
Ca2+ Signalling Induced by NGF Identifies a Subset of Capsaicin-Excitable Neurons Displaying Enhanced Chemo-Nociception in Dorsal Root Ganglion Explants from Adult pirt-GCaMP3 Mouse

Abstract
:1. Introduction
2. Results
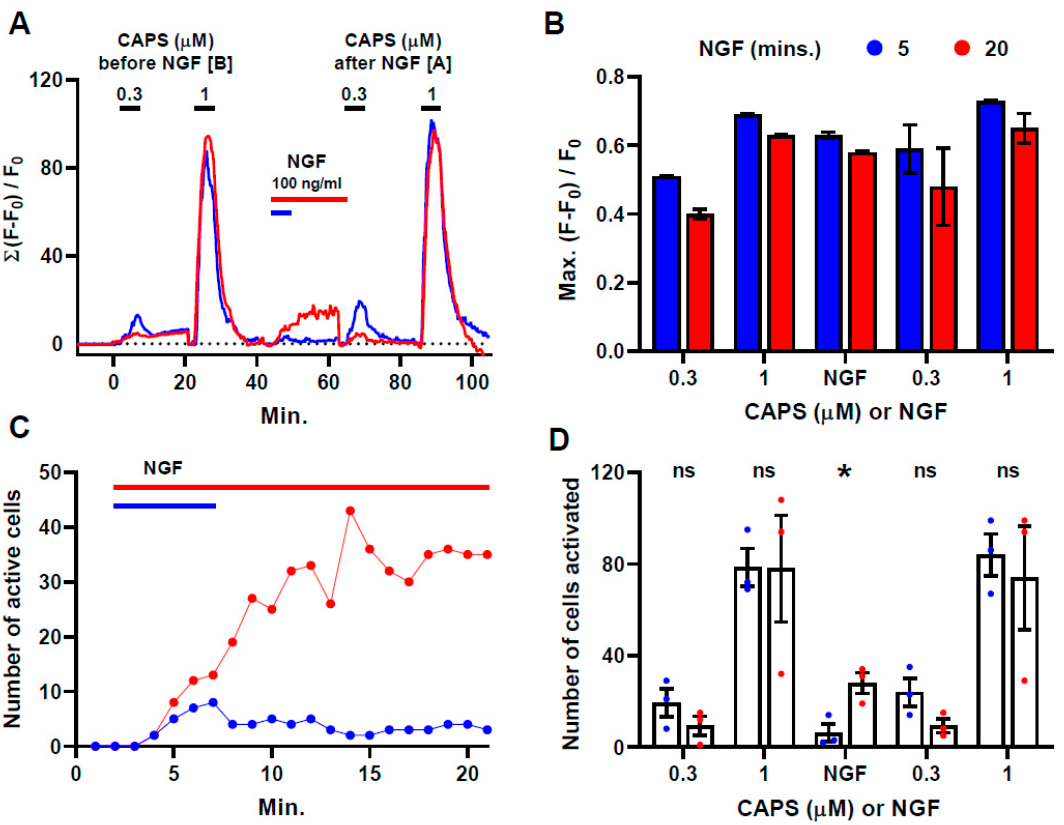
2.1. NGF Induces a Slow but Persistent Increase in [Ca2+]i Signals in a Sub-Population of CAPS-Excitable Neurons
2.2. NGF Attenuates CAPS-Induced Tachyphylaxis Which Occurs in a Sub-Population of the DRGNs
2.3. NGF-Excitable Cells Are More Responsive to CAPS
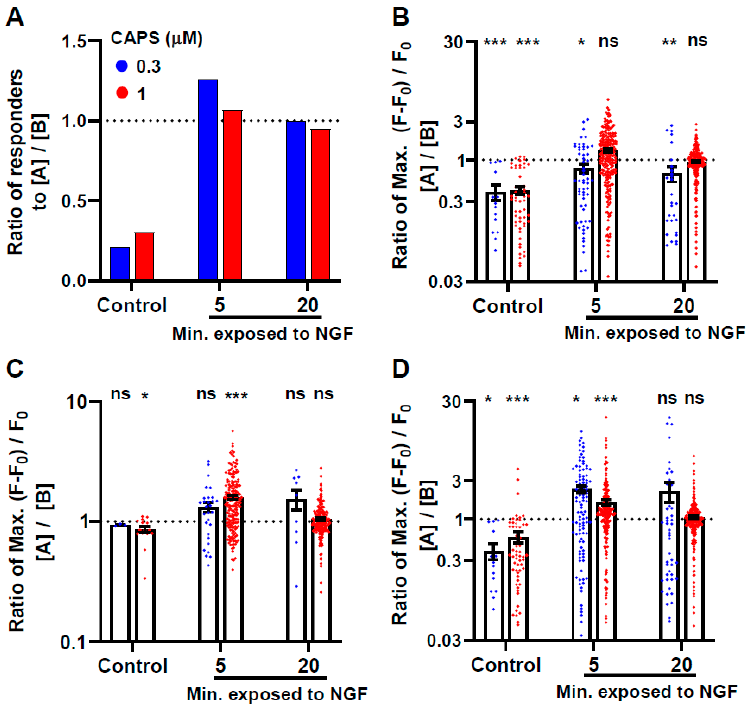
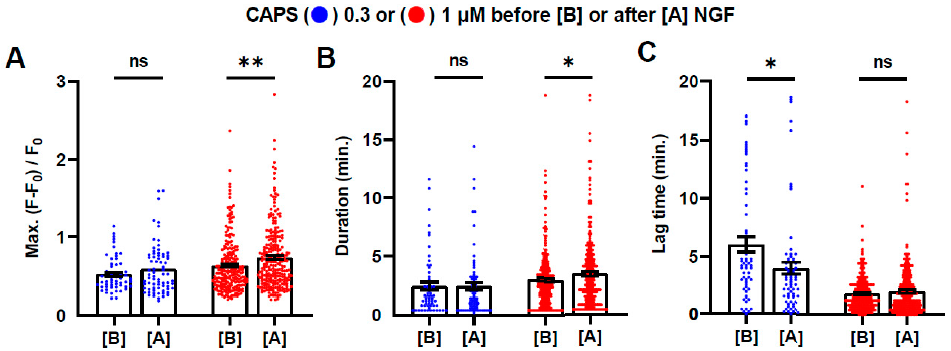
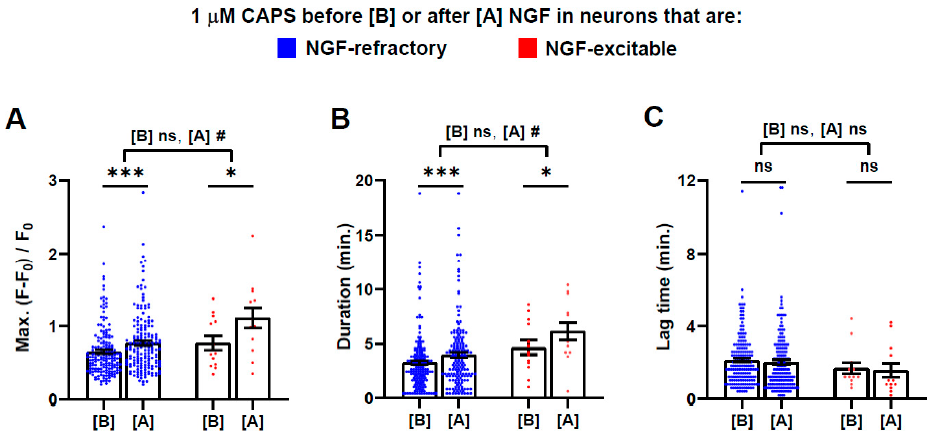
2.4. Brief Exposure to NGF Further Enhances CAPS-Induced Signals
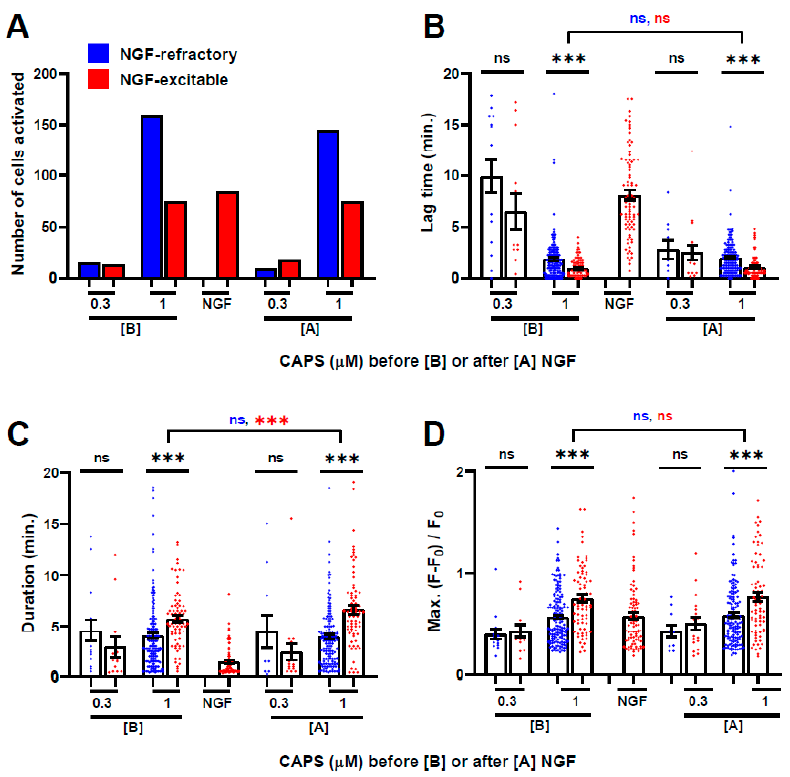
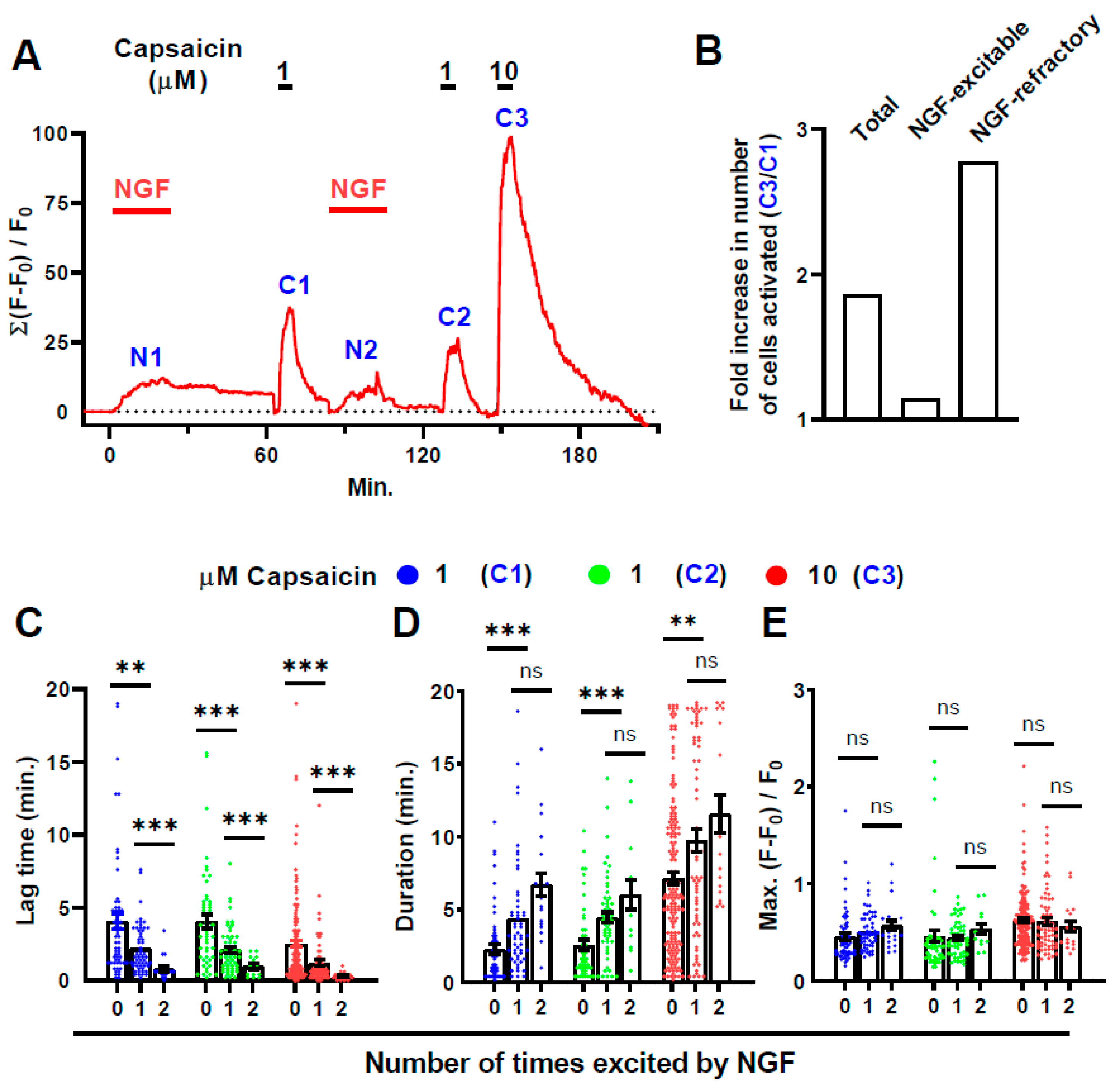
2.5. Prior Exposure to CAPS Is Not Necessary for the Activation by NGF of DRGNs: A Second Exposure to NGF Excites Even More Cells
2.6. Almost All the Cells Excited Only by 10, But Not 1, µΜ CAPS Were Recruited from a DRGN Population Refractory to NGF
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Confocal Imaging of DRG Explants
4.3. Image Analysis and Measurements of Signal Properties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| aCSF | Artificial cerebrospinal fluid |
| aCSF-BOC | Carboxygenated aCSF containing bovine serum albumin |
| [A] | After NGF |
| [B] | Before NGF |
| [Ca2+]i | Intracellular Ca2+ concentration |
| CAPS | Capsaicin |
| DRG | Dorsal root ganglia |
| DRGNs | DRG neurons |
| NGF | Nerve growth factor |
| ROI | Region of interest |
| s.d. | Standard deviation |
| s.e.m. | Standard error of the mean |
| SNAP-25 | Synaptosomal-associated protein of 25 kDa |
| TG | Trigeminal ganglia |
