, J.M. Molecular characterization of the ventricular conduction system in the developing mouse heart: Topographical correlation in normal and congenitally malformed hearts. Cardiovasc. Res. 2001, 49, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Gourdie, R.G.; Green, C.R.; Severs, N.J.; Anderson, R.H.; Thompson, R.P. Evidence for a distinct gap-junctional phenotype in ventricular conduction tissues of the developing and mature avian heart. Circ. Res. 1993, 72, 278–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Severs, N.J.; Bruce, A.F.; Dupont, E.; Rothery, S. Remodelling of gap junctions and connexin expression in diseased myocardium. Cardiovasc. Res. 2008, 80, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Coppen, S.R.; Severs, N.J.; Gourdie, R.G. Connexin45 (alpha 6) expression delineates an extended conduction system in the embryonic and mature rodent heart. Dev. Genet. 1999, 24, 82–90. [Google Scholar] [CrossRef]
- Kumai, M.; Nishii, K.; Nakamura, K.; Takeda, N.; Suzuki, M.; Shibata, Y. Loss of connexin45 causes a cushion defect in early cardiogenesis. Development 2000, 127, 3501–3512. [Google Scholar] [PubMed]
- Gros, D.; Theveniau-Ruissy, M.; Bernard, M.; Calmels, T.; Kober, F.; Sohl, G.; Willecke, K.; Nargeot, J.; Jongsma, H.J.; Mangoni, M.E. Connexin 30 is expressed in the mouse sino-atrial node and modulates heart rate. Cardiovasc. Res. 2010, 85, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Van Weerd, J.H.; Christoffels, V.M. The formation and function of the cardiac conduction system. Development 2016, 143, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Benes, J., Jr.; Ammirabile, G.; Sankova, B.; Campione, M.; Krejci, E.; Kvasilova, A.; Sedmera, D. The role of connexin40 in developing atrial conduction. FEBS Lett. 2014, 588, 1465–1469. [Google Scholar] [CrossRef] [Green Version]
- Becker, D.L.; Cook, J.E.; Davies, C.S.; Evans, W.H.; Gourdie, R.G. Expression of major gap junction connexin types in the working myocardium of eight chordates. Cell Biol. Int. 1998, 22, 527–543. [Google Scholar] [CrossRef] [PubMed]
- Ai, X.; Pogwizd, S.M. Connexin 43 downregulation and dephosphorylation in nonischemic heart failure is associated with enhanced colocalized protein phosphatase type 2A. Circ. Res. 2005, 96, 54–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akar, F.G.; Nass, R.D.; Hahn, S.; Cingolani, E.; Shah, M.; Hesketh, G.G.; DiSilvestre, D.; Tunin, R.S.; Kass, D.A.; Tomaselli, G.F. Dynamic changes in conduction velocity and gap junction properties during development of pacing-induced heart failure. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1223–H1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, B.; Boukens, B.J.; Crossley, D.A., 2nd; Conner, J.; Mohan, R.A.; van Duijvenboden, K.; Postma, A.V.; Gloschat, C.R.; Elsey, R.M.; Sedmera, D.; et al. Specialized impulse conduction pathway in the alligator heart. eLife 2018, 7, e32120. [Google Scholar] [CrossRef]
- Kvasilova, A.; Olejnickova, V.; Jensen, B.; Christoffels, V.M.; Kolesova, H.; Sedmera, D.; Gregorovicova, M. The formation of the atrioventricular conduction axis is linked in development to ventricular septation. J. Exp. Biol. 2020, 223 Pt 19. [Google Scholar] [CrossRef]
