
Sitosterolemia: Twenty Years of Discovery of the Function of ABCG5ABCG8


Abstract
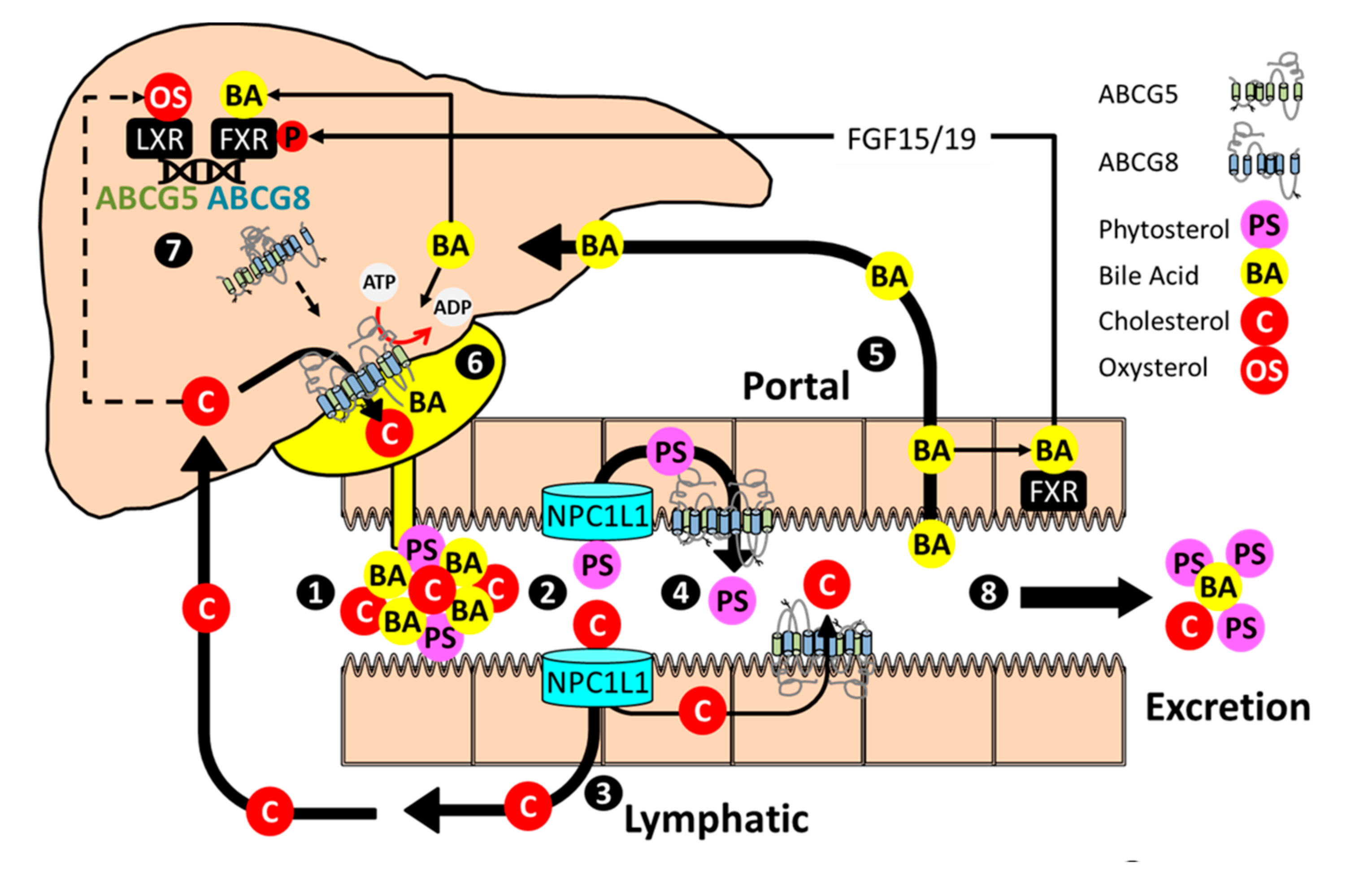
:1. Discovery to Therapy
2. Animal Models of Sitosterolemia
3. Heterologous Expression of G5G8
4. Beyond Phytosterols
5. Transcriptional Regulation of G5G8
6. Post-Transcriptional Regulation
7. Sitosterolemia and/or Familial Hypercholesterolemia
8. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bhattacharyya, A.K.; Connor, W.E. Beta-sitosterolemia and xanthomatosis. A newly described lipid storage disease in two sisters. J. Clin. Investig. 1974, 53, 1033–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miettinen, T.A. Phytosterolaemia, xanthomatosis and premature atherosclerotic arterial disease: A case with high plant sterol absorption, impaired sterol elimination and low cholesterol synthesis. Eur. J. Clin. Investig. 1980, 10, 27–35. [Google Scholar] [CrossRef]
- Kwiterovich, P.O., Jr.; Bachorik, P.S.; Smith, H.H.; McKusick, V.A.; Connor, W.E.; Teng, B.; Sniderman, A.D. Hyperapobetalipoproteinaemia in two families with xanthomas and phytosterolaemia. Lancet 1981, 1, 466–469. [Google Scholar] [CrossRef]
- Shulman, R.S.; Bhattacharyya, A.K.; Connor, W.E.; Fredrickson, D.S. Beta-sitosterolemia and xanthomatosis. N. Engl. J. Med. 1976, 294, 482–483. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.J.; Wang, C.; Salen, G.; Lam, K.C.; Chan, T.K. Sitosterol and cholesterol metabolism in a patient with coexisting phytosterolemia and cholestanolemia. Metab. Clin. Exp. 1983, 32, 126–133. [Google Scholar] [CrossRef]
- Beaty, T.H.; Kwiterovich, P.O., Jr.; Khoury, M.J.; White, S.; Bachorik, P.S.; Smith, H.H.; Teng, B.; Sniderman, A. Genetic analysis of plasma sitosterol, apoprotein B, and lipoproteins in a large Amish pedigree with sitosterolemia. Am. J. Hum. Genet. 1986, 38, 492–504. [Google Scholar]
- Salen, G.; Horak, I.; Rothkopf, M.; Cohen, J.L.; Speck, J.; Tint, G.S.; Shore, V.; Dayal, B.; Chen, T.; Shefer, S. Lethal atherosclerosis associated with abnormal plasma and tissue sterol composition in sitosterolemia with xanthomatosis. J. Lipid Res. 1985, 26, 1126–1133. [Google Scholar] [CrossRef]
- Nguyen, L.B.; Salen, G.; Shefer, S.; Tint, G.S.; Shore, V.; Ness, G.C. Decreased cholesterol biosynthesis in sitosterolemia with xanthomatosis: Diminished mononuclear leukocyte 3-hydroxy-3-methylglutaryl coenzyme A reductase activity and enzyme protein associated with increased low-density lipoprotein receptor function. Metab. Clin. Exp. 1990, 39, 436–443. [Google Scholar] [CrossRef]
- Bhattacharyya, A.K.; Connor, W.E.; Lin, D.S.; McMurry, M.M.; Shulman, R.S. Sluggish sitosterol turnover and hepatic failure to excrete sitosterol into bile cause expansion of body pool of sitosterol in patients with sitosterolemia and xanthomatosis. Arterioscler. Thromb. A J. Vasc. Biol./Am. Heart Assoc. 1991, 11, 1287–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, D.C.; Iolascon, A.; Carella, M.; O’Marcaigh, A.S.; Kendra, J.R.; Jowitt, S.N.; Wales, J.K.; Vora, A.; Makris, M.; Manning, N.; et al. Stomatocytic haemolysis and macrothrombocytopenia (Mediterranean stomatocytosis/macrothrombocytopenia) is the haematological presentation of phytosterolaemia. Br. J. Haematol. 2005, 130, 297–309. [Google Scholar] [CrossRef]
- Mushtaq, T.; Wales, J.K.; Wright, N.P. Adrenal insufficiency in phytosterolaemia. Eur. J. Endocrinol. 2007, 157 (Suppl. 1), S61–S65. [Google Scholar] [CrossRef]
- Wang, Z.; Cao, L.; Su, Y.; Wang, G.; Wang, R.; Yu, Z.; Bai, X.; Ruan, C. Specific macrothrombocytopenia/hemolytic anemia associated with sitosterolemia. Am. J. Hematol. 2014, 89, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Bazerbachi, F.; Conboy, E.E.; Mounajjed, T.; Watt, K.D.; Babovic-Vuksanovic, D.; Patel, S.B.; Kamath, P.S. Cryptogenic Cirrhosis and Sitosterolemia: A Treatable Disease If Identified but Fatal If Missed. Ann. Hepatol. 2017, 16, 970–978. [Google Scholar] [CrossRef]
- Salen, G.; Shore, V.; Tint, G.S.; Forte, T.; Shefer, S.; Horak, I.; Horak, E.; Dayal, B.; Nguyen, L.; Batta, A.K.; et al. Increased sitosterol absorption, decreased removal, and expanded body pools compensate for reduced cholesterol synthesis in sitosterolemia with xanthomatosis. J. Lipid Res. 1989, 30, 1319–1330. [Google Scholar] [CrossRef]
- Cobb, M.M.; Salen, G.; Tint, G.S. Comparative effect of dietary sitosterol on plasma sterols and cholesterol and bile acid synthesis in a sitosterolemic homozygote and heterozygote subject. J. Am. Coll. Nutr. 1997, 16, 605–613. [Google Scholar]
- Nguyen, L.B.; Cobb, M.; Shefer, S.; Salen, G.; Ness, G.C.; Tint, G.S. Regulation of cholesterol biosynthesis in sitosterolemia: Effects of lovastatin, cholestyramine, and dietary sterol restriction. J. Lipid Res. 1991, 32, 1941–1948. [Google Scholar] [CrossRef]
- Patel, S.B.; Honda, A.; Salen, G. Sitosterolemia: Exclusion of genes involved in reduced cholesterol biosynthesis. J. Lipid Res. 1998, 39, 1055–1061. [Google Scholar] [CrossRef]
- Patel, S.B.; Salen, G.; Hidaka, H.; Kwiterovich, P.O.; Stalenhoef, A.F.; Miettinen, T.A.; Grundy, S.M.; Lee, M.H.; Rubenstein, J.S.; Polymeropoulos, M.H.; et al. Mapping a gene involved in regulating dietary cholesterol absorption. The sitosterolemia locus is found at chromosome 2p21. J. Clin. Investig. 1998, 102, 1041–1044. [Google Scholar] [CrossRef] [PubMed]