| TNFα | Tumour necrosis factor alpha |
| TrkA | Tropomyosin kinase receptor A |
| TRPA1 | TRP ankyrin family member 1 |
| TRPV1 | Transient receptor potential vanilloid 1 |
References
- Julius, D. TRP Channels and Pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef] [Green Version]
- Nilius, B.; Owsianik, G. The transient receptor potential family of ion channels. Genome Biol. 2011, 12, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nat. Cell Biol. 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Bélanger, E.; Côté, S.L.; Desrosiers, P.; Prescott, S.A.; Côté, D.C.; de Koninck, Y. Sensory Afferents Use Different Coding Strategies for Heat and Cold. Cell Rep. 2018, 23, 2001–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Chu, Y.; Han, L.; Li, M.; Li, Z.; LaVinka, P.C.; Sun, S.; Tang, Z.; Park, K.; Caterina, M.J.; et al. Central Terminal Sensitization of TRPV1 by Descending Serotonergic Facilitation Modulates Chronic Pain. Neuron 2014, 81, 873–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnington, J.K.; McNaughton, P.A. Signalling pathways involved in the sensitisation of mouse nociceptive neurones by nerve growth factor. J. Physiol. 2003, 551, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Chuang, H.-H.; Prescott, E.D.; Kong, H.; Shields, S.D.; Jordt, S.-E.; Basbaum, A.I.; Chao, M.V.; Julius, D. Bradykinin and nerve growth factor release the capsaicin receptor from PtdIns(4,5)P2-mediated inhibition. Nat. Cell Biol. 2001, 411, 957–962. [Google Scholar] [CrossRef]
- Denk, F.; Bennett, D.L.; McMahon, S.B. Nerve Growth Factor and Pain Mechanisms. Annu. Rev. Neurosci. 2017, 40, 307–325. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, X.; McNaughton, P.A. Inflammatory Pain: The Cellular Basis of Heat Hyperalgesia. Curr. Neuropharmacol. 2006, 4, 197–206. [Google Scholar] [CrossRef]