- Miquerol, L.; Meysen, S.; Mangoni, M.; Bois, P.; van Rijen, H.V.; Abran, P.; Jongsma, H.; Nargeot, J.; Gros, D. Architectural and functional asymmetry of the His-Purkinje system of the murine heart. Cardiovasc. Res. 2004, 63, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankova, B.; Benes, J., Jr.; Krejci, E.; Dupays, L.; Theveniau-Ruissy, M.; Miquerol, L.; Sedmera, D. The effect of connexin40 deficiency on ventricular conduction system function during development. Cardiovasc. Res. 2012, 95, 469–479. [Google Scholar] [CrossRef] [Green Version]
- Martinez, M.E.; Walton, R.D.; Bayer, J.D.; Haissaguerre, M.; Vigmond, E.J.; Hocini, M.; Bernus, O. Role of the Purkinje-Muscle Junction on the Ventricular Repolarization Heterogeneity in the Healthy and Ischemic Ovine Ventricular Myocardium. Front. Physiol. 2018, 9, 718. [Google Scholar] [CrossRef] [Green Version]
- Boyden, P.A.; Hirose, M.; Dun, W. Cardiac Purkinje cells. Heart Rhythm. 2010, 7, 127–135. [Google Scholar] [CrossRef]
- Rentschler, S.; Vaidya, D.M.; Tamaddon, H.; Degenhardt, K.; Sassoon, D.; Morley, G.E.; Jalife, J.; Fishman, G.I. Visualization and functional characterization of the developing murine cardiac conduction system. Development 2001, 128, 1785–1792. [Google Scholar]
- Morley, G.E.; Vaidya, D. Understanding conduction of electrical impulses in the mouse heart using high-resolution video imaging technology. Microsc. Res. Tech. 2001, 52, 241–250. [Google Scholar] [CrossRef]
- Sedmera, D.; Gourdie, R.G. Why do we have Purkinje fibers deep in our heart? Physiol. Res. 2014, 63 (Suppl. 1), S9–S18. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthi, S.; Perotti, L.E.; Borgstrom, N.P.; Ajijola, O.A.; Frid, A.; Ponnaluri, A.V.; Weiss, J.N.; Qu, Z.; Klug, W.S.; Ennis, D.B.; et al. Simulation Methods and Validation Criteria for Modeling Cardiac Ventricular Electrophysiology. PLoS ONE 2014, 9, e114494. [Google Scholar] [CrossRef]
- Wiegerinck, R.F.; van Veen, T.A.; Belterman, C.N.; Schumacher, C.A.; Noorman, M.; de Bakker, J.M.; Coronel, R. Transmural dispersion of refractoriness and conduction velocity is associated with heterogeneously reduced connexin43 in a rabbit model of heart failure. Heart Rhythm. 2008, 5, 1178–1185. [Google Scholar] [CrossRef]
- Akar, F.G.; Spragg, D.D.; Tunin, R.S.; Kass, D.A.; Tomaselli, G.F. Mechanisms underlying conduction slowing and arrhythmogenesis in nonischemic dilated cardiomyopathy. Circ. Res. 2004, 95, 717–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glukhov, A.V.; Fedorov, V.V.; Lou, Q.; Ravikumar, V.K.; Kalish, P.W.; Schuessler, R.B.; Moazami, N.; Efimov, I.R. Transmural dispersion of repolarization in failing and nonfailing human ventricle. Circ. Res. 2010, 106, 981–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tawara, S. Reizleitungssystem des Säugetierherzens—Eine anatomisch—Pathlogische Studie Über das Atrioventrikularbündel und die Purkinjeschen Fäden; Verlag von Gustav Fischer: Jena, Germany, 1906; p. 252. [Google Scholar]
- Ryu, S.; Yamamoto, S.; Andersen, C.R.; Nakazawa, K.; Miyake, F.; James, T.N. Intramural Purkinje cell network of sheep ventricles as the terminal pathway of conduction system. Anat. Rec. 2009, 292, 12–22. [Google Scholar] [CrossRef]
- Gourdie, R.G.; Mima, T.; Thompson, R.P.; Mikawa, T. Terminal diversification of the myocyte lineage generates Purkinje fibers of the cardiac conduction system. Development 1995, 121, 1423–1431. [Google Scholar] [PubMed]
- Takebayashi-Suzuki, K.; Yanagisawa, M.; Gourdie, R.G.; Kanzawa, N.; Mikawa, T. In vivo induction of cardiac Purkinje fiber differentiation by coexpression of preproendothelin-1 and endothelin converting enzyme-1. Development 2000, 127, 3523–3532. [Google Scholar]