- Peet, D.J.; Turley, S.D.; Ma, W.; Janowski, B.A.; Lobaccaro, J.M.; Hammer, R.E.; Mangelsdorf, D.J. Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXR alpha. Cell 1998, 93, 693–704. [Google Scholar] [CrossRef] [Green Version]
- Repa, J.J.; Turley, S.D.; Lobaccaro, J.A.; Medina, J.; Li, L.; Lustig, K.; Shan, B.; Heyman, R.A.; Dietschy, J.M.; Mangelsdorf, D.J. Regulation of absorption and ABC1-mediated efflux of cholesterol by RXR heterodimers. Science 2000, 289, 1524–1529. [Google Scholar] [CrossRef]
- Ewart, G.D.; Cannell, D.; Cox, G.B.; Howells, A.J. Mutational analysis of the traffic ATPase (ABC) transporters involved in uptake of eye pigment precursors in Drosophila melanogaster. Implications for structure-function relationships. J. Biol. Chem. 1994, 269, 10370–10377. [Google Scholar] [CrossRef]
- Berge, K.E.; Tian, H.; Graf, G.A.; Yu, L.; Grishin, N.V.; Schultz, J.; Kwiterovich, P.; Shan, B.; Barnes, R.; Hobbs, H.H. Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science 2000, 290, 1771–1775. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Lu, K.; Hazard, S.; Yu, H.; Shulenin, S.; Hidaka, H.; Kojima, H.; Allikmets, R.; Sakuma, N.; Pegoraro, R.; et al. Identification of a gene, ABCG5, important in the regulation of dietary cholesterol absorption. Nat. Genet. 2001, 27, 79–83. [Google Scholar] [CrossRef]
- Van Heek, M.; France, C.F.; Compton, D.S.; McLeod, R.L.; Yumibe, N.P.; Alton, K.B.; Sybertz, E.J.; Davis, H.R., Jr. In vivo metabolism-based discovery of a potent cholesterol absorption inhibitor, SCH58235, in the rat and rhesus monkey through the identification of the active metabolites of SCH48461. J. Pharm. Exp. Ther. 1997, 283, 157–163. [Google Scholar]
- Davis, H.R., Jr.; Zhu, L.J.; Hoos, L.M.; Tetzloff, G.; Maguire, M.; Liu, J.; Yao, X.; Iyer, S.P.; Lam, M.H.; Lund, E.G.; et al. Niemann-Pick C1 Like 1 (NPC1L1) is the intestinal phytosterol and cholesterol transporter and a key modulator of whole-body cholesterol homeostasis. J. Biol. Chem. 2004, 279, 33586–33592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salen, G.; von Bergmann, K.; Lutjohann, D.; Kwiterovich, P.; Kane, J.; Patel, S.B.; Musliner, T.; Stein, P.; Musser, B.; Multicenter Sitosterolemia Study Group. Ezetimibe effectively reduces plasma plant sterols in patients with sitosterolemia. Circulation 2004, 109, 966–971. [Google Scholar] [CrossRef] [Green Version]
- Lutjohann, D.; von Bergmann, K.; Sirah, W.; Macdonell, G.; Johnson-Levonas, A.O.; Shah, A.; Lin, J.; Sapre, A.; Musliner, T. Long-term efficacy and safety of ezetimibe 10 mg in patients with homozygous sitosterolemia: A 2-year, open-label extension study. Int J. Clin. Pract. 2008, 62, 1499–1510. [Google Scholar] [CrossRef] [PubMed]
- Escola-Gil, J.C.; Quesada, H.; Julve, J.; Martin-Campos, J.M.; Cedo, L.; Blanco-Vaca, F. Sitosterolemia: Diagnosis, investigation, and management. Curr. Atheroscler. Rep. 2014, 16, 424. [Google Scholar] [CrossRef]
- Plosch, T.; Bloks, V.W.; Terasawa, Y.; Berdy, S.; Siegler, K.; Van Der Sluijs, F.; Kema, I.P.; Groen, A.K.; Shan, B.; Kuipers, F.; et al. Sitosterolemia in ABC-transporter G5-deficient mice is aggravated on activation of the liver-X receptor. Gastroenterology 2004, 126, 290–300. [Google Scholar] [CrossRef]
- Klett, E.L.; Lu, K.; Kosters, A.; Vink, E.; Lee, M.H.; Altenburg, M.; Shefer, S.; Batta, A.K.; Yu, H.; Chen, J.; et al. A mouse model of sitosterolemia: Absence of Abcg8/sterolin-2 results in failure to secrete biliary cholesterol. BMC Med. 2004, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Hammer, R.E.; Li-Hawkins, J.; Von Bergmann, K.; Lutjohann, D.; Cohen, J.C.; Hobbs, H.H. Disruption of Abcg5 and Abcg8 in mice reveals their crucial role in biliary cholesterol secretion. Proc. Natl. Acad. Sci. USA 2002, 99, 16237–16242. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Batta, A.; Zheng, S.; Fitzgibbon, W.R.; Ullian, M.E.; Yu, H.; Tso, P.; Salen, G.; Patel, S.B. The missense mutation in Abcg5 gene in spontaneously hypertensive rats (SHR) segregates with phytosterolemia but not hypertension. BMC Genet. 2005, 6, 40. [Google Scholar]
- Chase, T.H.; Lyons, B.L.; Bronson, R.T.; Foreman, O.; Donahue, L.R.; Burzenski, L.M.; Gott, B.; Lane, P.; Harris, B.; Ceglarek, U.; et al. The mouse mutation “thrombocytopenia and cardiomyopathy” (trac) disrupts Abcg5: A spontaneous single gene model for human hereditary phytosterolemia/sitosterolemia. Blood 2010, 115, 1267–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruit, J.K.; Drayer, A.L.; Bloks, V.W.; Blom, N.; Olthof, S.G.; Sauer, P.J.; de Haan, G.; Kema, I.P.; Vellenga, E.; Kuipers, F. Plant sterols cause macrothrombocytopenia in a mouse model of sitosterolemia. J. Biol. Chem. 2008, 283, 6281–6287. [Google Scholar] [CrossRef] [Green Version]