- Shu, X.; Mendell, L.M. Nerve growth factor acutely sensitizes the response of adult rat sensory neurons to capsaicin. Neurosci. Lett. 1999, 274, 159–162. [Google Scholar] [CrossRef]
- Shu, X.; Mendell, L.M. Acute Sensitization by NGF of the Response of Small-Diameter Sensory Neurons to Capsaicin. J. Neurophysiol. 2001, 86, 2931–2938. [Google Scholar] [CrossRef]
- Liu, J.; Yu, J.; Cheung, C. Immune Actions on the Peripheral Nervous System in Pain. Int. J. Mol. Sci. 2021, 22, 1448. [Google Scholar] [CrossRef]
- Andreev, N.Y.; Dimitrieva, N.; Koltzenburg, M.; McMahon, S.B. Peripheral administration of nerve growth factor in the adult rat produces a thermal hyperalgesia that requires the presence of sympathetic post-ganglionic neurones. Pain 1995, 63, 109–115. [Google Scholar] [CrossRef]
- Lewin, G.R.; Ritter, A.M.; Mendell, L.M. Nerve growth factor-induced hyperalgesia in the neonatal and adult rat. J. Neurosci. 1993, 13, 2136–2148. [Google Scholar] [CrossRef] [PubMed]
- Petty, B.G.; Cornblath, D.R.; Adornato, B.T.; Chaudhry, V.; Flexner, C.; Wachsman, M.; Sinicropi, D.; Burton, L.E.; Peroutka, S.J. The effect of systemically administered recombinant human nerve growth factor in healthy human subjects. Ann. Neurol. 1994, 36, 244–246. [Google Scholar] [CrossRef] [PubMed]
- Bannwarth, B.; Kostine, M. Nerve Growth Factor Antagonists: Is the Future of Monoclonal Antibodies Becoming Clearer? Drugs 2017, 77, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Barker, P.A.; Mantyh, P.; Arendt-Nielsen, L.; Viktrup, L.; Tive, L. Nerve Growth Factor Signaling and Its Contribution to Pain. J. Pain Res. 2020, 13, 1223–1241. [Google Scholar] [CrossRef]
- Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with aδ/c-fibers and colocalization with trk receptors. J. Comp. Neurol. 2005, 493, 596–606. [Google Scholar] [CrossRef]
- Bergmann, I.; Reiter, R.; Toyka, K.V.; Koltzenburg, M. Nerve growth factor evokes hyperalgesia in mice lacking the low-affinity neurotrophin receptor p75. Neurosci. Lett. 1998, 255, 87–90. [Google Scholar] [CrossRef]
- McMahon, S.B.; Armanini, M.P.; Ling, L.H.; Phillips, H.S. Expression and coexpression of Trk receptors in subpopulations of adult primary sensory neurons projecting to identified peripheral targets. Neuron 1994, 12, 1161–1171. [Google Scholar] [CrossRef]
- Omerbasic, D.; Smith, E.S.J.; Moroni, M.; Homfeld, J.; Eigenbrod, O.; Bennett, N.C.; Reznick, J.; Faulkes, C.G.; Selbach, M.; Lewin, G.R. Hypofunctional TrkA Accounts for the Absence of Pain Sensitization in the African Naked Mole-Rat. Cell Rep. 2016, 17, 748–758. [Google Scholar] [CrossRef] [Green Version]
- Michael, G.J.; Priestley, J.V. Differential Expression of the mRNA for the Vanilloid Receptor Subtype 1 in Cells of the Adult Rat Dorsal Root and Nodose Ganglia and Its Downregulation by Axotomy. J. Neurosci. 1999, 19, 1844–1854. [Google Scholar] [CrossRef] [Green Version]
- Stein, A.T.; Ufret-Vincenty, C.A.; Hua, L.; Santana, L.F.; Gordon, S.E. Phosphoinositide 3-Kinase Binds to TRPV1 and Mediates NGF-stimulated TRPV1 Trafficking to the Plasma Membrane. J. Gen. Physiol. 2006, 128, 509–522. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Huang, J.; McNaughton, P.A. NGF rapidly increases membrane expression of TRPV1 heat-gated ion channels. EMBO J. 2005, 24, 4211–4223. [Google Scholar] [CrossRef] [Green Version]
