
Functional Characteristics Analysis of Dehydrins in Larix kaempferi under Osmotic Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Transcriptome Based Screening and Cloning of DHN Genes in Larix Kaempferi
2.2. Alignment Analysis of DHNs in Plants
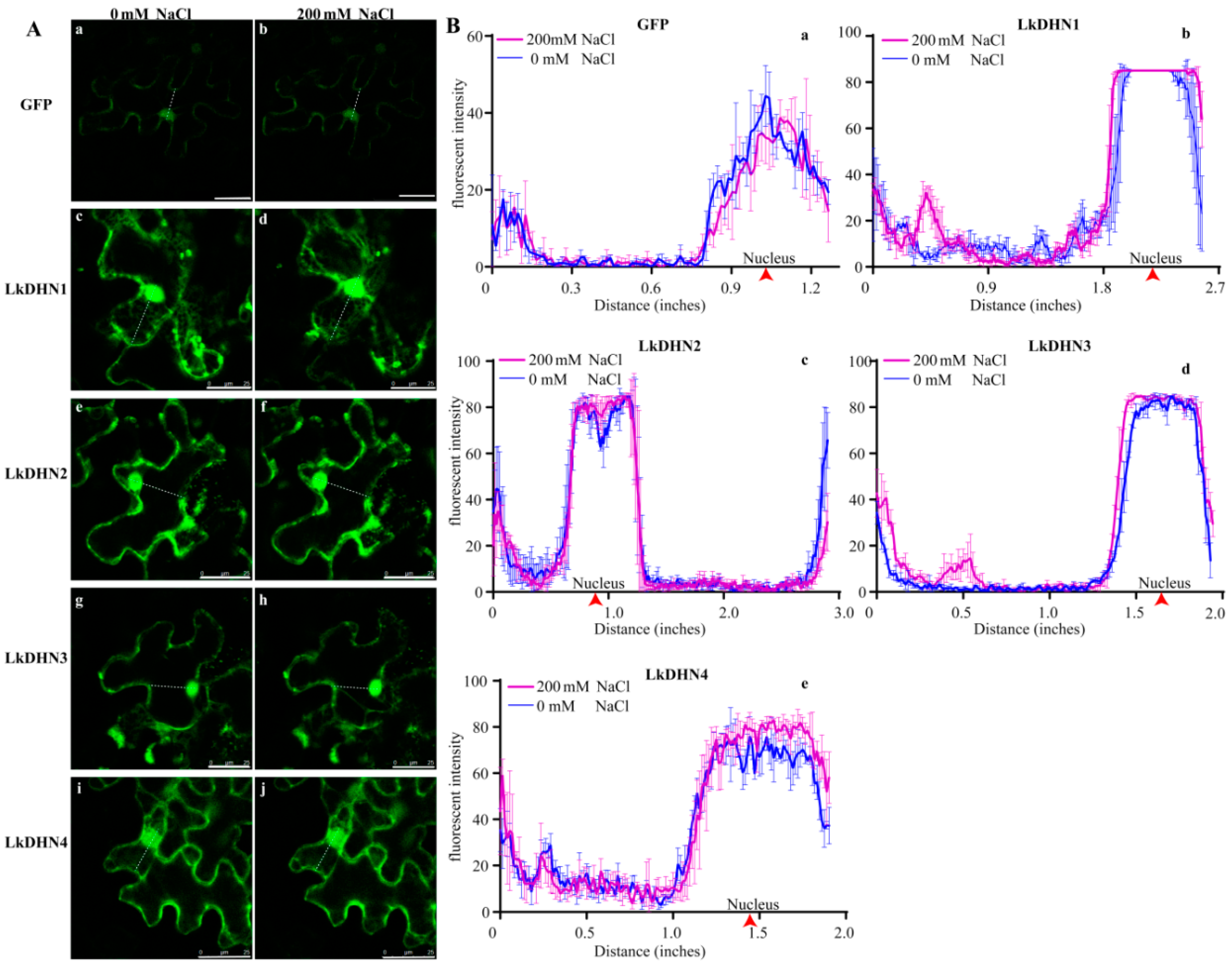
2.3. Subcellular Localization of LkDHNs
2.4. Stress Treatment Enhanced the Signal of LkDHNs in Nucleus
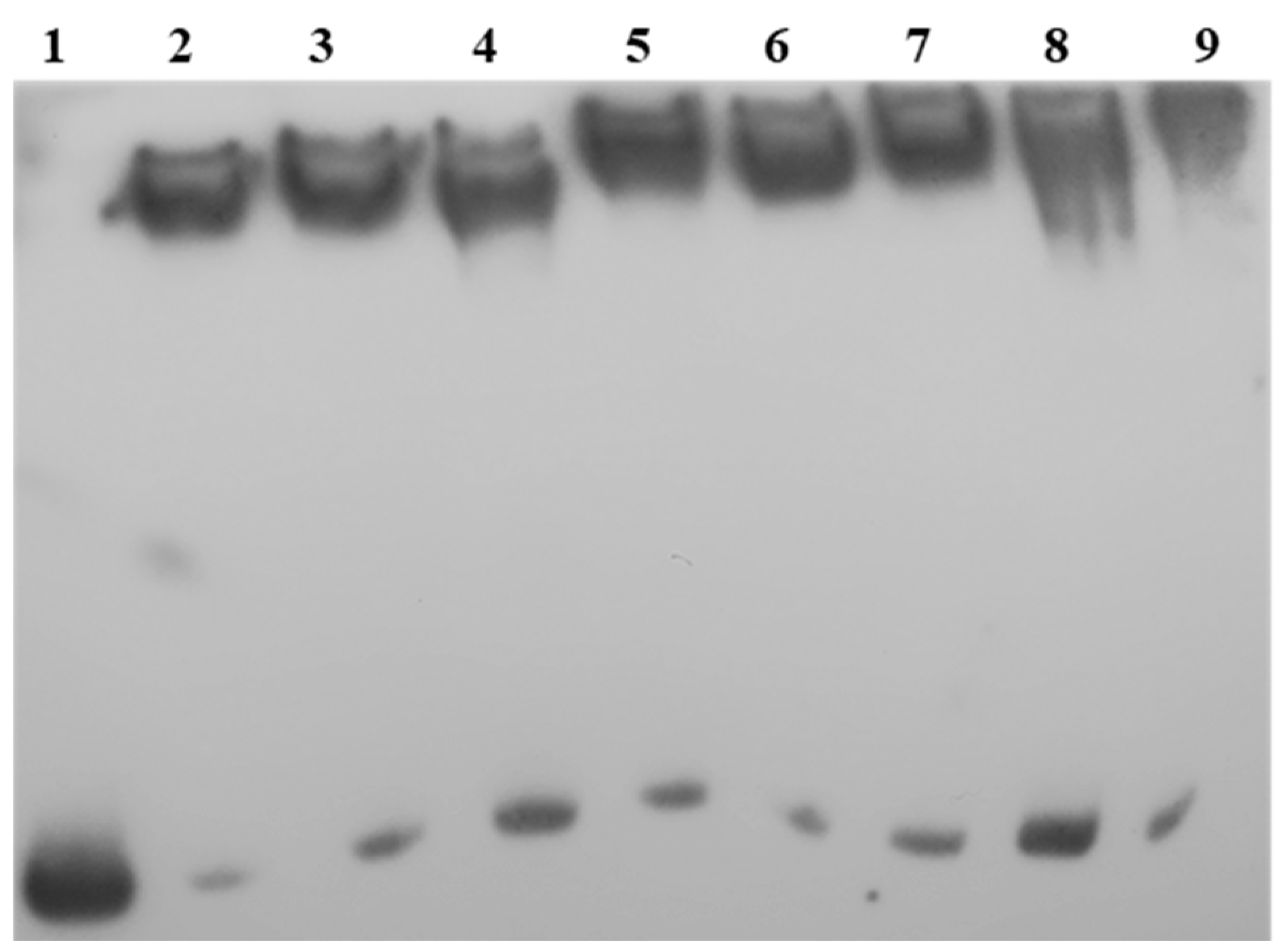
2.5. LkDHNs Bind Double-Stranded DNAs
2.6. Overexpression of LkDHNs Increased the Tolerance of Tobacco Protoplasts and Yeasts to Osmotic Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Transcriptome Based Identification of DHN Genes in L. kaempferi
4.3. DHN Sequence Alignments
4.4. Subcellular Location of LkDHNs
4.5. The Binding Ability Analysis of LkDHNs and DNA by Electrophoretic Mobility Shift Assay
4.6. Preparation of Tobacco Protoplasts by Enzymatic Hydrolysis
4.7. Stress Resistance Assay of LkDHNs in Yeast Transformants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BMGY | Buffered complex glycerol medium |
| BMMY | Buffered methanol complex medium |
| BSA | Bovine albumin |
| CSE | Caffeoyl shikimate esterase |
| DAPI | 4,6-diamidino-2-phenylindole |
| dsDNAs | Double-stranded DNAs |
| EMSA | Electrophoretic mobility shift assay |
| GFP | Green fluorescent protein |
| LB | Lysogeny broth medium |
| LEA | Late embryogenesis abundant protein |
| MES | 2-(N-morpholino) ethanesulfonicacid |
| NLS | nuclear localization signals |
| ROS | Reactive oxygen species |
| YPD | Yeast extract-peptone-dextrose |
References
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.K.; Arora, S. Role of Secondary Metabolites and Brassinosteroids in Plant Defense Against Environmental Stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Akram, N.A.; Ashraf, M.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Plant responses to environmental stresses—from gene to biotechnology. AoB Plants 2017, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, J.; Dennsteele, C.V.; Iii, L.D. Sequence and characterization of 6 Lea proteins and their genes from cotton. Plant Mol. Biol. 1988, 11, 277–291. [Google Scholar] [CrossRef]
- Iii, L.D.; Greenway, S.C.; Galau, G.A. Developmental biochemistry of cottonseed embryogenesis and germination: Changing messenger ribonucleic acid populations as shown by in vitro and in vivo protein synthesis. Biochemistry 1981, 20, 4162. [Google Scholar]
