
Quantitative Analysis of UV-B Radiation Interception in 3D Plant Structures and Intraindividual Distribution of Phenolic Contents



Abstract
:1. Introduction
2. Results
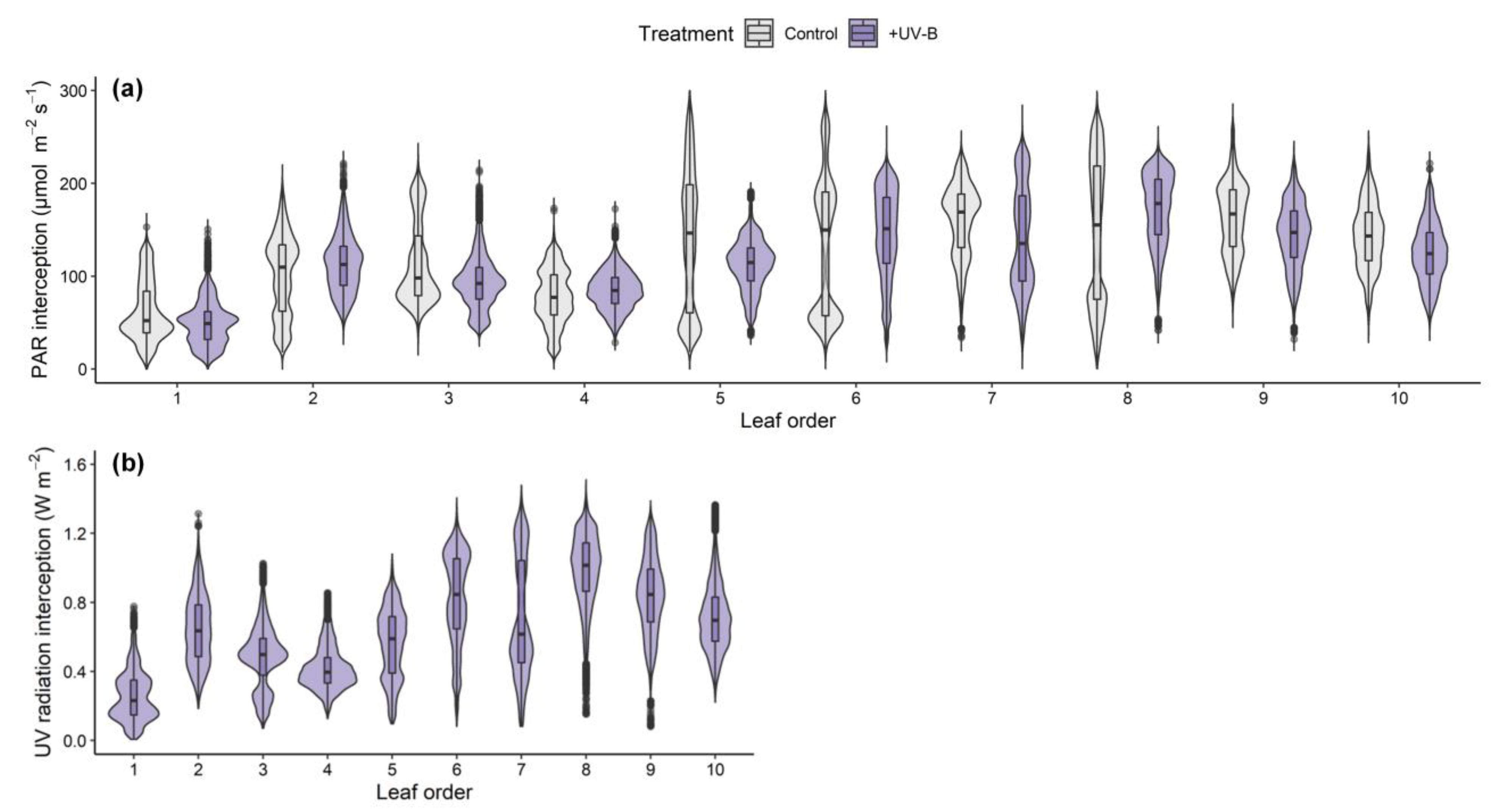
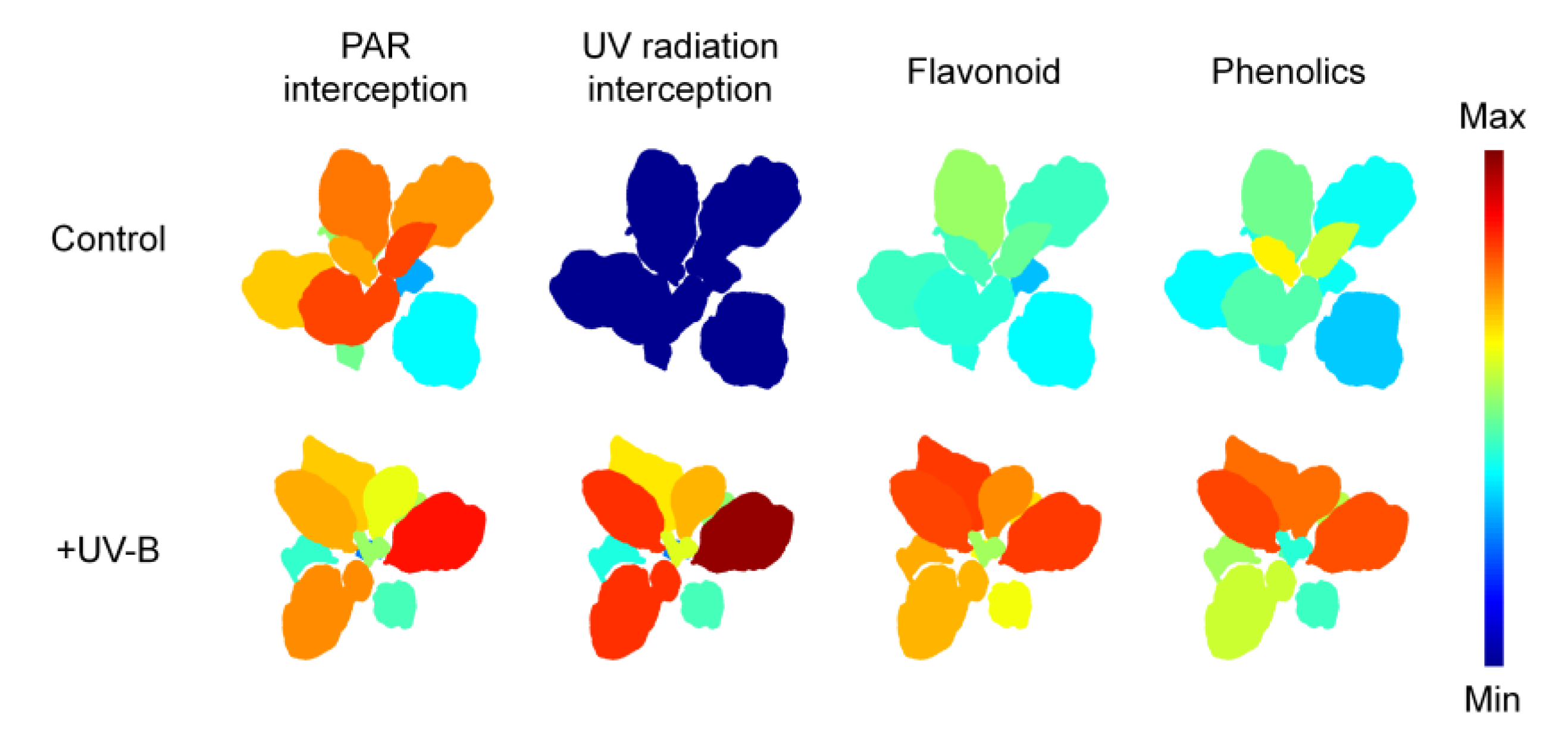
2.1. Distributions of PAR and UV-B Radiation Interceptions on Kale
2.2. Growth Characteristics of Kales According to UV-B and Leaf Order
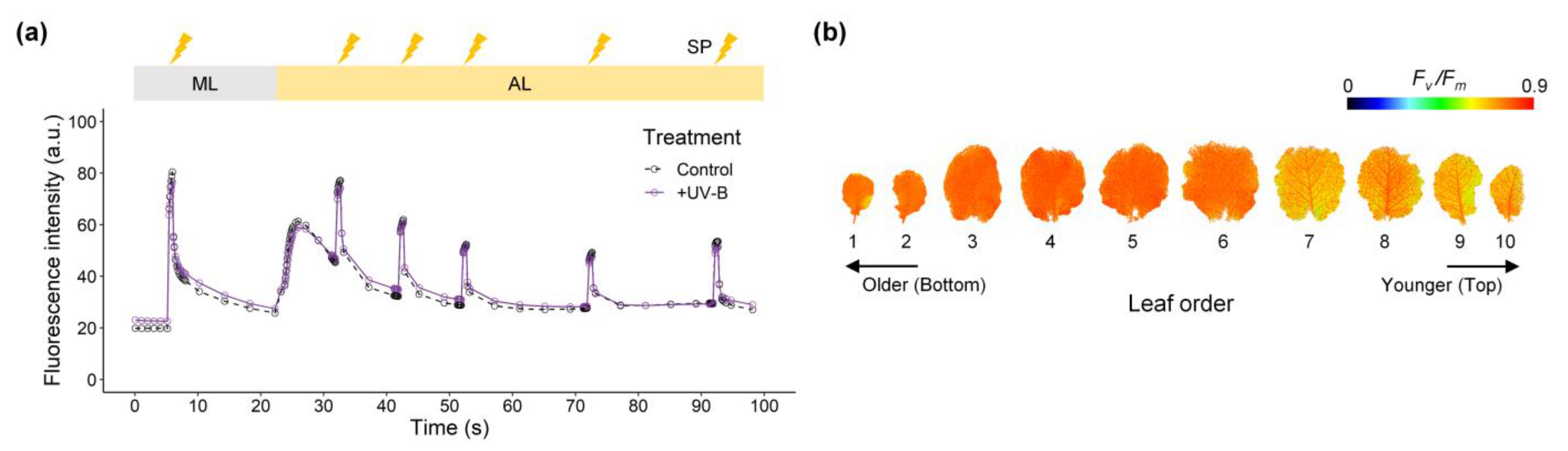
2.3. Leaf Photochemistry and Nonphotochemical Quenching
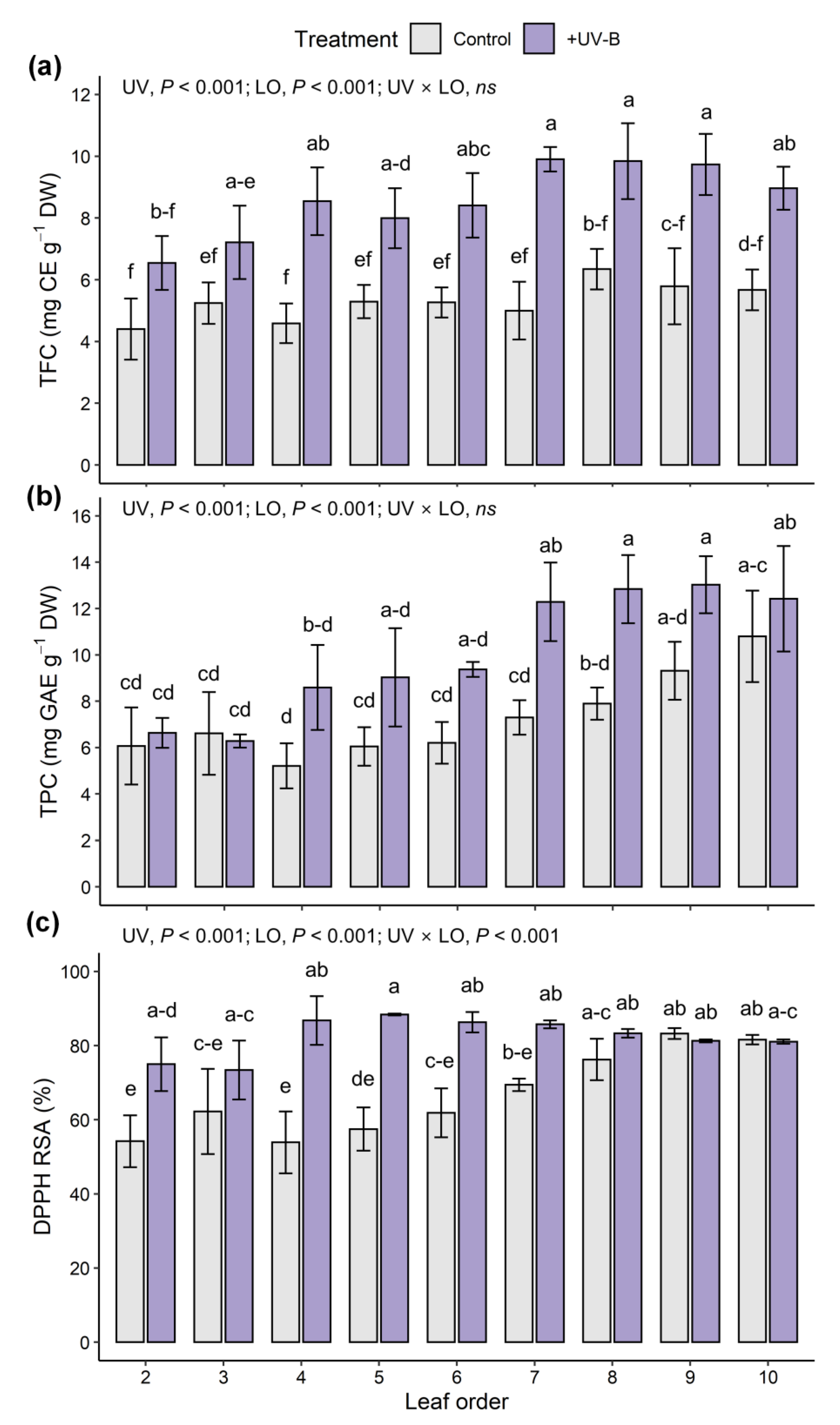
2.4. Phenolic Compounds and Antioxidant Capacity
3. Discussion
3.1. Leaf Morphology, Optical Property and UV-B Radiation Interception
3.2. The Effect of UV-B on Photosynthetic Characteristics
3.3. The Effect of UV-B Radiation on Phenolic and Flavonoid Compounds
3.4. Leaf Age as a Physiological Factor Determining UV-B Induced Phenolic Content
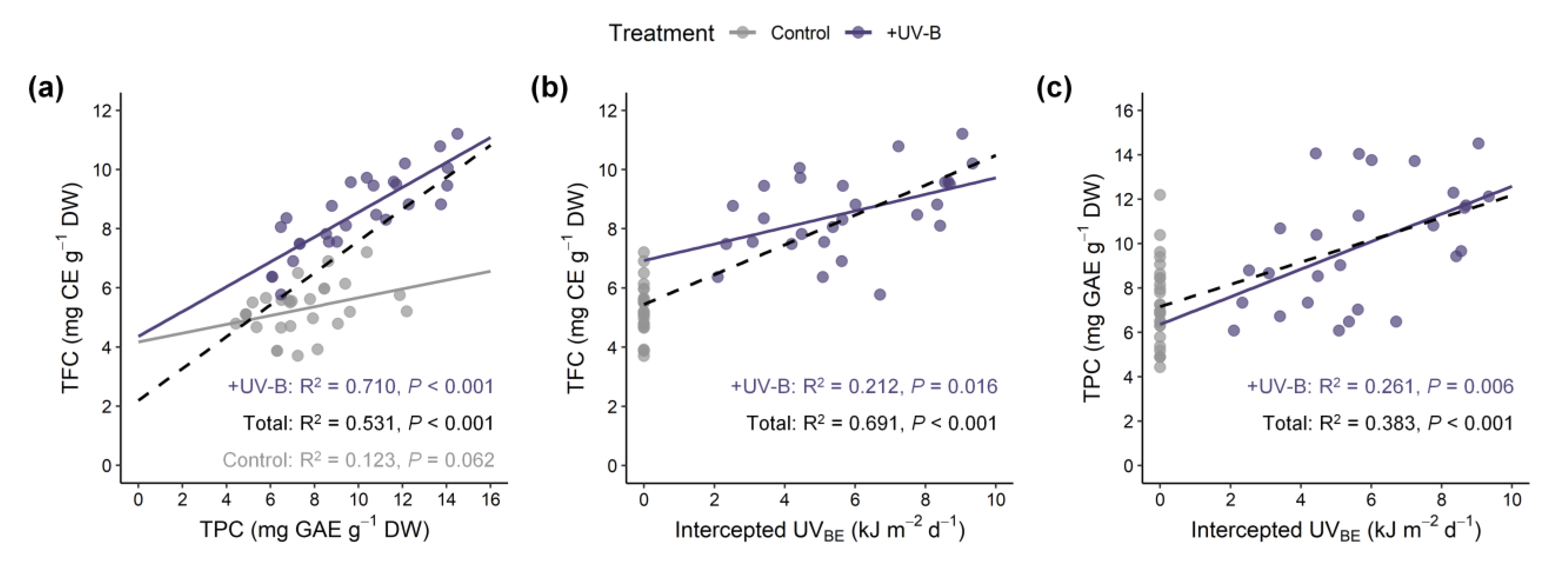
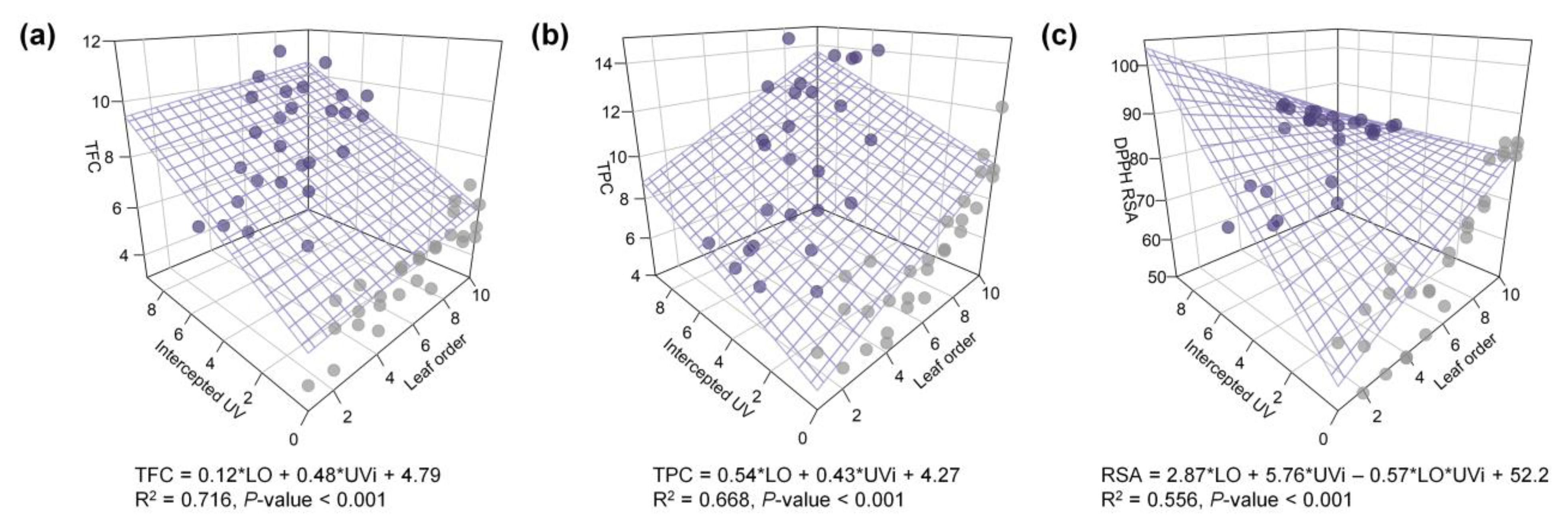
3.5. Potential for Estimating UV-B Induced Phenolic Content in 3D Plant Structure
4. Materials and Methods
4.1. Plant Material and Growth Condition
4.2. UV Treatment
4.3. Chlorophyll Fluorescence Imaging
4.4. Radiation Interception on Plant
4.4.1. D-Scanned Plant Model Generation
4.4.2. Optical Properties and Simulation Environment
4.4.3. Ray Tracing Simulation
4.5. Phenolic Compounds and Antioxidant Capacity
4.5.1. Sample Extraction
4.5.2. Total Flavonoid Content
4.5.3. Total Phenolic Content
4.5.4. Antioxidant Capacity
4.6. Plant Growth Characteristics
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Jansen, M.A.; Gaba, V.; Greenberg, B.M. Higher plants and UV-B radiation: Balancing damage, repair and acclimation. Trends Plant Sci. 1998, 3, 131–135. [Google Scholar] [CrossRef]
- Hu, Z.; Li, H.; Chen, S.; Yang, Y. Chlorophyll content and photosystem II efficiency in soybean exposed to supplemental ultraviolet-B radiation. Photosynthetica 2013, 51, 151–157. [Google Scholar] [CrossRef]
- Bashri, G.; Singh, M.; Mishra, R.K.; Kumar, J.; Singh, V.P.; Prasad, S.M. Kinetin regulates UV-B-Induced damage to growth, photosystem II photochemistry, and nitrogen metabolism in tomato seedlings. J. Plant Growth Regul. 2017, 37, 233–245. [Google Scholar] [CrossRef]
- Khudyakova, A.Y.; Kreslavski, V.D.; Shmarev, A.N.; Lyubimov, V.Y.; Shirshikova, G.N.; Pashkovskiy, P.P.; Kuznetsov, V.V.; Allakhverdiev, S.I.; Allahverdiev, S.I. Impact of UV-B radiation on the photosystem II activity, pro-/antioxidant balance and expression of light-activated genes in Arabidopsis thaliana hy4 mutants grown under light of different spectral composition. J. Photochem. Photobiol. B Biol. 2019, 194, 14–20. [Google Scholar] [CrossRef]