- Camprubí-Robles, M.; Planells-Cases, R.; Ferrer-Montiel, A. Differential contribution of SNARE-dependent exocytosis to inflammatory potentiation of TRPV1 in nociceptors. FASEB J. 2009, 23, 3722–3733. [Google Scholar] [CrossRef] [PubMed]
- Nugent, M.; Yusef, Y.R.; Meng, J.; Wang, J.; Dolly, J.O. A SNAP-25 cleaving chimera of botulinum neurotoxin /A and /E prevents TNFα−induced elevation of the activities of native TRP channels on early postnatal rat dorsal root ganglion neurons. Neuropharmacology 2018, 138, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Devesa, I.; Ferrandiz-Huertas, C.; Mathivanan, S.; Wolf, C.; Lujan, R.; Changeux, J.P.; Ferrer-Montiel, A. alphaCGRP is essential for algesic exocytotic mobilization of TRPV1 channels in peptidergic nociceptors. Proc. Natl. Acad. Sci. USA 2014, 111, 18345–18350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Wang, J.; Steinhoff, M.; Dolly, J.O. TNFalpha induces co-trafficking of TRPV1/TRPA1 in VAMP1-containing vesicles to the plasmalemma via Munc18-1/syntaxin1/SNAP-25 mediated fusion. Sci. Rep. 2016, 6, 21226. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Hu, J.; Xie, C.; Mei, K.; Pham, C.; Mo, X.; Hepp, R.; Soares, S.; Nothias, F.; Wang, Y.; et al. Recovery from tachyphylaxis of TRPV1 coincides with recycling to the surface membrane. Proc. Natl. Acad. Sci. USA 2019, 116, 5170–5175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Anderson, M.; Park, K.; Zheng, Q.; Agarwal, A.; Gong, C.; Young, L.; He, S.; La Vinka, P.C.; Zhou, F.; et al. Coupled Activation of Primary Sensory Neurons Contributes to Chronic Pain. Neuron 2016, 91, 1085–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Oxford, G.S. Phosphoinositide-3-kinase and mitogen activated protein kinase signaling pathways mediate acute NGF sensitization of TRPV1. Mol. Cell. Neurosci. 