- Jensen, A.B.; Goday, A.; Figueras, M.; Jessop, A.C.; Pages, M. Phosphorylation mediates the nuclear targeting of the maize Rab17 protein. Plant J. 1998, 13, 691–697. [Google Scholar] [CrossRef] [Green Version]
- Garayarroyo, A.; Colmeneroflores, J.M.; Garciarrubio, A.; Covarrubias, A.A. Highly hydrophilic proteins in prokaryotes and eukaryotes are common during conditions of water deficit. J. Biol. Chem. 2000, 275, 5668–5674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perdiguero, P.; Collada, C.; Soto, Ã.L. Novel dehydrins lacking complete K-segments in Pinaceae. The exception rather than the rule. Front. plant Sci. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Malik, A.A.; Veltri, M.; Boddington, K.F.; Singh, K.K.; Graether, S.P. Genome Analysis of Conserved Dehydrin Motifs in Vascular Plants. Front. Plant Sci. 2017, 8, 709. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Terashima, S.; Kuboi, T. Characterization and cryoprotective activity of cold-responsive dehydrin from Citrus unshiu. J. Plant Physiol. 2001, 158, 1333–1339. [Google Scholar] [CrossRef]
- Hughes, S.L.; Schart, V.; Malcolmson, J.; Hogarth, K.A.; Martynowicz, D.M.; Tralman-Baker, E.; Patel, S.N.; Graether, S.P. The Importance of Size and Disorder in the Cryoprotective Effects of Dehydrins. Plant Physiol. 2013, 163, 1376–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Close, T.J. Dehydrins: Emergence of a biochemical role of a family of plant dehydration proteins. Physiol. Plant 1996, 97, 795–803. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Zhang, T.; Yang, X.; Li, D. Functional characterization of KS-type dehydrin ZmDHN13 and its related conserved domains under oxidative stress. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Falavigna, V.D.S.; Malabarba, J.; Silveira, C.P.; Buffon, V.; Mariath, J.E.A.; Pasquali, G.; Margis-Pinheiro, M.; Revers, L.F. Characterization of the nucellus-specific dehydrin MdoDHN11 demonstrates its involvement in the tolerance to water deficit. Plant Cell Rep. 2019, 38, 1099–1107. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yu, C.; Li, H.; Ouyang, B.; Wang, T.; Zhang, J.; Wang, X.; Ye, Z. Overexpression of ShDHN, a dehydrin gene from Solanum habrochaites enhances tolerance to multiple abiotic stresses in tomato. Plant Sci. 2015, 231, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Monna, S.; Murata, T.; Nakano, T.; Amano, S.; Nachbar, M.; WãTzig, H. The Arabidopsis KS-type dehydrin recovers lactate dehydrogenase activity inhibited by copper with the contribution of His residues. Plant Sci. 2016, 245, 135–142. [Google Scholar] [CrossRef]
