
Supplementing with L-Tryptophan Increases Medium Protein and Alters Expression of Genes and Proteins Involved in Milk Protein Synthesis and Energy Metabolism in Bovine Mammary Cells


 ,
, 
 and
and
Abstract
:1. Introduction
2. Results
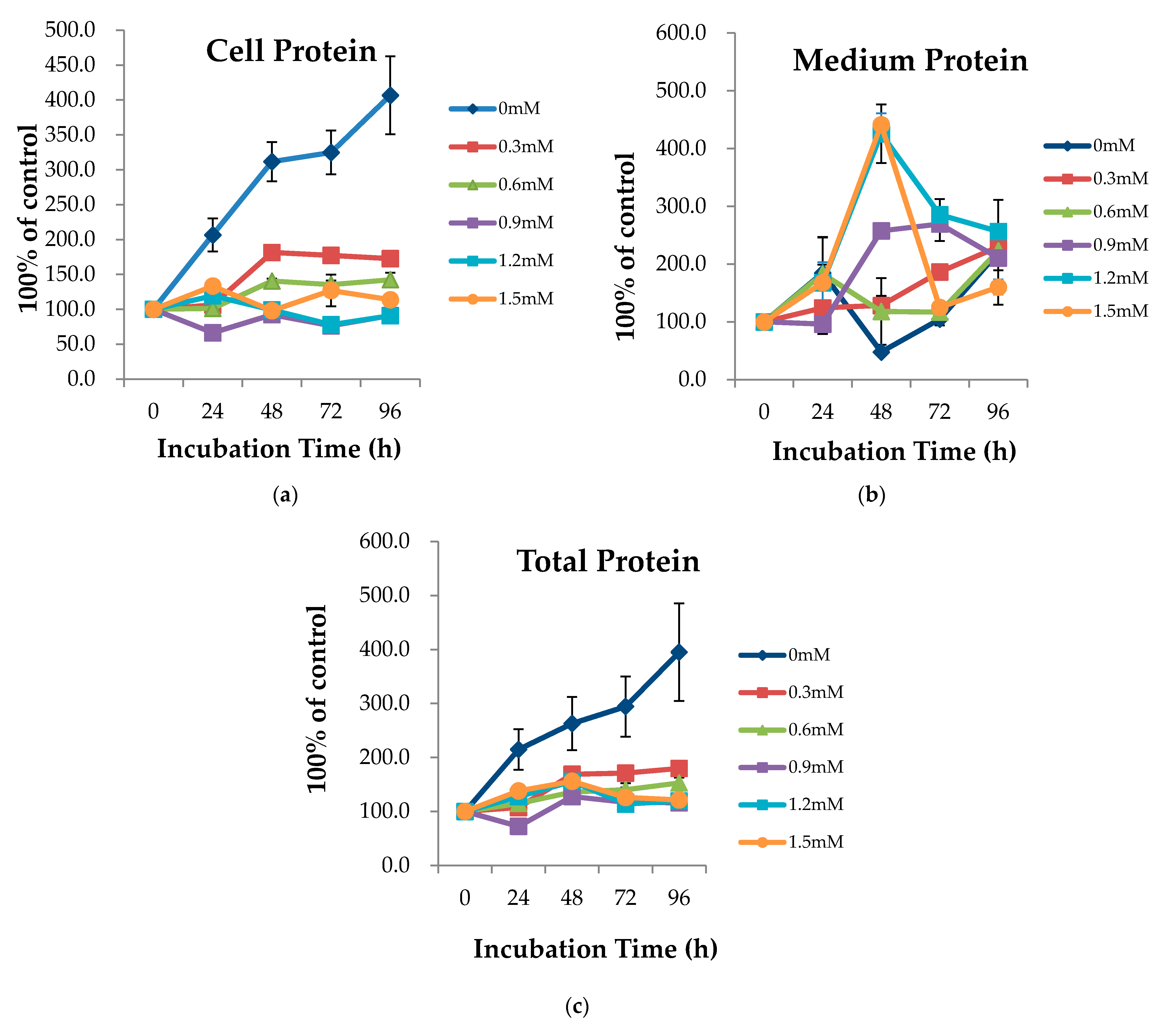
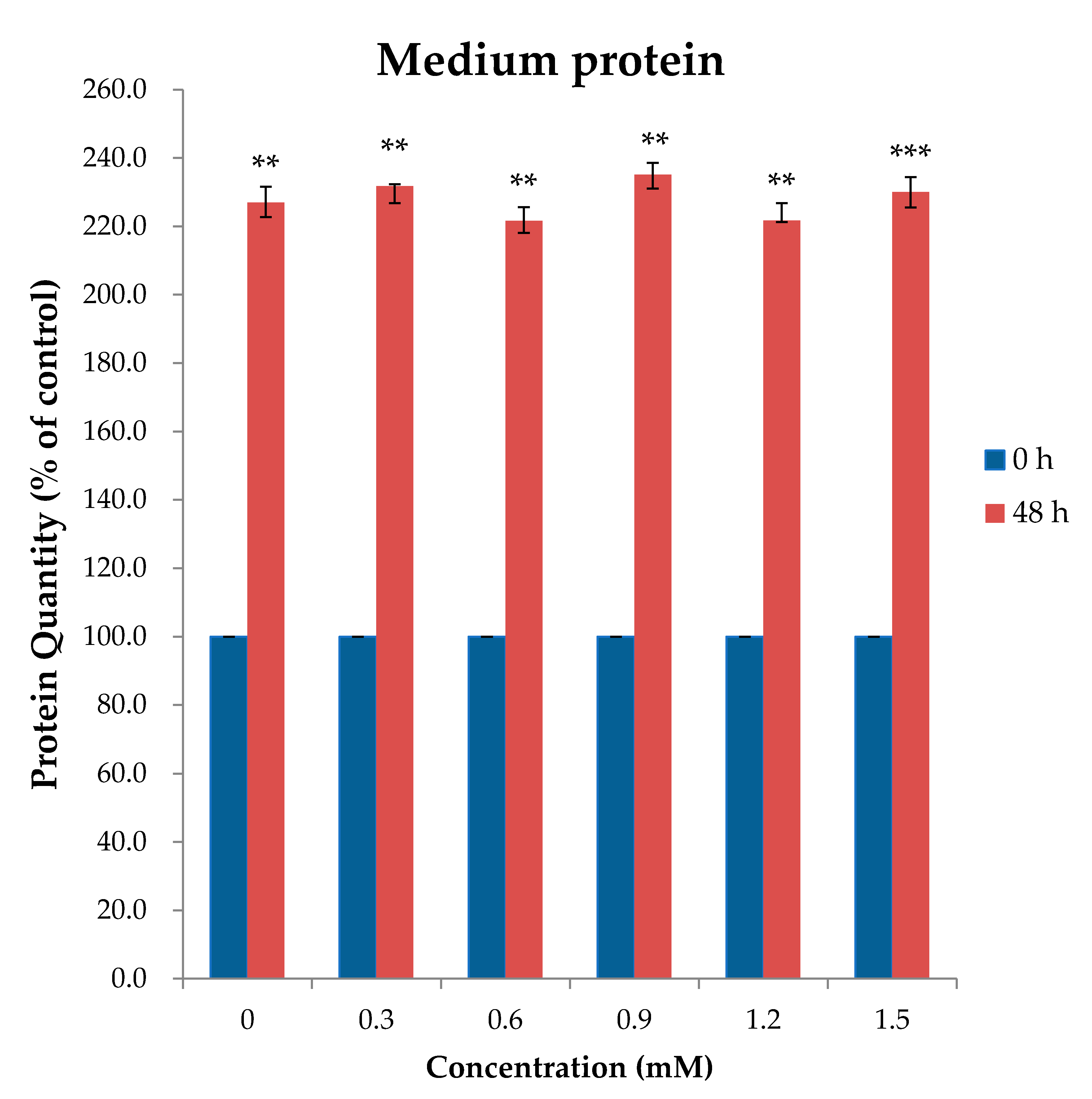
2.1. AA Time and Dosage Sampling
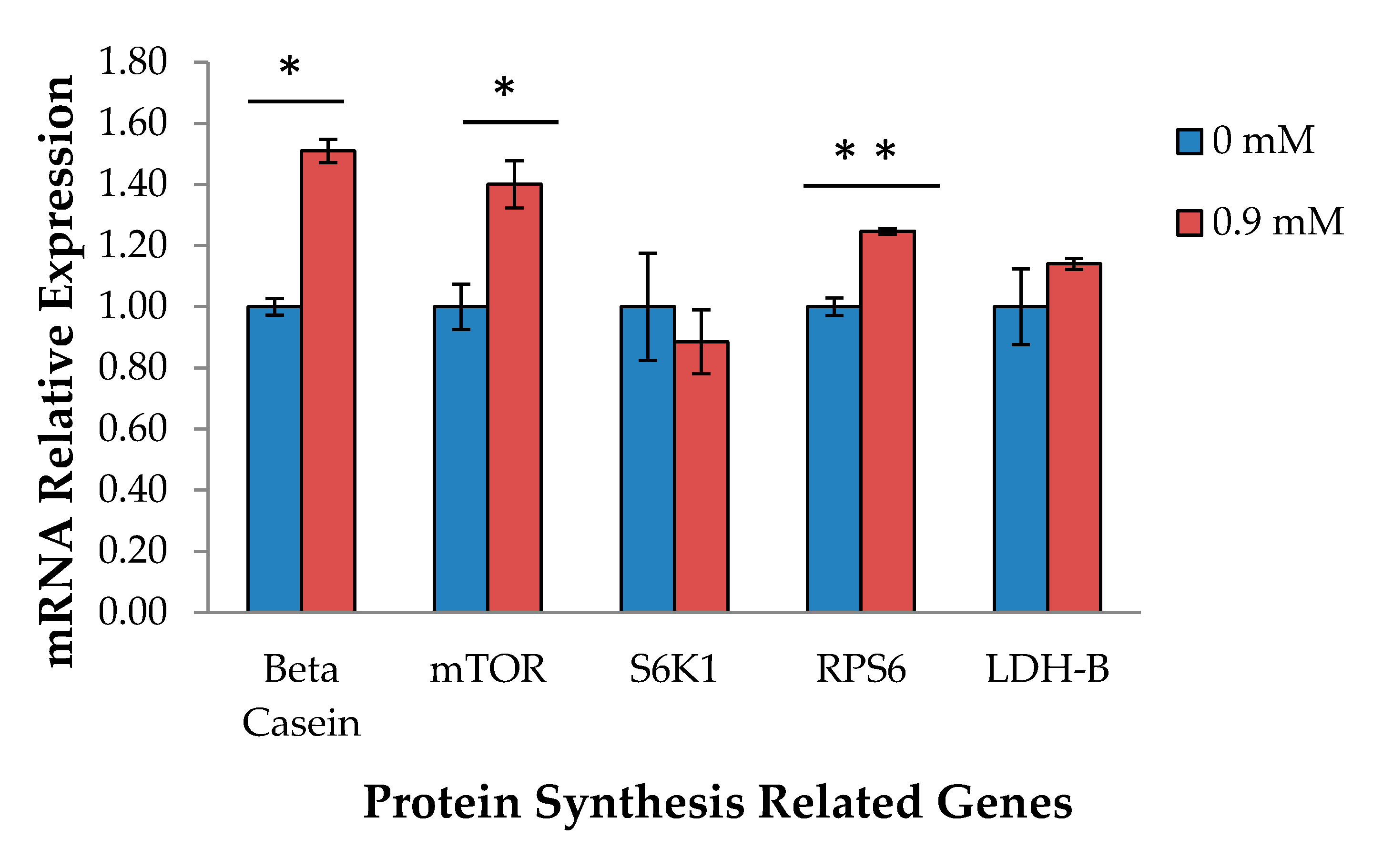
2.2. Real-Time Polymerase Chain Reaction (RT-PCR)
2.3. Proteome Analysis
3. Discussion
3.1. AA Time and Dosage Sampling
3.2. CSN2 and Protein Synthesis-Related Gene Expression
3.3. Proteomics Analysis
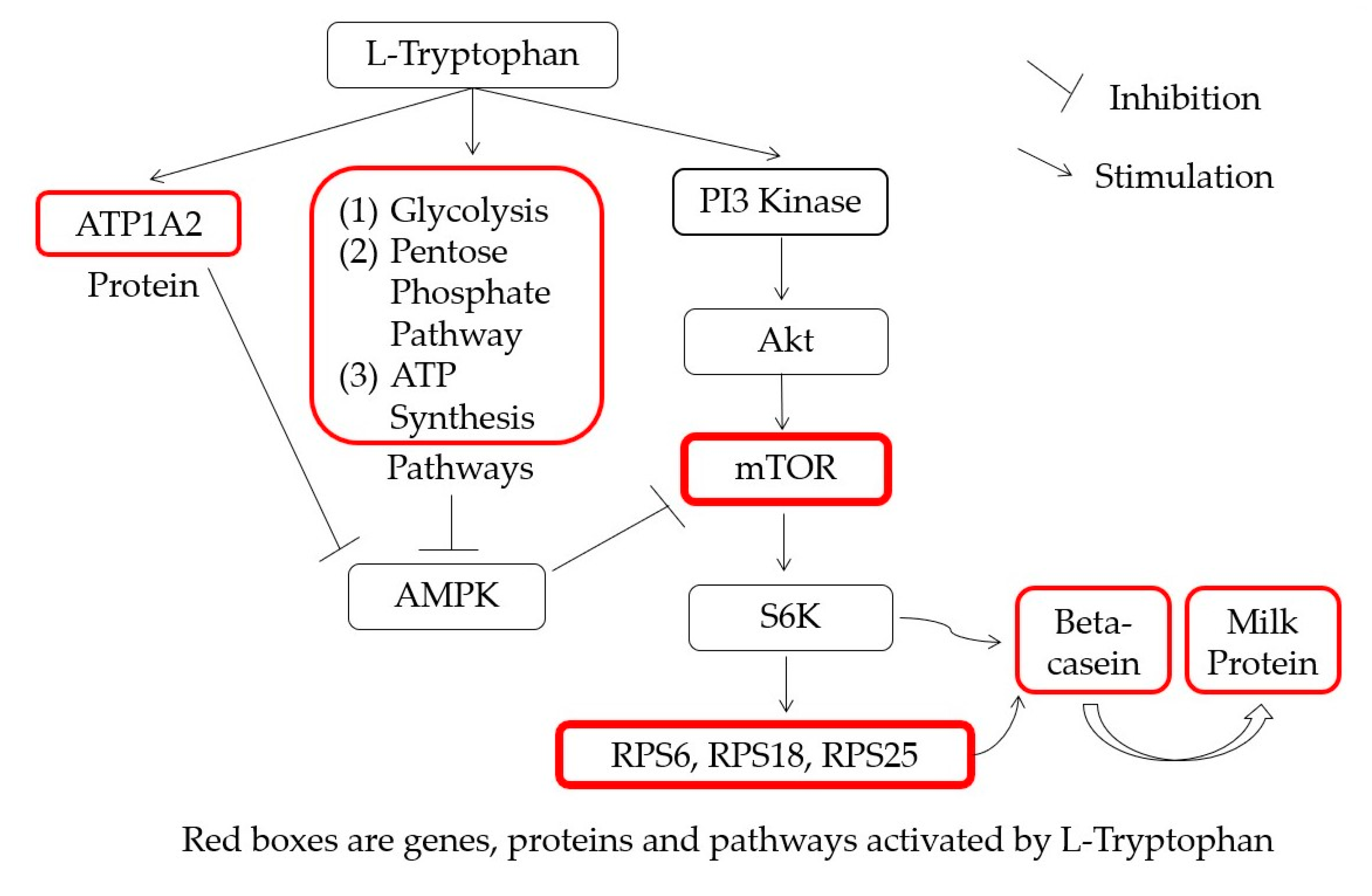
3.4. Metabolic Pathway Analysis
4. Materials and Methods
4.1. AA Dose and Sampling Time
4.2. RNA Extraction and cDNA Synthesis
4.3. Real-Time Polymerase Chain Reaction (RT-PCR)
4.4. Protein Extraction and Quantification
4.5. Proteome Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CSN2 | Beta-casein (β-casein) |
| ATP1A2 | ATP1A2 Sodium/potassium-transporting ATPase subunit alpha-2 |
| AMPK | AMP-activated protein kinase |
| PI3 Kinase | Phosphoinositide 3-kinase |
| Akt | Protein kinase B |
| mTOR | mammalian target of rapamycin |
| S6K | S6 Kinase |
| RPS6 | Ribosomal protein S6 |
| RPS18 | 40S ribosomal protein S18 |
| RPS25 | 40S ribosomal protein S25 |
References
- Haque, M.N.; Guinard-Flament, J.; Lamberton, P.; Mustière, C.; Lemosquet, S. Changes in Mammary Metabolism in Response to the Provision of an Ideal Amino Acid Profile at 2 Levels of Metabolizable Protein Supply in Dairy Cows: Consequences