2007, 34, 689–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, S.B.; Bennett, D.L.H.; Priestley, J.V.; Shelton, D.L. The biological effects of endogenous nerve growth factor on adult sensory neurons revealed by a trkA-IgG fusion molecule. Nat. Med. 1995, 1, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Bannwarth, B.; Kostine, M. Targeting Nerve Growth Factor (NGF) for Pain Management: What Does the Future Hold for NGF Antagonists? Drugs 2014, 74, 619–626. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Neurons Excited by the Reference Stimulus and Exposure to 3:- | |||||||
|---|---|---|---|---|---|---|---|
| Cells Activated by Reference Stimulus 1,2 | 1 μM CAPS [B] | 100 ng/mL NGF | 0.3 μM CAPS [A] | 1 μM CAPS [A] | 1 μM CAPS [A] or [B] 5 | Only Excited by the Reference Stimulus | |
| 0.3 µM CAPS [B] 4 | 28 | 23 (82) | 13 (46) | 9 (32) | 20 (71) | 26 (93) | 1 (4) |
| 1 µM CAPS [B] 4 | 234 | - | 75 (32) | 20 (9) | 195 (83) | 234 (100) | 28 (12) |
| 100 ng/mL NGF (20 min) | 84 | - | - | 18 (21) | 76 (90) | 81 (96) | 2 (2) 6 |
| 0.3 µM CAPS [A] 4 | 28 | - | - | - | 24 (86) | 26 (93) | 2 (7) |
| 1 µM CAPS [A] 4 | 219 | - | - | - | - | 219 (100) | 14 (6) |
| Number of Neurons Excited by the Reference Stimulus and Exposure to 3: - | |||||||
|---|---|---|---|---|---|---|---|
| Cells Activated by Reference Stimulus 1,2 | 1 μM CAPS [C1] | 100 ng/mL NGF [N2] | 1 μM CAPS [C2] | 1 μM CAPS [C1 or C2] 4 | 10 μM CAPS [C3] | Only Excited by the Reference Stimulus | |
| 100 ng/mL NGF [N1] 5 | 57 | 47 (82) | 21 (37) | 35 (61) | 49 (86) | 48 (84) | 3 (5) |
| 1 μM CAPS [C1] 5 | 135 | - | 50 (37) | 80 (59) | 135 (100) | 116 (86) | 6 (4) |