- Hara, M.; Shinoda, Y.; Tanaka, Y.; Kuboi, T. DNA binding of citrus dehydrin promoted by zinc ion. Plant Cell Environ. 2009, 32, 532–541. [Google Scholar] [CrossRef]
- Lin, C.-H.; Peng, P.-H.; Ko, C.-Y.; Markhart, A.H.; Lin, T.-Y. Characterization of a Novel Y2K-type Dehydrin VrDhn1 from Vigna radiata. Plant Cell Physiol. 2012, 53, 930–942. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, P.; Indoliya, Y.; Singh, P.K.; Singh, P.C.; Chauhan, P.S.; Pande, V.; Chakrabarty, D. Role of dehydrin-FK506-binding protein complex in enhancing drought tolerance through the ABA-mediated signaling pathway. Environ. Exp. Bot. 2019, 158, 136–149. [Google Scholar] [CrossRef]
- Eriksson, S.; Eremina, N.; Barth, A.; Danielsson, J.; Harryson, P. Membrane-Induced Folding of the Plant Stress Dehydrin Lti30. Plant Physiol. 2016, 171, 932–943. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Marzinek, J.K.; Jefferies, D.; Bond, P.J.; Harryson, P.; Wohland, T. The disordered plant dehydrin Lti30 protects the membrane during water-related stress by cross-linking lipids. J. Biol. Chem. 2019, 294, 6468–6482. [Google Scholar] [CrossRef] [Green Version]
- Schulp, C.J.E.; Nabuurs, G.-J.; Verburg, P.H.; de Waal, R.W. Effect of tree species on carbon stocks in forest floor and mineral soil and implications for soil carbon inventories. For. Ecol. Manag. 2008, 256, 482–490. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, X.; Zhang, L. Structural and Functional Dynamics of Dehydrins: A Plant Protector Protein under Abiotic Stress. Int. J. Mol. Sci. 2018, 19, 3420. [Google Scholar] [CrossRef] [Green Version]
- Maszkowska, J.; Dębski, J.; Kulik, A.; Kistowski, M.; Bucholc, M.; Lichocka, M.; Klimecka, M.; Sztatelman, O.; Szymańska, K.P.; Dadlez, M.; et al. Phosphoproteomic analysis reveals that dehydrins ERD10 and ERD14 are phosphorylated by SNF1-related protein kinase 2.10 in response to osmotic stress. Plant Cell Environ. 2018. [Google Scholar] [CrossRef]