- Durrer, D.; van Dam, R.T.; Freud, G.E.; Janse, M.J.; Meijler, F.L.; Arzbaecher, R.C. Total excitation of the isolated human heart. Circulation 1970, 41, 899–912. [Google Scholar] [CrossRef] [Green Version]
- Choquet, C.; Kelly, R.G.; Miquerol, L. Nkx2-5 defines distinct scaffold and recruitment phases during formation of the murine cardiac Purkinje fiber network. Nat. Commun. 2020, 11, 5300. [Google Scholar] [CrossRef]
- Angst, B.D.; Khan, L.U.; Severs, N.J.; Whitely, K.; Rothery, S.; Thompson, R.P.; Magee, A.I.; Gourdie, R.G. Dissociated spatial patterning of gap junctions and cell adhesion junctions during postnatal differentiation of ventricular myocardium. Circ. Res. 1997, 80, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Litchenberg, W.H.; Norman, L.W.; Holwell, A.K.; Martin, K.L.; Hewett, K.W.; Gourdie, R.G. The rate and anisotropy of impulse propagation in the postnatal terminal crest are correlated with remodeling of Cx43 gap junction pattern. Cardiovasc. Res. 2000, 45, 379–387. [Google Scholar] [CrossRef]
- Kolesova, H.; Capek, M.; Radochova, B.; Janacek, J.; Sedmera, D. Comparison of different tissue clearing methods and 3D imaging techniques for visualization of GFP-expressing mouse embryos and embryonic hearts. Histochem. Cell Biol. 2016, 146, 141–152. [Google Scholar] [CrossRef]
- Miquerol, L.; Moreno-Rascon, N.; Beyer, S.; Dupays, L.; Meilhac, S.M.; Buckingham, M.E.; Franco, D.; Kelly, R.G. Biphasic development of the Mammalian ventricular conduction system. Circ. Res. 2010, 107, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, D.; Tamaddon, H.S.; Lo, C.W.; Taffet, S.M.; Delmar, M.; Morley, G.E.; Jalife, J. Null mutation of connexin43 causes slow propagation of ventricular activation in the late stages of mouse embryonic development. Circ. Res. 2001, 88, 1196–1202. [Google Scholar] [CrossRef] [Green Version]
- Olejnickova, V.; Sankova, B.; Sedmera, D.; Janacek, J. Trabecular Architecture Determines Impulse Propagation through the Early Embryonic Mouse Heart. Front. Physiol. 2018, 9, 1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scher, A.M.; Young, A.C.; Malmgren, A.L.; Erickson, R.V. Activation of the interventricular septum. Circ. Res. 1955, 3, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckhardt, A.; Kulhava, L.; Miksik, I.; Pataridis, S.; Hlavackova, M.; Vasinova, J.; Kolar, F.; Sedmera, D.; Ostadal, B. Proteomic analysis of cardiac ventricles: Baso-apical differences. Mol. Cell Biochem. 2018, 445, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Wessels, A.; Sedmera, D. Developmental anatomy of the heart: A tale of mice and man. Physiol. Genom. 2003, 15, 165–176. [Google Scholar] [CrossRef]
- Sedmera, D.; Grobety, M.; Reymond, C.; Baehler, P.; Kucera, P.; Kappenberger, L. Pacing-induced ventricular remodeling in the chick embryonic heart. Pediatr. Res. 1999, 45, 845–852. [Google Scholar] [CrossRef] [Green Version]
- Sedmera, D.; Pexieder, T.; Vuillemin, M.; Thompson, R.P.; Anderson, R.H. Developmental patterning of the myocardium. Anat. Rec. 2000, 258, 319–337. [Google Scholar] [CrossRef]
- Kajstura, J.; Urbanek, K.; Perl, S.; Hosoda, T.; Zheng, H.; Ogorek, B.; Ferreira-Martins, J.; Goichberg, P.; Rondon-Clavo, C.; Sanada, F.; et al. Cardiomyogenesis in the adult human heart. Circ. Res. 2010, 107, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wang, X.; Bunger, P.C.; Gerdes, A.M. Formation of binucleated cardiac myocytes in rat heart: I. Role of actin-myosin contractile ring. J. Mol. Cell. Cardiol. 1997, 29, 1541–1551. [Google Scholar] [CrossRef]