- Mosadegh, H.; Trivellini, A.; Lucchesini, M.; Ferrante, A.; Maggini, R.; Vernieri, P.; Sodi, A.M. UV-B Physiological changes under conditions of distress and eustress in sweet basil. Plants 2019, 8, 396. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, G.I. Signal transduction in responses to UV-B radiation. Annu. Rev. Plant Biol. 2009, 60, 407–431. [Google Scholar] [CrossRef] [PubMed]
- Hideg, É.; Jansen, M.A.; Strid, Å. UV-B exposure, ROS, and stress: Inseparable companions or loosely linked associates? Trends Plant Sci. 2013, 18, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robson, T.M.; Klem, K.; Urban, O.; Jansen, M.A.K. Re-interpreting plant morphological responses to UV-B radiation. Plant Cell Environ. 2014, 38, 856–866. [Google Scholar] [CrossRef] [PubMed]
- Takshak, S.; Agrawal, S. Defence strategies adopted by the medicinal plant Coleus forskohlii against supplemental ultraviolet-B radiation: Augmentation of secondary metabolites and antioxidants. Plant Physiol. Biochem. 2015, 97, 124–138. [Google Scholar] [CrossRef]
- Schulze, E.-D.; Beck, E.; Buchmann, N.; Clemens, S.; Müller-Hohenstein, K.; Scherer-Lorenzen, M. Light. In Plant Ecology; Springer: Berlin/Heidelberg, Germany, 2019; pp. 57–90. ISBN 978-3-662-56233-8. [Google Scholar]
- Clé, C.; Hill, L.M.; Niggeweg, R.; Martin, C.R.; Guisez, Y.; Prinsen, E.; Jansen, M.A. Modulation of chlorogenic acid biosynthesis in Solanum lycopersicum; consequences for phenolic accumulation and UV-tolerance. Phytochemistry 2008, 69, 2149–2156. [Google Scholar] [CrossRef]
- Solovchenko, A.E.; Merzlyak, M.N. Screening of visible and UV radiation as a photoprotective mechanism in plants. Russ. J. Plant Physiol. 2008, 55, 719–737. [Google Scholar] [CrossRef]
- Mewis, I.; Schreiner, M.; Nguyen, C.N.; Krumbein, A.; Ulrichs, C.; Lohse, M.; Zrenner, R. UV-B irradiation changes specifically the secondary metabolite profile in broccoli sprouts: Induced signaling overlaps with defense response to biotic stressors. Plant Cell Physiol. 2012, 53, 1546–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, C.; Yao, X.-Q.; He, X.-L.; Chu, J.-Z.; Ma, C.-H.; Shi, X.-F. Effects of enhanced UV-B radiation on biochemical traits in postharvest flowers of medicinal chrysanthemum. Photochem. Photobiol. 2015, 91, 845–850. [Google Scholar] [CrossRef]
- Takshak, S.; Agrawal, S. Defense potential of secondary metabolites in medicinal plants under UV-B stress. J. Photochem. Photobiol. B Biol. 2019, 193, 51–88. [Google Scholar] [CrossRef]
- Bilger, W.; Johnsen, T.; Schreiber, U. UV-excited chlorophyll fluorescence as a tool for the assessment of UV-protection by the epidermis of plants. J. Exp. Bot. 2001, 52, 2007–2014. [Google Scholar] [CrossRef] [PubMed]