on Efficiency. J. Dairy Sci. 2015, 98, 3951–3968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.S.; Loor, J.J.; Liu, H.Y.; Liu, L.; Hosseini, A.; Zhao, W.S.; Liu, J.X. Optimal Ratios of Essential Amino Acids Stimulate β-Casein Synthesis via Activation of the Mammalian Target of Rapamycin Signaling Pathway in MAC-T Cells and Bovine Mammary Tissue Explants. J. Dairy Sci. 2017, 100, 6676–6688. [Google Scholar] [CrossRef]
- Walczak, K.; Langner, E.; Makuch-Kocka, A.; Szelest, M.; Szalast, K.; Marciniak, S.; Plech, T. Effect of Tryptophan-Derived AhR Ligands, Kynurenine, Kynurenic Acid and FICZ, on Proliferation, Cell Cycle Regulation and Cell Death of Melanoma Cells—In Vitro Studies. Int. J. Mol. Sci. 2020, 21, 7946. [Google Scholar] [CrossRef]
- Lee, S.-B.; Lee, K.-W.; Wang, T.; Lee, J.-S.; Jung, U.-S.; Nejad, J.G.; Oh, Y.-K.; Baek, Y.-C.; Kim, K.H.; Lee, H.-G. Intravenous Administration of L-Tryptophan Stimulates Gastrointestinal Hormones and Melatonin Secretions: Study on Beef Cattle. J. Anim. Sci. Technol. 2019, 61, 239–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-B.; Lee, K.-W.; Wang, T.; Lee, J.-S.; Jung, U.-S.; Nejad, J.G.; Oh, Y.-K.; Baek, Y.-C.; Kim, K.H.; Lee, H.-G. Administration of Encapsulated L-Tryptophan Improves Duodenal Starch Digestion and Increases Gastrointestinal Hormones Secretions in Beef Cattle. Asian Australas. J. Anim. Sci. 2020, 33, 91–99. [Google Scholar] [CrossRef]