- Kanaji, T.; Kanaji, S.; Montgomery, R.R.; Patel, S.B.; Newman, P.J. Platelet hyperreactivity explains the bleeding abnormality and macrothrombocytopenia in a murine model of sitosterolemia. Blood 2013, 122, 2732–2742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solca, C.; Tint, G.S.; Patel, S.B. Dietary xenosterols lead to infertility and loss of abdominal adipose tissue in sterolin-deficient mice. J. Lipid Res. 2013, 54, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; von Bergmann, K.; Lutjohann, D.; Hobbs, H.H.; Cohen, J.C. Ezetimibe normalizes metabolic defects in mice lacking ABCG5 and ABCG8. J. Lipid Res. 2005, 46, 1739–1744. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Ma, Y.; Jia, L.; Ioannou, Y.A.; Davies, J.P.; Yu, L. Genetic inactivation of NPC1L1 protects against sitosterolemia in mice lacking ABCG5/ABCG8. J. Lipid Res. 2009, 50, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDaniel, A.L.; Alger, H.M.; Sawyer, J.K.; Kelley, K.L.; Kock, N.D.; Brown, J.M.; Temel, R.E.; Rudel, L.L. Phytosterol feeding causes toxicity in ABCG5/G8 knockout mice. Am. J. Pathol. 2013, 182, 1131–1138. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; McDonald, J.G.; Patel, A.; Zhang, Y.; Umetani, M.; Xu, F.; Westover, E.J.; Covey, D.F.; Mangelsdorf, D.J.; Cohen, J.C.; et al. Sterol intermediates from cholesterol biosynthetic pathway as liver X receptor ligands. J. Biol. Chem. 2006, 281, 27816–27826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, B.A.; Taylor, O.A.; Prendergast, D.R.; Zimmerman, T.L.; Von Furstenberg, R.; Moore, D.D.; Karpen, S.J. Stigmasterol, a soy lipid-derived phytosterol, is an antagonist of the bile acid nuclear receptor FXR. Pediatr. Res. 2007, 62, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Sabeva, N.S.; McPhaul, C.M.; Li, X.; Cory, T.J.; Feola, D.J.; Graf, G.A. Phytosterols differentially influence ABC transporter expression, cholesterol efflux and inflammatory cytokine secretion in macrophage foam cells. J. Nutr. Biochem. 2011, 22, 777–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plat, J.; Nichols, J.A.; Mensink, R.P. Plant sterols and stanols: Effects on mixed micellar composition and LXR (target gene) activation. J. Lipid Res. 2005, 46, 2468–2476. [Google Scholar] [CrossRef] [Green Version]
- Salen, G.; Tint, G.S.; Shefer, S.; Shore, V.; Nguyen, L. Increased sitosterol absorption is offset by rapid elimination to prevent accumulation in heterozygotes with sitosterolemia. Arterioscler. Thromb. A J. Vasc. Biol. Am. Heart Assoc. 1992, 12, 563–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lütjohann, D.; Björkhem, I.; Ose, L. Phytosterolaemia in a Norwegian family: Diagnosis and characterization of the first Scandinavian case. Scand. J. Clin. Lab. Investig. 1996, 56, 229–240. [Google Scholar] [CrossRef]
- Zhang, L.S.; Xu, M.; Yang, Q.; Lou, D.; Howles, P.N.; Tso, P. ABCG5/G8 deficiency in mice reduces dietary triacylglycerol and cholesterol transport into the lymph. Lipids 2015, 50, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.M.; Sawyer, J.K.; Kelley, K.L.; Davis, M.A.; Kent, C.R.; Rudel, L.L. ACAT2 and ABCG5/G8 are both required for efficient cholesterol absorption in mice: Evidence from thoracic lymph duct cannulation. J. Lipid Res. 2012, 53, 1598–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.H.; Patel, S.B.; Carey, M.C.; Wang, D.Q. Quantifying anomalous intestinal sterol uptake, lymphatic transport, and biliary secretion in Abcg8(-/-) mice. Hepatology 2007, 45, 998–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Li-Hawkins, J.; Hammer, R.E.; Berge, K.E.; Horton, J.D.; Cohen, J.C.; Hobbs, H.H. Overexpression of ABCG5 and ABCG8 promotes biliary cholesterol secretion and reduces fractional absorption of dietary cholesterol. J. Clin. Investig. 2002, 110, 671–680. [Google Scholar] [CrossRef]
- Jakulj, L.; Vissers, M.N.; van Roomen, C.P.; van der Veen, J.N.; Vrins, C.L.J.; Kunne, C.; Stellaard, F.; Kastelein, J.J.P.; Groen, A.K. Ezetimibe stimulates faecal neutral sterol excretion depending on abcg8 function in mice. FEBS Lett. 2010, 584, 3625–3628. [Google Scholar] [CrossRef] [Green Version]
- de Boer, J.F.; Schonewille, M.; Boesjes, M.; Wolters, H.; Bloks, V.W.; Bos, T.; van Dijk, T.H.; Jurdzinski, A.; Boverhof, R.; Wolters, J.C.; et al. Intestinal Farnesoid X Receptor Controls Transintestinal Cholesterol Excretion in Mice. Gastroenterology 2017, 152, 1126–1138.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonde, Y.; Plosch, T.; Kuipers, F.; Angelin, B.; Rudling, M. Stimulation of murine biliary cholesterol secretion by thyroid hormone is dependent on a functional ABCG5/G8 complex. Hepatology 2012, 56, 1828–1837. [Google Scholar] [CrossRef] [Green Version]
- Kosters, A.; Frijters, R.J.; Kunne, C.; Vink, E.; Schneiders, M.S.; Schaap, F.G.; Nibbering, C.P.; Patel, S.B.; Groen, A.K. Diosgenin-induced biliary cholesterol secretion in mice requires Abcg8. Hepatology 2005, 41, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Gupta, S.; Xu, F.; Liverman, A.D.; Moschetta, A.; Mangelsdorf, D.J.; Repa, J.J.; Hobbs, H.H.; Cohen, J.C. Expression of ABCG5 and ABCG8 is required for regulation of biliary cholesterol secretion. J. Biol. Chem. 2005, 280, 8742–8747. [Google Scholar] [CrossRef] [Green Version]
- Yamanashi, Y.; Takada, T.; Yoshikado, T.; Shoda, J.; Suzuki, H. NPC2 regulates biliary cholesterol secretion via stimulation of ABCG5/G8-mediated cholesterol transport. Gastroenterology 2011, 140, 1664–1674. [Google Scholar] [CrossRef]
- Dikkers, A.; de Boer, J.F.; Groen, A.K.; Tietge, U.J. Hepatic ABCG5/G8 overexpression substantially increases biliary cholesterol secretion but does not impact in vivo macrophage-to-feces RCT. Atherosclerosis 2015, 243, 402–406. [Google Scholar] [CrossRef]