- Hagen, S.F.; Borge, G.I.A.; Solhaug, K.A.; Bengtsson, G.B. Effect of cold storage and harvest date on bioactive compounds in curly kale (Brassica oleracea L. var. acephala). Postharvest Biol. Technol. 2009, 51, 36–42. [Google Scholar] [CrossRef]
- Dou, H.; Niu, G.; Gu, M. Pre-harvest UV-B radiation and photosynthetic photon flux density interactively affect plant photosynthesis, growth, and secondary metabolites accumulation in basil (Ocimum basilicum) plants. Agronomy 2019, 9, 434. [Google Scholar] [CrossRef] [Green Version]
- Majer, P.; Hideg, É. Developmental stage is an important factor that determines the antioxidant responses of young and old grapevine leaves under UV irradiation in a green-house. Plant Physiol. Biochem. 2012, 50, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Csepregi, K.; Coffey, A.; Cunningham, N.; Prinsen, E.; Hideg, É.; Jansen, M.A. Developmental age and UV-B exposure co-determine antioxidant capacity and flavonol accumulation in Arabidopsis leaves. Environ. Exp. Bot. 2017, 140, 19–25. [Google Scholar] [CrossRef]
- Robson, T.M.; Aphalo, P.J.; Banaś, A.K.; Barnes, P.W.; Brelsford, C.C.; Jenkins, G.I.; Kotilainen, T.K.; Łabuz, J.; Martínez-Abaigar, J.; Morales, L.O.; et al. A perspective on ecologically relevant plant-UV research and its practical application. Photochem. Photobiol. Sci. 2019, 18, 970–988. [Google Scholar] [CrossRef] [Green Version]
- Liakoura, V.; Bornman, J.F.; Karabourniotis, G. The ability of abaxial and adaxial epidermis of sun and shade leaves to attenuate UV-A and UV-B radiation in relation to the UV absorbing capacity of the whole leaf methanolic extracts. Physiol. Plant. 2002, 117, 33–43. [Google Scholar] [CrossRef]
- Tang, L.; Hou, C.; Huang, H.; Chen, C.; Zou, J.; Lin, D. Light interception efficiency analysis based on three-dimensional peach canopy models. Ecol. Inform. 2015, 30, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Lee, J.W.; Ahn, T.I.; Shin, J.H.; Park, K.S.; Son, J.E. Sweet pepper (Capsicum annuum L.) canopy photosynthesis modeling using 3D plant architecture and light ray-tracing. Front. Plant Sci. 2016, 7, 1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, D.H.; Lee, J.W.; Kang, W.H.; Hwang, I.H.; Son, J.E. Estimation of whole plant photosynthetic rate of Irwin mango under artificial and natural lights using a three-dimensional plant model and ray-tracing. Int. J. Mol. Sci. 2018, 19, 152. [Google Scholar] [CrossRef] [Green Version]
- Dornbusch, T.; Wernecke, P.; Diepenbrock, W. A method to extract morphological traits of plant organs from 3D point clouds as a database for an architectural plant model. Ecol. Model. 2007, 200, 119–129. [Google Scholar] [CrossRef]
- Paulus, S.; Schumann, H.; Kuhlmann, H.; Léon, J. High-precision laser scanning system for capturing 3D plant architecture and analysing growth of cereal plants. Biosyst. Eng. 2014, 121, 1–11. [Google Scholar] [CrossRef]