- Priatno, W.; Jo, Y.-H.; Nejad, J.G.; Lee, J.-S.; Moon, J.-O.; Lee, H.-G. “Dietary Supplementation of L-Tryptophan” Increases Muscle Development, Adipose Tissue Catabolism and Fatty Acid Transportation in the Muscles of Hanwoo Steers. J. Anim. Sci. Technol. 2020, 62, 595–604. [Google Scholar] [CrossRef]
- Barik, S. The Uniqueness of Tryptophan in Biology: Properties, Metabolism, Interactions and Localization in Proteins. Int. J. Mol. Sci. 2020, 21, 8776. [Google Scholar] [CrossRef] [PubMed]
- Mazinani, M.; Naserian, A.; Rude, B.; Tahmasbi, A.; Valizadeh, R. Effects of Feeding Rumen-Protected Amino Acids on the Performance of Feedlot Calves. J. Adv. Vet. Anim. Res. 2020, 7, 229. [Google Scholar] [CrossRef]
- Nayak, B.N.; Buttar, H.S. Evaluation of the Antioxidant Properties of Tryptophan and Its Metabolites in in Vitro Assay. J. Complement. Integr. Med. 2016, 13, 129–136. [Google Scholar] [CrossRef]
- Friedman, M. Analysis, Nutrition, and Health Benefits of Tryptophan. Int. J. Tryptophan Res. 2018, 11, 117864691880228. [Google Scholar] [CrossRef] [Green Version]
- Gostner, J.M.; Geisler, S.; Stonig, M.; Mair, L.; Sperner-Unterweger, B.; Fuchs, D. Tryptophan Metabolism and Related Pathways in Psychoneuroimmunology: The Impact of Nutrition and Lifestyle. Neuropsychobiology 2020, 79, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.; Geiger, D.; Schauer, M.; Gatterer, H.; Burtscher, M.; Fuchs, D. Effects of Exhaustive Aerobic Exercise on Tryptophan-Kynurenine Metabolism in Trained Athletes. PLoS ONE 2016, 11, e0153617. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, S.; Nakamura, N.; Nakamura, H.; Mizutani, T.; Kaneda, Y.; Yamaguchi, K.; Matsumoto, T.; Kitagawa, J.; Kanemura, N.; Shiraki, M.; et al. Low Levels of Serum Tryptophan Underlie Skeletal Muscle Atrophy. Nutrients 2020, 12, 978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Areta, J.L.; Burke, L.M.; Camera, D.M.; West, D.W.D.; Crawshay, S.; Moore, D.R.; Stellingwerff, T.; Phillips, S.M.; Hawley, J.A.; Coffey, V.G. Reduced Resting Skeletal Muscle Protein Synthesis Is Rescued by Resistance Exercise and Protein Ingestion Following Short-Term Energy Deficit. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E989–E997. [Google Scholar] [CrossRef] [PubMed]
- Török, N.; Tanaka, M.; Vécsei, L. Searching for Peripheral Biomarkers in Neurodegenerative Diseases: The Tryptophan-Kynurenine Metabolic Pathway. Int. J. Mol. Sci. 2020, 21, 9338. [Google Scholar] [CrossRef] [PubMed]
- Toerien, C.A.; Trout, D.R.; Cant, J.P. Nutritional Stimulation of Milk Protein Yield of Cows Is Associated with Changes in Phosphorylation of Mammary Eukaryotic Initiation Factor 2 and Ribosomal S6 Kinase 1. J. Nutr. 2010, 140, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Kimball, S.R. Regulation of Global and Specific MRNA Translation by Amino Acids. J. Nutr. 2002, 132, 883–886. [Google Scholar] [CrossRef] [Green Version]
- Meijer, A.J. Amino Acids as Regulators and Components of Nonproteinogenic Pathways. J. Nutr. 2003, 133, 2057S–2062S. [Google Scholar] [CrossRef]
- Baumrucker, C.R. Amino Acid Transport Systems in Bovine Mammary Tissue. J. Dairy Sci. 1985, 68, 2436–2451. [Google Scholar] [CrossRef]