| 100 ng/mL NGF [N2] 5 | 72 | - | - | 48 (67) | 60 (83) | 58 (81) | 5 (7) |
| 1 μM CAPS [C2] 5 | 121 | - | - | - | 121 (100) | 121 (100) | 20 (17) |
| 10 μM CAPS [C3] 5 | 251 | - | - | - | 132 (53) | - | 107 (43) |
| 100 ng/mL NGF [N1 or N2] 5,6 | 108 | 76 (70) | 72 (67) | 69 (64) | 88 (81) | 87 (81) | 5 (5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lawrence, G.W.; Zurawski, T.H.; Dolly, J.O. Ca2+ Signalling Induced by NGF Identifies a Subset of Capsaicin-Excitable Neurons Displaying Enhanced Chemo-Nociception in Dorsal Root Ganglion Explants from Adult pirt-GCaMP3 Mouse. Int. J. Mol. Sci. 2021, 22, 2589. https://doi.org/10.3390/ijms22052589
Lawrence GW, Zurawski TH, Dolly JO. Ca2+ Signalling Induced by NGF Identifies a Subset of Capsaicin-Excitable Neurons Displaying Enhanced Chemo-Nociception in Dorsal Root Ganglion Explants from Adult pirt-GCaMP3 Mouse. International Journal of Molecular Sciences. 2021; 22(5):2589. https://doi.org/10.3390/ijms22052589
Chicago/Turabian StyleLawrence, Gary W., Tomas H. Zurawski, and J. Oliver Dolly. 2021. "Ca2+ Signalling Induced by NGF Identifies a Subset of Capsaicin-Excitable Neurons Displaying Enhanced Chemo-Nociception in Dorsal Root Ganglion Explants from Adult pirt-GCaMP3 Mouse" International Journal of Molecular Sciences 22, no. 5: 2589. https://doi.org/10.3390/ijms22052589
APA StyleLawrence, G. W., Zurawski, T. H., & Dolly, J. O. (2021). Ca2+ Signalling Induced by NGF Identifies a Subset of Capsaicin-Excitable Neurons Displaying Enhanced Chemo-Nociception in Dorsal Root Ganglion Explants from Adult pirt-GCaMP3 Mouse. International Journal of Molecular Sciences, 22(5), 2589. https://doi.org/10.3390/ijms22052589