- Kim, D.; Kang, W.H.; Hwang, I.; Kim, J.; Kim, J.H.; Park, K.S.; Son, J.E. Use of structurally-accurate 3D plant models for estimating light interception and photosynthesis of sweet pepper (Capsicum annuum) plants. Comput. Electron. Agric. 2020, 177, 105689. [Google Scholar] [CrossRef]
- Kang, W.H.; Hwang, I.; Jung, D.H.; Kim, D.; Kim, J.; Kim, J.H.; Park, K.S.; Son, J.E. Time change in spatial distributions of light interception and photosynthetic rate of paprika estimated by ray-tracing simulation. Prot. Hortic. Plant Fact. 2019, 28, 279–285. [Google Scholar] [CrossRef]
- Hitz, T.; Henke, M.; Graeff-Hönninger, S.; Munz, S. Three-dimensional simulation of light spectrum and intensity within an LED growth chamber. Comput. Electron. Agric. 2019, 156, 540–548. [Google Scholar] [CrossRef]
- Kim, J.; Kang, W.H.; Son, J.E. Interpretation and evaluation of electrical lighting in plant factories with ray-tracing simulation and 3D plant modeling. Agronomy 2020, 10, 1545. [Google Scholar] [CrossRef]
- Heinze, M.; Hanschen, F.S.; Wiesner-Reinhold, M.; Baldermann, S.; Graefe, J.; Schreiner, M.; Neugart, S. Effects of developmental stages and reduced UVB and low UV conditions on plant secondary metabolite profiles in pak choi (Brassica rapa subsp. chinensis). J. Agric. Food Chem. 2018, 66, 1678–1692. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.I.; Kim, J.-S.; Kim, D.; Kim, C.Y.; Son, J.E. Harvest strategies to maximize the annual production of bioactive compounds, glucosinolates, and total antioxidant activities of kale in plant factories. Hortic. Environ. Biotechnol. 2019, 60, 883–894. [Google Scholar] [CrossRef]
- Wang, S.Y.; Lin, H.-S. Antioxidant activity in fruits and leaves of blackberry, raspberry, and strawberry varies with cultivar and developmental stage. J. Agric. Food Chem. 2000, 48, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Vagiri, M.; Conner, S.; Stewart, D.; Andersson, S.C.; Verrall, S.; Johansson, E.; Rumpunen, K. Phenolic compounds in blackcurrant (Ribes nigrum L.) leaves relative to leaf position and harvest date. Food Chem. 2015, 172, 135–142. [Google Scholar] [CrossRef]
- Lee, J.-H.; Oh, M.-M. Short-term low temperature increases phenolic antioxidant levels in kale. Hortic. Environ. Biotechnol. 2015, 56, 588–596. [Google Scholar] [CrossRef]
- Lee, J.-H.; Oh, M.-M.; Son, K.-H. Short-term ultraviolet (UV)-A light-emitting diode (LED) radiation improves biomass and bioactive compounds of kale. Front. Plant Sci. 2019, 10, 1042. [Google Scholar] [CrossRef] [Green Version]
- Yoon, H.I.; Zhang, W.; Son, J.E. Optimal duration of drought stress near harvest for promoting bioactive compounds and antioxidant capacity in kale with or without UV-B radiation in plant factories. Plants 2020, 9, 295. [Google Scholar] [CrossRef] [Green Version]