- Reynolds, C.K.; Harmon, D.L.; Cecava, M.J. Absorption and Delivery of Nutrients for Milk Protein Synthesis by Portal-Drained Viscera. J. Dairy Sci. 1994, 77, 2787–2808. [Google Scholar] [CrossRef]
- Shennan, D.B.; Peaker, M. Transport of Milk Constituents by the Mammary Gland. Physiol. Rev. 2000, 80, 925–951. [Google Scholar] [CrossRef] [PubMed]
- Burgos, S.A.; Dai, M.; Cant, J.P. Nutrient Availability and Lactogenic Hormones Regulate Mammary Protein Synthesis through the Mammalian Target of Rapamycin Signaling Pathway. J. Dairy Sci. 2010, 93, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arriola Apelo, S.I.; Singer, L.M.; Lin, X.Y.; McGilliard, M.L.; St-Pierre, N.R.; Hanigan, M.D. Isoleucine, Leucine, Methionine, and Threonine Effects on Mammalian Target of Rapamycin Signaling in Mammary Tissue. J. Dairy Sci. 2014, 97, 1047–1056. [Google Scholar] [CrossRef]
- Peterson, R.T.; Desai, B.N.; Hardwick, J.S.; Schreiber, S.L. Protein Phosphatase 2A Interacts with the 70-KDa S6 Kinase and Is Activated by Inhibition of FKBP12-Rapamycinassociated Protein. Proc. Natl. Acad. Sci. USA 1999, 96, 4438–4442. [Google Scholar] [CrossRef] [Green Version]
- Navé, B.T.; Ouwens, M.; Withers, D.J.; Alessi, D.R.; Shepherd, P.R. Mammalian Target of Rapamycin Is a Direct Target for Protein Kinase B: Identification of a Convergence Point for Opposing Effects of Insulin and Amino-Acid Deficiency on Protein Translation. Biochem. J. 1999, 344, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Moshel, Y.; Rhoads, R.E.; Barash, I. Role of Amino Acids in Translational Mechanisms Governing Milk Protein Synthesis in Murine and Ruminant Mammary Epithelial Cells. J. Cell. Biochem. 