- Luo, D.; Hou, X.; Zhang, Y.; Meng, Y.; Zhang, H.; Liu, S.; Wang, X.; Chen, R. CaDHN5, a dehydrin gene from pepper, plays an important role in salt and osmotic stress responses. Int. J. Mol. Sci. 2019, 20, 1989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-C.; Kim, Y.-H.; Jeong, J.C.; Kim, C.Y.; Lee, H.-S.; Bang, J.-W.; Kwak, S.-S. Sweetpotato late embryogenesis abundant 14 (IbLEA14) gene influences lignification and increases osmotic- and salt stress-tolerance of transgenic calli. Planta 2011, 233, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Welin, B.V.; Olson, Å.; Palva, E.T. Structure and organization of two closely related low-temperature-induced dhn/lea/rab-like genes in Arabidopsis thaliana L. Heynh. Plant Mol. Biol. 1995, 29, 391–395. [Google Scholar] [CrossRef]
- Shekhawat, U.K.S.; Srinivas, L.; Ganapathi, T.R. MusaDHN-1, a novel multiple stress-inducible SK3-type dehydrin gene, contributes affirmatively to drought- and salt-stress tolerance in banana. Planta 2011, 234, 915. [Google Scholar] [CrossRef]
- Wang, X.; Chao, N.; Zhang, M.; Jiang, X.; Gai, Y. Functional characteristics of caffeoyl shikimate esterase in Larix kaempferi and monolignol biosynthesis in gymnosperms. Int. J. Mol. Sci. 2019, 20, 6071. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Li, Y.; Lu, C.; Tang, Y.; Jiang, X.; Gai, Y. Isolation and characterization of Populus xyloglucan endotransglycosylase/hydrolase (XTH) involved in osmotic stress responses. Int. J. Biol. Macromol. 2020, 155, 1277–1287. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zhang, M.; Xie, B.; Jiang, X.; Gai, Y. Functional Characteristics Analysis of Dehydrins in Larix kaempferi under Osmotic Stress. Int. J. Mol. Sci. 2021, 22, 1715. https://doi.org/10.3390/ijms22041715
Wang X, Zhang M, Xie B, Jiang X, Gai Y. Functional Characteristics Analysis of Dehydrins in Larix kaempferi under Osmotic Stress. International Journal of Molecular Sciences. 2021; 22(4):1715. https://doi.org/10.3390/ijms22041715
Chicago/Turabian StyleWang, Xuechun, Meng Zhang, Baohui Xie, Xiangning Jiang, and Ying Gai. 2021. "Functional Characteristics Analysis of Dehydrins in Larix kaempferi under Osmotic Stress" International Journal of Molecular Sciences 22, no. 4: 1715. https://doi.org/10.3390/ijms22041715
APA StyleWang, X., Zhang, M., Xie, B., Jiang, X., & Gai, Y. (2021). Functional Characteristics Analysis of Dehydrins in Larix kaempferi under Osmotic Stress. International Journal of Molecular Sciences, 22(4), 1715. https://doi.org/10.3390/ijms22041715