- Coy, D.J.; Wooton-Kee, C.R.; Yan, B.; Sabeva, N.; Su, K.; Graf, G.; Vore, M. ABCG5/ABCG8-independent biliary cholesterol excretion in lactating rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G228–G235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groen, A.; Kunne, C.; Jongsma, G.; van den Oever, K.; Mok, K.S.; Petruzzelli, M.; Vrins, C.L.; Bull, L.; Paulusma, C.C.; Oude Elferink, R.P. Abcg5/8 independent biliary cholesterol excretion in Atp8b1-deficient mice. Gastroenterology 2008, 134, 2091–2100. [Google Scholar] [CrossRef]
- Wiersma, H.; Gatti, A.; Nijstad, N.; Oude Elferink, R.P.; Kuipers, F.; Tietge, U.J. Scavenger receptor class B type I mediates biliary cholesterol secretion independent of ATP-binding cassette transporter g5/g8 in mice. Hepatology 2009, 50, 1263–1272. [Google Scholar] [CrossRef]
- Wang, H.H.; Li, X.; Patel, S.B.; Wang, D.Q. Evidence that the adenosine triphosphate-binding cassette G5/G8-independent pathway plays a determinant role in cholesterol gallstone formation in mice. Hepatology 2016, 64, 853–864. [Google Scholar] [CrossRef] [Green Version]
- Wilund, K.R.; Yu, L.; Xu, F.; Hobbs, H.H.; Cohen, J.C. High-level expression of ABCG5 and ABCG8 attenuates diet-induced hypercholesterolemia and atherosclerosis in Ldlr-/-mice. J. Lipid Res. 2004, 45, 1429–1436. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.E.; Basso, F.; Shamburek, R.D.; Amar, M.J.; Vaisman, B.; Szakacs, G.; Joyce, C.; Tansey, T.; Freeman, L.; Paigen, B.J.; et al. Hepatic ABCG5 and ABCG8 overexpression increases hepatobiliary sterol transport but does not alter aortic atherosclerosis in transgenic mice. J. Biol. Chem. 2004, 279, 22913–22925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basso, F.; Freeman, L.A.; Ko, C.; Joyce, C.; Amar, M.J.; Shamburek, R.D.; Tansey, T.; Thomas, F.; Wu, J.; Paigen, B.; et al. Hepatic ABCG5/G8 overexpression reduces apoB-lipoproteins and atherosclerosis when cholesterol absorption is inhibited. J. Lipid Res. 2007, 48, 114–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calpe-Berdiel, L.; Rotllan, N.; Fievet, C.; Roig, R.; Blanco-Vaca, F.; Escola-Gil, J.C. Liver X receptor-mediated activation of reverse cholesterol transport from macrophages to feces in vivo requires ABCG5/G8. J. Lipid Res. 2008, 49, 1904–1911. [Google Scholar] [CrossRef] [Green Version]
- Altemus, J.B.; Patel, S.B.; Sehayek, E. Liver-specific induction of Abcg5 and Abcg8 stimulates reverse cholesterol transport in response to ezetimibe treatment. Metab. Clin. Exp. 2014, 63, 1334–1341. [Google Scholar] [CrossRef]
- Su, K.; Sabeva, N.S.; Wang, Y.; Liu, X.; Lester, J.D.; Liu, J.; Liang, S.; Graf, G.A. Acceleration of biliary cholesterol secretion restores glycemic control and alleviates hypertriglyceridemia in obese db/db mice. Arter. Thromb. Vasc. Biol. 2014, 34, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Mitsche, M.A.; Lutjohann, D.; Cohen, J.C.; Xie, X.S.; Hobbs, H.H. Relative roles of ABCG5/ABCG8 in liver and intestine. J. Lipid Res. 2015, 56, 319–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berge, K.E.; von Bergmann, K.; Lutjohann, D.; Guerra, R.; Grundy, S.M.; Hobbs, H.H.; Cohen, J.C. Heritability of plasma noncholesterol sterols and relationship to DNA sequence polymorphism in ABCG5 and ABCG8. J. Lipid Res. 2002, 43, 486–494. [Google Scholar] [CrossRef]
- Buch, S.; Schafmayer, C.; Volzke, H.; Becker, C.; Franke, A.; von Eller-Eberstein, H.; Kluck, C.; Bassmann, I.; Brosch, M.; Lammert, F.; et al. A genome-wide association scan identifies the hepatic cholesterol transporter ABCG8 as a susceptibility factor for human gallstone disease. Nat. Genet. 2007, 39, 995–999. [Google Scholar] [CrossRef]
- Grunhage, F.; Acalovschi, M.; Tirziu, S.; Walier, M.; Wienker, T.F.; Ciocan, A.; Mosteanu, O.; Sauerbruch, T.; Lammert, F. Increased gallstone risk in humans conferred by common variant of hepatic ATP-binding cassette transporter for cholesterol. Hepatology 2007, 46, 793–801. [Google Scholar] [CrossRef]
- Stender, S.; Frikke-Schmidt, R.; Nordestgaard, B.G.; Tybjaerg-Hansen, A. The ABCG5/8 cholesterol transporter and myocardial infarction versus gallstone disease. J. Am. Coll. Cardiol. 2014, 63, 2121–2128. [Google Scholar] [CrossRef] [Green Version]
- Helgadottir, A.; Thorleifsson, G.; Alexandersson, K.F.; Tragante, V.; Thorsteinsdottir, M.; Eiriksson, F.F.; Gretarsdottir, S.; Bjornsson, E.; Magnusson, O.; Sveinbjornsson, G.; et al. Genetic variability in the absorption of dietary sterols affects the risk of coronary artery disease. Eur. Heart J. 2020, 41, 2618–2628. [Google Scholar] [CrossRef]
- Jiang, Z.Y.; Cai, Q.; Chen, E.Z. Association of three common single nucleotide polymorphisms of ATP binding cassette G8 gene with gallstone disease: A meta-analysis. PLoS ONE 2014, 9, e87200. [Google Scholar]
- Viturro, E.; de Oya, M.; Lasuncion, M.A.; Gorgojo, L.; Moreno, J.M.; Benavente, M.; Cano, B.; Garces, C. Cholesterol and saturated fat intake determine the effect of polymorphisms at ABCG5/ABCG8 genes on lipid levels in children. Genet. Med. 2006, 8, 594–599. [Google Scholar] [CrossRef] [Green Version]