- Mosadegh, H.; Trivellini, A.; Ferrante, A.; Lucchesini, M.; Vernieri, P.; Mensuali, A. Applications of UV-B lighting to enhance phenolic accumulation of sweet basil. Sci. Hortic. 2018, 229, 107–116. [Google Scholar] [CrossRef]
- Yoon, H.I.; Kim, D.; Son, J.E. Spatial and temporal bioactive compound contents and chlorophyll fluorescence of kale (Brassica oleracea L.) under UV-B exposure near harvest time in controlled environments. Photochem. Photobiol. 2020, 96, 845–852. [Google Scholar] [CrossRef]
- Nascimento, L.B.D.S.; Brunetti, C.; Agati, G.; Iacono, C.L.; Detti, C.; Giordani, E.; Ferrini, F.; Gori, A. Short-term pre-harvest UV-B supplement enhances the polyphenol content and antioxidant capacity of Ocimum basilicum leaves during storage. Plants 2020, 9, 797. [Google Scholar] [CrossRef]
- Barnes, P.W.; Ballaré, C.L.; Caldwell, M.M. Photomorphogenic effects of UV-B radiation on plants: Consequences for light competition. J. Plant Physiol. 1996, 148, 15–20. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, M.; Li, Z.; Duan, L.; Wang, S. Enhanced UV-B radiation increases glyphosate resistance in velvetleaf (Abutilon theophrasti). Photochem. Photobiol. 2012, 88, 1428–1432. [Google Scholar] [CrossRef]
- Yang, Y.; Niu, K.; Hu, Z.; Niklas, K.J.; Sun, S. Linking species performance to community structure as affected by UV-B radiation: An attenuation experiment. J. Plant Ecol. 2017, 11, 286–296. [Google Scholar] [CrossRef]
- Cen, Y.; Bornman, J.F. The effect of exposure to enhanced UV-B radiation on the penetration of monochromatic and polychromatic UV-B radiation in leaves of Brassica napus. Physiol. Plant. 1993, 87, 249–255. [Google Scholar] [CrossRef]
- Agati, G.; Biricolti, S.; Guidi, L.; Ferrini, F.; Fini, A.; Tattini, M. The biosynthesis of flavonoids is enhanced similarly by UV radiation and root zone salinity in L. vulgare leaves. J. Plant Physiol. 2011, 168, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Strauss, S.; Lempe, J.; Prusinkiewicz, P.; Tsiantis, M.; Smith, R.S. Phyllotaxis: Is the golden angle optimal for light capture? New Phytol. 2019, 225, 499–510. [Google Scholar] [CrossRef]
- Wargent, J.J.; Jordan, B.R. From ozone depletion to agriculture: Understanding the role of UV radiation in sustainable crop production. New Phytol. 2013, 197, 1058–1076. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Lüscher, J.; Morales, F.; Delrot, S.; Sánchez-Díaz, M.; Gomès, E.; Aguirreolea, J.; Pascual, I. Short- and long-term physiological responses of grapevine leaves to UV-B radiation. Plant Sci. 2013, 213, 114–122. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Morales, F.; Delrot, S.; Sánchez-Díaz, M.; Gomès, E.; Aguirreolea, J.; Pascual, I. Characterization of the adaptive response of grapevine (cv. Tempranillo) to UV-B radiation under water deficit conditions. Plant Sci. 2015, 232, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Tilbrook, K.; Arongaus, A.B.; Binkert, M.; Heijde, M.; Yin, R.; Ulm, R. The UVR8 UV-B photoreceptor: Perception, signaling and response. Arab. Book 2013, 11, e0164. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Agrawal, S.; Agrawal, M. UVR8 mediated plant protective responses under low UV-B radiation leading to photosynthetic acclimation. J. Photochem. Photobiol. B Biol. 2014, 137, 67–76. [Google Scholar] [CrossRef]
- Allorent, G.; Lefebvre-Legendre, L.; Chappuis, R.; Kuntz, M.; Truong, T.B.; Niyogi, K.K.; Ulm, R.; Goldschmidt-Clermont, M. UV-B photoreceptor-mediated protection of the photosynthetic machinery in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2016, 113, 14864–14869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.-J.; Son, J.E.; Oh, M.-M. Growth and phenolic compounds of Lactuca sativa L. grown in a closed-type plant production system with UV-A, -B, or -C lamp. J. Sci. Food Agric. 2014, 94, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Inostroza-Blancheteau, C.; Reyes-Díaz, M.; Arellano, A.; Latsague, M.; Acevedo, P.; Loyola, R.; Arce-Johnson, P.; Alberdi, M. Effects of UV-B radiation on anatomical characteristics, phenolic compounds and gene expression of the phenylpropanoid pathway in highbush blueberry leaves. Plant Physiol. Biochem. 2014, 85, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ma, Y.; Weng, Y.; Yang, R.; Gu, Z.; Wang, P. Effects of UV-B radiation on phenolic accumulation, antioxidant activity and physiological changes in wheat (Triticum aestivum L.) seedlings. Food Biosci. 2019, 30, 100409. [Google Scholar] [CrossRef]
- Matsuura, H.N.; de Costa, F.; Yendo, A.C.A.; Fett-Neto, A.G. Photoelicitation of bioactive secondary metabolites by ultraviolet radiation: Mechanisms, strategies, and applications. In Biotechnology for Medicinal Plants; Chandra, S., Lata, H., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 9783642299, pp. 171–190. ISBN 978-3-642-29973-5. [Google Scholar]
- Vidovic, M.; Morina, F.; Milić, S.; Zechmann, B.; Albert, A.; Winkler, J.B.; Jovanovic, S.V. Ultraviolet-B component of sunlight stimulates photosynthesis and flavonoid accumulation in variegated Plectranthus coleoides leaves depending on background light. Plant Cell Environ. 2014, 38, 968–979. [Google Scholar] [CrossRef]
- Reifenrath, K.; Müller, C. Species-specific and leaf-age dependent effects of ultraviolet radiation on two Brassicaceae. Phytochemistry 2007, 68, 875–885. [Google Scholar] [CrossRef]