- Li, F.; Wang, X.; Capasso, J.M.; Gerdes, A.M. Rapid transition of cardiac myocytes from hyperplasia to hypertrophy during postnatal development. J. Mol. Cell. Cardiol. 1996, 28, 1737–1746. [Google Scholar] [CrossRef]
- Greiner, J.; Sankarankutty, A.C.; Seemann, G.; Seidel, T.; Sachse, F.B. Confocal Microscopy-Based Estimation of Parameters for Computational Modeling of Electrical Conduction in the Normal and Infarcted Heart. Front. Physiol. 2018, 9, 239. [Google Scholar] [CrossRef]
- Pallante, B.A.; Giovannone, S.; Fang-Yu, L.; Zhang, J.; Liu, N.; Kang, G.; Dun, W.; Boyden, P.A.; Fishman, G.I. Contactin-2 expression in the cardiac Purkinje fiber network. Circ. Arrhythm. Electrophysiol. 2010, 3, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Veeraraghavan, R.; Hoeker, G.S.; Alvarez-Laviada, A.; Hoagland, D.; Wan, X.; King, D.R.; Sanchez-Alonso, J.; Chen, C.; Jourdan, J.; Isom, L.L.; et al. The adhesion function of the sodium channel beta subunit (beta1) contributes to cardiac action potential propagation. eLife 2018, 7, e37610. [Google Scholar] [CrossRef]
- Gourdie, R.G. The Cardiac Gap Junction has Discrete Functions in Electrotonic and Ephaptic Coupling. Anat. Rec. 2019, 302, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedmera, D.; Thompson, R.P.; Kolar, F. Effect of increased pressure loading on heart growth in neonatal rats. J. Mol. Cell. Cardiol. 2003, 35, 301–309. [Google Scholar] [CrossRef]
- De la Rosa, A.J.; Dominguez, J.N.; Sedmera, D.; Sankova, B.; Hove-Madsen, L.; Franco, D.; Aranega, A. Functional suppression of Kcnq1 leads to early sodium channel remodeling and cardiac conduction system dysmorphogenesis. Cardiovasc. Res. 2013, 98, 504–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olejnickova, V.; Kocka, M.; Kvasilova, A.; Kolesova, H.; Dziacky, A.; Gidor, T.; Gidor, L.; Sankova, B.; Gregorovicova, M.; Gourdie, R.G.; et al. Gap Junctional Communication via Connexin43 between Purkinje Fibers and Working Myocytes Explains the Epicardial Activation Pattern in the Postnatal Mouse Left Ventricle. Int. J. Mol. Sci. 2021, 22, 2475. https://doi.org/10.3390/ijms22052475
Olejnickova V, Kocka M, Kvasilova A, Kolesova H, Dziacky A, Gidor T, Gidor L, Sankova B, Gregorovicova M, Gourdie RG, et al. Gap Junctional Communication via Connexin43 between Purkinje Fibers and Working Myocytes Explains the Epicardial Activation Pattern in the Postnatal Mouse Left Ventricle. International Journal of Molecular Sciences. 2021; 22(5):2475. https://doi.org/10.3390/ijms22052475
Chicago/Turabian StyleOlejnickova, Veronika, Matej Kocka, Alena Kvasilova, Hana Kolesova, Adam Dziacky, Tom Gidor, Lihi Gidor, Barbora Sankova, Martina Gregorovicova, Robert G. Gourdie, and et al. 2021. "Gap Junctional Communication via Connexin43 between Purkinje Fibers and Working Myocytes Explains the Epicardial Activation Pattern in the Postnatal Mouse Left Ventricle" International Journal of Molecular Sciences 22, no. 5: 2475. https://doi.org/10.3390/ijms22052475
APA StyleOlejnickova, V., Kocka, M., Kvasilova, A., Kolesova, H., Dziacky, A., Gidor, T., Gidor, L., Sankova, B., Gregorovicova, M., Gourdie, R. G., & Sedmera, D. (2021). Gap Junctional Communication via Connexin43 between Purkinje Fibers and Working Myocytes Explains the Epicardial Activation Pattern in the Postnatal Mouse Left Ventricle. International Journal of Molecular Sciences, 22(5), 2475. https://doi.org/10.3390/ijms22052475