- Jakulj, L.; Vissers, M.N.; Tanck, M.W.; Hutten, B.A.; Stellaard, F.; Kastelein, J.J.; Dallinga-Thie, G.M. ABCG5/G8 polymorphisms and markers of cholesterol metabolism: Systematic review and meta-analysis. J. Lipid Res. 2010, 51, 3016–3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Kampen, O.; Buch, S.; Nothnagel, M.; Azocar, L.; Molina, H.; Brosch, M.; Erhart, W.; von Schonfels, W.; Egberts, J.; Seeger, M.; et al. Genetic and functional identification of the likely causative variant for cholesterol gallstone disease at the ABCG5/8 lithogenic locus. Hepatology 2013, 57, 2407–2417. [Google Scholar] [CrossRef]
- Ma, L.; Yang, J.; Runesha, H.B.; Tanaka, T.; Ferrucci, L.; Bandinelli, S.; Da, Y. Genome-wide association analysis of total cholesterol and high-density lipoprotein cholesterol levels using the Framingham heart study data. BMC Med. Genet. 2010, 11, 55. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Yin, R.X.; Wei, X.L.; Yan, T.T.; Aung, L.H.; Wu, D.F.; Wu, J.Z.; Lin, W.X.; Liu, C.W.; Pan, S.L. ATP-binding cassette transporter G5 and G8 polymorphisms and several environmental factors with serum lipid levels. PLoS ONE 2012, 7, e37972. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Li, G.B.; Yao, M.; Zhang, D.Q.; Dai, B.; Ju, C.J.; Han, M. ABCG5/8 variants are associated with susceptibility to coronary heart disease. Mol. Med. Rep. 2014, 9, 2512–2520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, A.; Emdin, C.A.; Won, H.H.; Peloso, G.M.; Natarajan, P.; Ardissino, D.; Danesh, J.; Schunkert, H.; Correa, A.; Bown, M.J.; et al. Heterozygous ATP-binding Cassette Transporter G5 Gene Deficiency and Risk of Coronary Artery Disease. Circ. Genom. Precis. Med. 2020, 13, 417–423. [Google Scholar] [CrossRef]
- Chen, Z.C.; Shin, S.J.; Kuo, K.K.; Lin, K.D.; Yu, M.L.; Hsiao, P.J. Significant association of ABCG8:D19H gene polymorphism with hypercholesterolemia and insulin resistance. J. Hum. Genet. 2008, 53, 757–763. [Google Scholar] [CrossRef] [Green Version]
- Gok, O.; Karaali, Z.E.; Acar, L.; Kilic, U.; Ergen, A. ABCG5 and ABCG8 gene polymorphisms in type 2 diabetes mellitus in the Turkish population. Can. J. Diabetes 2015, 39, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, A.; Fatima, S.; Lamri, A.; Bellili-Munoz, N.; Halimi, J.M.; Saulnier, P.J.; Hadjadj, S.; Velho, G.; Marre, M.; Roussel, R.; et al. ABCG8 polymorphisms and renal disease in type 2 diabetic patients. Metab. Clin. Exp. 2015, 64, 713–719. [Google Scholar] [CrossRef]
- Sabeva, N.S.; Rouse, E.J.; Graf, G.A. Defects in the leptin axis reduce abundance of the ABCG5-ABCG8 sterol transporter in liver. J. Biol. Chem. 2007, 282, 22397–22405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Su, K.; Sabeva, N.S.; Ji, A.; van der Westhuyzen, D.R.; Foufelle, F.; Gao, X.; Graf, G.A. GRP78 rescues the ABCG5 ABCG8 sterol transporter in db/db mice. Metab. Clin. Exp. 2015, 64, 1435–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, K.; Sabeva, N.S.; Liu, J.; Wang, Y.; Bhatnagar, S.; van der Westhuyzen, D.R.; Graf, G.A. The ABCG5 ABCG8 sterol transporter opposes the development of fatty liver disease and loss of glycemic control independently of phytosterol accumulation. J. Biol. Chem. 2012, 287, 28564–28575. [Google Scholar] [CrossRef] [Green Version]
- Van der Velde, A.E.; Vrins, C.L.J.; van den Oever, K.; Kunne, C.; Oude Elferink, R.P.J.; Kuipers, F.; Groen, A.K. Direct Intestinal Cholesterol Secretion Contributes Significantly to Total Fecal Neutral Sterol Excretion in Mice. Gastroenterology 2007, 133, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Grefhorst, A.; Verkade, H.J.; Groen, A.K. The TICE Pathway: Mechanisms and Lipid-Lowering Therapies. Methodist Debakey Cardiovasc. J. 2019, 15, 70–76. [Google Scholar]
- Nakano, T.; Inoue, I.; Murakoshi, T. A Newly Integrated Model for Intestinal Cholesterol Absorption and Efflux Reappraises How Plant Sterol Intake Reduces Circulating Cholesterol Levels. Nutrients 2019, 11, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Repa, J.J.; Berge, K.E.; Pomajzl, C.; Richardson, J.A.; Hobbs, H.; Mangelsdorf, D.J. Regulation of ATP-binding cassette sterol transporters ABCG5 and ABCG8 by the liver X receptors alpha and beta. J. Biol. Chem. 2002, 277, 18793–18800. [Google Scholar] [CrossRef] [Green Version]
- Freeman, L.A.; Kennedy, A.; Wu, J.; Bark, S.; Remaley, A.T.; Santamarina-Fojo, S.; Brewer, H.B., Jr. The orphan nuclear receptor LRH-1 activates the ABCG5/ABCG8 intergenic promoter. J. Lipid Res. 2004, 45, 1197–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumi, K.; Tanaka, T.; Uchida, A.; Magoori, K.; Urashima, Y.; Ohashi, R.; Ohguchi, H.; Okamura, M.; Kudo, H.; Daigo, K.; et al. Cooperative interaction between hepatocyte nuclear factor 4 alpha and GATA transcription factors regulates ATP-binding cassette sterol transporters ABCG5 and ABCG8. Mol. Cell Biol. 2007, 27, 4248–4260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Matozel, M.; Boehme, S.; Kong, B.; Nilsson, L.M.; Guo, G.; Ellis, E.; Chiang, J.Y. Overexpression of cholesterol 7alpha-hydroxylase promotes hepatic bile acid synthesis and secretion and maintains cholesterol homeostasis. Hepatology 2011, 53, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Einarsson, C.; Murphy, C.; Parini, P.; Bjorkhem, I.; Gafvels, M.; Eggertsen, G. Studies on LXR- and FXR-mediated effects on cholesterol homeostasis in normal and cholic acid-depleted mice. J. Lipid Res. 2006, 47, 421–430. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lu, H.; Lu, Y.F.; Lei, X.; Cui, J.Y.; Ellis, E.; Strom, S.C.; Klaassen, C.D. Potency of individual bile acids to regulate bile acid synthesis and transport genes in primary human hepatocyte cultures. Toxicol. Sci. 2014, 141, 538–546. [Google Scholar] [CrossRef] [Green Version]