- Sun, M.; Gu, X.; Fu, H.; Zhang, L.; Chen, R.; Cui, L.; Zheng, L.; Zhang, D.; Tian, J. Change of secondary metabolites in leaves of Ginkgo biloba L. in response to UV-B induction. Innov. Food Sci. Emerg. Technol. 2010, 11, 672–676. [Google Scholar] [CrossRef]
- Reuber, S.; Bornman, J.F.; Weissenbock, G. A flavonoid mutant of barley (Hordeum vulgare L.) exhibits increased sensitivity to UV-B radiation in the primary leaf. Plant Cell Environ. 1996, 19, 593–601. [Google Scholar] [CrossRef]
- Neugart, S.; Zietz, M.; Schreiner, M.; Rohn, S.; Kroh, L.W.; Krumbein, A. Structurally different flavonol glycosides and hydroxycinnamic acid derivatives respond differently to moderate UV-B radiation exposure. Physiol. Plant. 2012, 145, 582–593. [Google Scholar] [CrossRef]
- Hofmann, R.W.; Campbell, B.D.; Fountain, D.W.; Jordan, B.R.; Greer, D.H.; Hunt, D.Y.; Hunt, C.L. Multivariate analysis of intraspecific responses to UV-B radiation in white clover (Trifolium repens L.). Plant Cell Environ. 2001, 24, 917–927. [Google Scholar] [CrossRef]
- Flint, S.D.; Caldwell, M.M. A biological spectral weighting function for ozone depletion research with higher plants. Physiol. Plant. 2003, 117, 137–144. [Google Scholar] [CrossRef]
- Cen, H.; Weng, H.; Yao, J.; He, M.; Lv, J.; Hua, S.; Li, H.; He, Y. Chlorophyll fluorescence imaging uncovers photosynthetic fingerprint of Citrus Huanglongbing. Front. Plant Sci. 2017, 8, 1509. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Sun, D.; Cen, H.; Xu, H.; Weng, H.; Yuan, F.; He, Y. Phenotyping of Arabidopsis drought stress response using kinetic chlorophyll fluorescence and multicolor fluorescence imaging. Front. Plant Sci. 2018, 9, 603. [Google Scholar] [CrossRef] [PubMed]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Ainsworth, E.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Andarwulan, N.; Batari, R.; Sandrasari, D.A.; Bolling, B.; Wijaya, H. Flavonoid content and antioxidant activity of vegetables from Indonesia. Food Chem. 2010, 121, 1231–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Order 1 | Leaf Fresh Weight (g) | Leaf Dry Weight (g) | Leaf Area (cm2) | |||
|---|---|---|---|---|---|---|
| Control | +UV-B | Control | +UV-B | Control | +UV-B | |
| 2 | 2.36 ± 1.54 2 | 2.66 ± 0.40 | 0.18 ± 0.11 | 0.21 ± 0.01 | 32.25 ± 9.65 | 45.23 ± 4.16 |
| 3 | 5.15 ± 2.84 | 4.87 ± 1.39 | 0.41 ± 0.26 | 0.38 ± 0.08 | 84.49 ± 49.35 | 87.24 ± 28.17 |
| 4 | 8.31 ± 1.55 | 8.57 ± 0.99 | 0.70 ± 0.20 | 0.82 ± 0.19 | 168.31 ± 34.38 | 155.03 ± 13.80 |
| 5 | 8.97 ± 1.33 | 8.44 ± 1.25 | 0.88 ± 0.14 | 0.86 ± 0.15 | 179.44 ± 16.96 | 186.87 ± 14.14 |
| 6 | 10.17 ± 2.56 | 8.62 ± 0.69 | 0.93 ± 0.33 | 0.89 ± 0.17 | 200.07 ± 36.42 | 178.14 ± 13.40 |
| 7 | 10.12 ± 1.09 | 7.06 ± 1.97 | 0.96 ± 0.21 | 0.79 ± 0.29 | 182.72 ± 19.97 | 147.06 ± 28.89 |
| 8 | 7.40 ± 1.76 | 5.80 ± 1.54 | 0.77 ± 0.21 | 0.66 ± 0.18 | 139.49 ± 17.85 | 115.14 ± 33.49 |
| 9 | 5.38 ± 0.86 | 4.18 ± 1.05 | 0.57 ± 0.09 | 0.50 ± 0.12 | 91.20 ± 1.82 | 64.23 ± 19.42 |
| 10 | 3.59 ± 0.53 | 2.82 ± 0.50 | 0.43 ± 0.08 | 0.37 ± 0.06 | 49.94 ± 6.36 | 37.74 ± 14.75 |
| UV treatment | * 3 | ns | * | |||
| Leaf order | *** | *** | *** | |||
| UV × Leaf order | ns | ns | ns | |||
| Treatment | Total Growth Parameter | Total Radiation Interception Per Plant | |||
|---|---|---|---|---|---|
| Fresh Weight (g) | Dry Weight (g) | Leaf Area (cm2) | PAR 2 (mmol Plant−1 d−1) | UVBE 3 (MJ Plant−1 d−1) | |
| Control | 79.5 ± 14.0 1 | 7.39 ± 1.83 | 1142.5 ± 184.8 | 914.33 ± 209.50 | - |
| +UV-B | 67.8 ± 6.3 | 6.91 ± 0.94 | 1006.0 ± 67.6 | 739.48 ± 102.24 | 2.91 ± 0.47 |
| Leaf Order 1 | Fv/Fm | ΦPSII | NPQ | |||
|---|---|---|---|---|---|---|
| Control | +UV-B | Control | +UV-B | Control | +UV-B | |
| 2 | 0.76 ab 2 | 0.77 a | 0.57 abc | 0.61 ab | 0.49 b | 0.40 b |
| 3 | 0.78 a | 0.78 a | 0.62 a | 0.58 abc | 0.39 b | 0.45 b |
| 4 | 0.78 a | 0.78 a | 0.56 abcd | 0.52 bcde | 0.65 b | 0.47 b |
| 5 | 0.78 a | 0.77 a | 0.50 cdef | 0.47 defg | 0.45 b | 0.47 b |
| 6 | 0.78 a | 0.76 a | 0.46 efg | 0.43 fg | 0.43 b | 0.55 b |
| 7 | 0.74 abc | 0.71 abcd | 0.44 efg | 0.42 fg | 0.45 b | 0.71 ab |
| 8 | 0.74 abc | 0.68 bcd | 0.44 fg | 0.42 fg | 0.49 b | 0.57 b |
| 9 | 0.71 abcd | 0.67 cd | 0.45 efg | 0.41 g | 0.49 b | 0.85 ab |
| 10 | 0.72 abcd | 0.66 d | 0.48 efg | 0.41 g | 0.57 b | 1.27 a |
| UV treatment | ** | * | ** | |||
| Leaf order | *** | *** | ** | |||
| UV × Leaf order | ns | ns | * | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, H.I.; Kim, H.Y.; Kim, J.; Oh, M.-M.; Son, J.E. Quantitative Analysis of UV-B Radiation Interception in 3D Plant Structures and Intraindividual Distribution of Phenolic Contents. Int. J. Mol. Sci. 2021, 22, 2701. https://doi.org/10.3390/ijms22052701
Yoon HI, Kim HY, Kim J, Oh M-M, Son JE. Quantitative Analysis of UV-B Radiation Interception in 3D Plant Structures and Intraindividual Distribution of Phenolic Contents. International Journal of Molecular Sciences. 2021; 22(5):2701. https://doi.org/10.3390/ijms22052701
Chicago/Turabian StyleYoon, Hyo In, Hyun Young Kim, Jaewoo Kim, Myung-Min Oh, and Jung Eek Son. 2021. "Quantitative Analysis of UV-B Radiation Interception in 3D Plant Structures and Intraindividual Distribution of Phenolic Contents" International Journal of Molecular Sciences 22, no. 5: 2701. https://doi.org/10.3390/ijms22052701
APA StyleYoon, H. I., Kim, H. Y., Kim, J., Oh, M. -M., & Son, J. E. (2021). Quantitative Analysis of UV-B Radiation Interception in 3D Plant Structures and Intraindividual Distribution of Phenolic Contents. International Journal of Molecular Sciences, 22(5), 2701. https://doi.org/10.3390/ijms22052701