- Byun, S.; Jung, H.; Chen, J.; Kim, Y.C.; Kim, D.H.; Kong, B.; Guo, G.; Kemper, B.; Kemper, J.K. Phosphorylation of hepatic farnesoid X receptor by FGF19 signaling-activated Src maintains cholesterol levels and protects from atherosclerosis. J. Biol. Chem. 2019, 294, 8732–8744. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shen, J.; Wu, T.; Kuang, J.; Liu, Q.; Cheng, S.; Pu, S.; Chen, L.; Li, R.; Li, Y.; et al. Irisin Is Controlled by Farnesoid X Receptor and Regulates Cholesterol Homeostasis. Front. Pharm. 2019, 10, 548. [Google Scholar] [CrossRef]
- Cheng, S.; Zou, M.; Liu, Q.; Kuang, J.; Shen, J.; Pu, S.; Chen, L.; Li, H.; Wu, T.; Li, R.; et al. Activation of Constitutive Androstane Receptor Prevents Cholesterol Gallstone Formation. Am. J. Pathol. 2017, 187, 808–818. [Google Scholar] [CrossRef]
- Sberna, A.L.; Assem, M.; Gautier, T.; Grober, J.; Guiu, B.; Jeannin, A.; Pais de Barros, J.P.; Athias, A.; Lagrost, L.; Masson, D. Constitutive androstane receptor activation stimulates faecal bile acid excretion and reverse cholesterol transport in mice. J. Hepatol. 2011, 55, 154–161. [Google Scholar] [CrossRef]
- Gooijert, K.E.; Havinga, R.; Wolters, H.; Wang, R.; Ling, V.; Tazuma, S.; Verkade, H.J. The mechanism of increased biliary lipid secretion in mice with genetic inactivation of bile salt export pump. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G450–G457. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.; Pijut, S.S.; Li, J.; Horn, J.; Bradford, E.M.; Leggas, M.; Barrett, T.A.; Graf, G.A. The combination of ezetimibe and ursodiol promotes fecal sterol excretion and reveals a G5G8-independent pathway for cholesterol elimination. J. Lipid Res. 2015, 56, 810–820. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.H.; Portincasa, P.; Mendez-Sanchez, N.; Uribe, M.; Wang, D.Q. Effect of ezetimibe on the prevention and dissolution of cholesterol gallstones. Gastroenterology 2008, 134, 2101–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamisako, T.; Ogawa, H.; Yamamoto, K. Effect of cholesterol, cholic acid and cholestyramine administration on the intestinal mRNA expressions related to cholesterol and bile acid metabolism in the rat. J. Gastroenterol. Hepatol. 2007, 22, 1832–1837. [Google Scholar] [CrossRef] [PubMed]
- Galman, C.; Bonde, Y.; Matasconi, M.; Angelin, B.; Rudling, M. Dramatically increased intestinal absorption of cholesterol following hypophysectomy is normalized by thyroid hormone. Gastroenterology 2008, 134, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Aleksunes, L.M.; Xu, J.; Lin, E.; Wen, X.; Goedken, M.J.; Slitt, A.L. Pregnancy represses induction of efflux transporters in livers of type I diabetic mice. Pharm. Res. 2013, 30, 2209–2220. [Google Scholar] [CrossRef]
- Biddinger, S.B.; Haas, J.T.; Yu, B.B.; Bezy, O.; Jing, E.; Zhang, W.; Unterman, T.G.; Carey, M.C.; Kahn, C.R. Hepatic insulin resistance directly promotes formation of cholesterol gallstones. Nat. Med. 2008, 14, 778–782. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, X.; Liu, H.; Guo, H.; Zhang, M.; Mei, D.; Liu, C.; He, L.; Liu, L.; Liu, X. Impaired hepatic and intestinal ATP-binding cassette transporter G5/8 was associated with high exposure of beta-sitosterol and the potential risks to blood-brain barrier integrity in diabetic rats. J. Pharm. Pharm. 2014, 66, 428–436. [Google Scholar] [CrossRef]
- Molusky, M.M.; Hsieh, J.; Lee, S.X.; Ramakrishnan, R.; Tascau, L.; Haeusler, R.A.; Accili, D.; Tall, A.R. Metformin and AMP Kinase Activation Increase Expression of the Sterol Transporters ABCG5/8 (ATP-Binding Cassette Transporter G5/G8) With Potential Antiatherogenic Consequences. Arter. Thromb. Vasc. Biol. 2018, 38, 1493–1503. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Bradfield, C.A.; Hussain, M.M. Global and hepatocyte-specific ablation of Bmal1 induces hyperlipidaemia and enhances atherosclerosis. Nat. Commun. 2016, 7, 13011. [Google Scholar] [CrossRef]
- Nishimoto, T.; Pellizzon, M.A.; Aihara, M.; Stylianou, I.M.; Billheimer, J.T.; Rothblat, G.; Rader, D.J. Fish oil promotes macrophage reverse cholesterol transport in mice. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1502–1508. [Google Scholar] [CrossRef] [Green Version]
- Kamisako, T.; Tanaka, Y.; Ikeda, T.; Yamamoto, K.; Ogawa, H. Dietary fish oil regulates gene expression of cholesterol and bile acid transporters in mice. Hepatol. Res. Off. J. Jpn. Soc. Hepatol. 2012, 42, 321–326. [Google Scholar] [CrossRef]
- Kim, E.H.; Bae, J.S.; Hahm, K.B.; Cha, J.Y. Endogenously synthesized n-3 polyunsaturated fatty acids in fat-1 mice ameliorate high-fat diet-induced non-alcoholic fatty liver disease. Biochem. Pharm. 2012, 84, 1359–1365. [Google Scholar] [CrossRef]
- De Vogel-van den Bosch, H.M.; de Wit, N.J.; Hooiveld, G.J.; Vermeulen, H.; van der Veen, J.N.; Houten, S.M.; Kuipers, F.; Muller, M.; van der Meer, R. A cholesterol-free, high-fat diet suppresses gene expression of cholesterol transporters in murine small intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1171–G1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, Y.; Hashizume, T.; Morioka, H.; Sadamitsu, S.; Ikari, A.; Miwa, M.; Sugatani, J. Diet-induced lipid accumulation in liver enhances ATP-binding cassette transporter g5/g8 expression in bile canaliculi. Drug Metab. Pharm. 2011, 26, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Apro, J.; Beckman, L.; Angelin, B.; Rudling, M. Influence of dietary sugar on cholesterol and bile acid metabolism in the rat: Marked reduction of hepatic Abcg5/8 expression following sucrose ingestion. Biochem. Biophys. Res. Commun. 2015, 461, 592–597. [Google Scholar] [CrossRef]
- Duan, L.P.; Wang, H.H.; Ohashi, A.; Wang, D.Q. Role of intestinal sterol transporters Abcg5, Abcg8, and Npc1l1 in cholesterol absorption in mice: Gender and age effects. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G269–G276. [Google Scholar] [CrossRef] [PubMed]
- Dieter, M.Z.; Maher, J.M.; Cheng, X.; Klaassen, C.D. Expression and regulation of the sterol half-transporter genes ABCG5 and ABCG8 in rats. Comp. Biochem. Physiol. C Toxicol. Pharm. 2004, 139, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, I.; Kudo, M.; Hamada, T.; Nagao, K.; Oshiro, Y.; Kato, M.; Sugawara, T.; Yamahira, T.; Ito, H.; Tamaru, S.; et al. Dietary soy protein isolate and its undigested high molecular fraction upregulate hepatic ATP-binding cassette transporter G5 and ATP-binding cassette transporter G8 mRNA and increase biliary secretion of cholesterol in rats. J. Nutr. Sci. Vitaminol. 2009, 55, 252–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, C.Y.; Sun, W.W.; Ma, Y.; Zhu, H.; Yang, P.; Wei, H.; Zeng, B.H.; Zhang, Q.; Liu, Y.; Li, W.X.; et al. Microbiota prevents cholesterol loss from the body by regulating host gene expression in mice. Sci. Rep. 2015, 5, 10512. [Google Scholar] [CrossRef] [Green Version]
- Prasnicka, A.; Cermanova, J.; Hroch, M.; Dolezelova, E.; Rozkydalova, L.; Smutny, T.; Carazo, A.; Chladek, J.; Lenicek, M.; Nachtigal, P.; et al. Iron depletion induces hepatic secretion of biliary lipids and glutathione in rats. Biochim. Biophys. Acta 2017, 1862, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.Y.; Yang, N.; Jiao, R.; Peng, C.; Guan, L.; Huang, Y.; Chen, Z.Y. Dietary calcium decreases plasma cholesterol by down-regulation of intestinal Niemann-Pick C1 like 1 and microsomal triacylglycerol transport protein and up-regulation of CYP7A1 and ABCG 5/8 in hamsters. Mol. Nutr. Food Res. 2011, 55, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Li, J.; Chao, Y.; Bi, Y.; Zhang, W.; Zhang, Y.; Ji, T.; Fu, Y.; Chen, Q.; Zhang, Q.; et al. beta-estradiol adjusts intestinal function via ERbeta and GPR30 mediated PI3K/AKT signaling activation to alleviate postmenopausal dyslipidemia. Biochem. Pharm. 2020, 180, 114134. [Google Scholar] [CrossRef]
- Li, R.; Liu, Y.; Shi, J.; Yu, Y.; Lu, H.; Yu, L.; Liu, Y.; Zhang, F. Diosgenin regulates cholesterol metabolism in hypercholesterolemic rats by inhibiting NPC1L1 and enhancing ABCG5 and ABCG8. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 1124–1133. [Google Scholar] [CrossRef]
- Graf, G.A.; Li, W.P.; Gerard, R.D.; Gelissen, I.; White, A.; Cohen, J.C.; Hobbs, H.H. Coexpression of ATP-binding cassette proteins ABCG5 and ABCG8 permits their transport to the apical surface. J. Clin. Investig. 2002, 110, 659–669. [Google Scholar] [CrossRef]
- Graf, G.A.; Yu, L.; Li, W.P.; Gerard, R.; Tuma, P.L.; Cohen, J.C.; Hobbs, H.H. ABCG5 and ABCG8 are obligate heterodimers for protein trafficking and biliary cholesterol excretion. J. Biol. Chem. 2003, 278, 48275–48282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okiyoneda, T.; Kono, T.; Niibori, A.; Harada, K.; Kusuhara, H.; Takada, T.; Shuto, T.; Suico, M.A.; Sugiyama, Y.; Kai, H. Calreticulin facilitates the cell surface expression of ABCG5/G8. Biochem. Biophys. Res. Commun. 2006, 347, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Graf, G.A.; Cohen, J.C.; Hobbs, H.H. Missense mutations in ABCG5 and ABCG8 disrupt heterodimerization and trafficking. J. Biol. Chem. 2004, 279, 24881–24888. [Google Scholar] [CrossRef] [Green Version]
- Hirata, T.; Okabe, M.; Kobayashi, A.; Ueda, K.; Matsuo, M. Molecular mechanisms of subcellular localization of ABCG5 and ABCG8. Biosci. Biotechnol. Biochem. 2009, 73, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Shuto, T.; Sato, T.; Kaneko, M.; Takada, T.; Suico, M.A.; Cyr, D.M.; Suzuki, H.; Kai, H. Inhibition of post-translational N-glycosylation by HRD1 that controls the fate of ABCG5/8 transporter. Sci. Rep. 2014, 4, 4258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, Y.; Yasui, K.; Hashizume, T.; Suto, A.; Mori, A.; Murata, Y.; Yamaguchi, M.; Ikari, A.; Sugatani, J. Involvement of a cyclic adenosine monophosphate-dependent signal in the diet-induced canalicular trafficking of adenosine triphosphate-binding cassette transporter g5/g8. Hepatology 2015, 62, 1215–1226. [Google Scholar] [CrossRef] [Green Version]
- Tachibana, S.; Hirano, M.; Hirata, T.; Matsuo, M.; Ikeda, I.; Ueda, K.; Sato, R. Cholesterol and plant sterol efflux from cultured intestinal epithelial cells is mediated by ATP-binding cassette transporters. Biosci. Biotechnol. Biochem. 2007, 71, 1886–1895. [Google Scholar] [CrossRef] [PubMed]
- Vrins, C.; Vink, E.; Vandenberghe, K.E.; Frijters, R.; Seppen, J.; Groen, A.K. The sterol transporting heterodimer ABCG5/ABCG8 requires bile salts to mediate cholesterol efflux. FEBS Lett. 2007, 581, 4616–4620. [Google Scholar] [CrossRef] [Green Version]
- Johnson, B.J.; Lee, J.Y.; Pickert, A.; Urbatsch, I.L. Bile acids stimulate ATP hydrolysis in the purified cholesterol transporter ABCG5/G8. Biochemistry 2010, 49, 3403–3411. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, D.W.; Lei, Y.; Xu, F.; Cohen, J.C.; Hobbs, H.H.; Xie, X.S. Purification and reconstitution of sterol transfer by native mouse ABCG5 and ABCG8. Biochemistry 2008, 47, 5194–5204. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Sun, F.; Zhang, D.W.; Ma, Y.; Xu, F.; Belani, J.D.; Cohen, J.C.; Hobbs, H.H.; Xie, X.S. Sterol transfer by ABCG5 and ABCG8: In vitro assay and reconstitution. J. Biol. Chem. 2006, 281, 27894–27904. [Google Scholar] [CrossRef] [Green Version]
- Muller, M.; Klein, I.; Kopacsi, S.; Remaley, A.T.; Rajnavolgyi, E.; Sarkadi, B.; Varadi, A. Co-expression of human ABCG5 and ABCG8 in insect cells generates an androstan stimulated membrane ATPase activity. FEBS Lett. 2006, 580, 6139–6144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.W.; Graf, G.A.; Gerard, R.D.; Cohen, J.C.; Hobbs, H.H. Functional asymmetry of nucleotide-binding domains in ABCG5 and ABCG8. J. Biol. Chem. 2006, 281, 4507–4516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Kinch, L.N.; Borek, D.M.; Wang, J.; Wang, J.; Urbatsch, I.L.; Xie, X.S.; Grishin, N.V.; Cohen, J.C.; Otwinowski, Z.; et al. Crystal structure of the human sterol transporter ABCG5/ABCG8. Nature 2016, 533, 561–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Grishin, N.; Kinch, L.; Cohen, J.C.; Hobbs, H.H.; Xie, X.S. Sequences in the nonconsensus nucleotide-binding domain of ABCG5/ABCG8 required for sterol transport. J. Biol. Chem. 2011, 286, 7308–7314. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.B.; Graf, G.A.; Temel, R.E. ABCG5 and ABCG8: More than a defense against xenosterols. J. Lipid Res. 2018, 59, 1103–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastida, J.M.; Benito, R.; Gonzalez-Porras, J.R.; Rivera, J. ABCG5 and ABCG8 gene variations associated with sitosterolemia and platelet dysfunction. Platelets 2020. [Google Scholar] [CrossRef]
- Wang, J.; Joy, T.; Mymin, D.; Frohlich, J.; Hegele, R.A. Phenotypic heterogeneity of sitosterolemia. J. Lipid Res. 2004, 45, 2361–2367. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Song, D.Y.; Jun, S.H.; Song, S.H.; Shin, C.H.; Ki, C.S.; Lee, K.; Song, J. High prevalence of increased sitosterol levels in hypercholesterolemic children suggest underestimation of sitosterolemia incidence. PLoS ONE 2020, 15, e0238079. [Google Scholar] [CrossRef]
- Tada, H.; Okada, H.; Nomura, A.; Yashiro, S.; Nohara, A.; Ishigaki, Y.; Takamura, M.; Kawashiri, M.A. Rare and Deleterious Mutations in ABCG5/ABCG8 Genes Contribute to Mimicking and Worsening of Familial Hypercholesterolemia Phenotype. Circ. J. 2019, 83, 1917–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lek, M.; Karczewski, K.J.; Minikel, E.V.; Samocha, K.E.; Banks, E.; Fennell, T.; O’Donnell-Luria, A.H.; Ware, J.S.; Hill, A.J.; Cummings, B.B.; et al. Analysis of protein-coding genetic variation in 60,706 humans. Nature 2016, 536, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Vauthier, V.; Ben Saad, A.; Elie, J.; Oumata, N.; Durand-Schneider, A.M.; Bruneau, A.; Delaunay, J.L.; Housset, C.; Ait-Slimane, T.; Meijer, L.; et al. Structural analogues of roscovitine rescue the intracellular traffic and the function of ER-retained ABCB4 variants in cell models. Sci. Rep. 2019, 9, 6653. [Google Scholar] [CrossRef]
- Davis, P.B.; Yasothan, U.; Kirkpatrick, P. Ivacaftor. Nat. Rev. Drug Discov. 2012, 11, 349–350. [Google Scholar] [CrossRef] [PubMed]
- Delaunay, J.L.; Durand-Schneider, A.M.; Dossier, C.; Falguieres, T.; Gautherot, J.; Davit-Spraul, A.; Ait-Slimane, T.; Housset, C.; Jacquemin, E.; Maurice, M. A functional classification of ABCB4 variations causing progressive familial intrahepatic cholestasis type 3. Hepatology 2016, 63, 1620–1631. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Typical | No Mutation | ABCG5 | ABCG8 |
|---|---|---|---|
| Class I | Nonsense, Frameshift, Deletions | 57 known or predicted | 58 known or predicted |
| Class II | Maturation | R389H, R419P, R419H, N437K | R189H, P231T, R263Q, L501P, G574E, L596R |
| Class III | Activity | ||
| Class IV | Stability | ||
| Class V | No Detectible Defect | ||
| Unclassified | Inconclusive Results | E146Q | R543S, G574R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, K.; Segard, A.; Graf, G.A. Sitosterolemia: Twenty Years of Discovery of the Function of ABCG5ABCG8. Int. J. Mol. Sci. 2021, 22, 2641. https://doi.org/10.3390/ijms22052641
Williams K, Segard A, Graf GA. Sitosterolemia: Twenty Years of Discovery of the Function of ABCG5ABCG8. International Journal of Molecular Sciences. 2021; 22(5):2641. https://doi.org/10.3390/ijms22052641
Chicago/Turabian StyleWilliams, Kori, Allison Segard, and Gregory A. Graf. 2021. "Sitosterolemia: Twenty Years of Discovery of the Function of ABCG5ABCG8" International Journal of Molecular Sciences 22, no. 5: 2641. https://doi.org/10.3390/ijms22052641
APA StyleWilliams, K., Segard, A., & Graf, G. A. (2021). Sitosterolemia: Twenty Years of Discovery of the Function of ABCG5ABCG8. International Journal of Molecular Sciences, 22(5), 2641. https://doi.org/10.3390/ijms22052641