2006, 98, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, A.A.; Nones, K.; Roy, N.C.; McNabb, W.C.; Mackenzie, D.S.; Pacheco, D.; McCoard, S. Initiation and Elongation Steps of MRNA Translation Are Involved in the Increase in Milk Protein Yield Caused by Growth Hormone Administration during Lactation. J. Dairy Sci. 2009, 92, 1889–1899. [Google Scholar] [CrossRef]
- Kim, E. Mechanisms of Amino Acid Sensing in MTOR Signaling Pathway. Nutr. Res. Pract. 2009, 3, 64. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Inoki, K.; Kim, E.; Guan, K.-L. TSC1/TSC2 and Rheb Have Different Effects on TORC1 and TORC2 Activity. Proc. Natl. Acad. Sci. USA 2006, 103, 6811–6816. [Google Scholar] [CrossRef] [Green Version]
- Goberdhan, D.C.I.; Ogmundsdóttir, M.H.; Kazi, S.; Reynolds, B.; Visvalingam, S.M.; Wilson, C.; Boyd, C.A.R. Amino Acid Sensing and MTOR Regulation: Inside or Out? Biochem. Soc. Trans. 2009, 37, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Mahoney, S.J.; Dempsey, J.M.; Blenis, J. Cell Signaling in Protein Synthesis Ribosome Biogenesis and Translation Initiation and Elongation. Prog. Mol. Biol. Transl. Sci. 2009, 90, 53–107. [Google Scholar] [CrossRef]
- Dunlop, E.A.; Tee, A.R. Mammalian Target of Rapamycin Complex 1: Signalling Inputs, Substrates and Feedback Mechanisms. Cell. Signal. 2009, 21, 827–835. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, P.M.J.; Kimball, S.R.; Suryawan, A.; Bush, J.A.; Nguyen, H.V.; Jefferson, L.S.; Davis, T.A. Regulation of Neonatal Liver Protein Synthesis by Insulin and Amino Acids in Pigs. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E994–E1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murgas Torrazza, R.; Suryawan, A.; Gazzaneo, M.C.; Orellana, R.A.; Frank, J.W.; Nguyen, H.V.; Fiorotto, M.L.; El-Kadi, S.; Davis, T.A. Leucine Supplementation of a Low-Protein Meal Increases Skeletal Muscle and Visceral Tissue Protein Synthesis in Neonatal Pigs by Stimulating MTOR-Dependent Translation Initiation. J. Nutr. 2010, 140, 2145–2152. [Google Scholar] [CrossRef] [PubMed]
- Appuhamy, J.A.D.R.N.; Knoebel, N.A.; Nayananjalie, W.A.D.; Escobar, J.; Hanigan, M.D. Isoleucine and Leucine Independently Regulate MTOR Signaling and Protein Synthesis in MAC-T Cells and Bovine Mammary Tissue Slices. J. Nutr. 2012, 142, 484–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Proud, C.G. The MTOR Pathway in the Control of Protein Synthesis. Physiology 2006, 21, 362–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appuhamy, J.A.D.R.N.; Bell, A.L.; Nayananjalie, W.A.D.; Escobar, J.; Hanigan, M.D. Essential Amino Acids Regulate Both Initiation and Elongation of MRNA Translation Independent of Insulin in MAC-T Cells and Bovine Mammary Tissue Slices. J. Nutr. 2011, 141, 1209–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prizant, R.L.; Barash, I. Negative Effects of the Amino Acids Lys, His, and Thr on S6K1 Phosphorylation in Mammary Epithelial Cells. J. Cell. Biochem. 2008, 105, 1038–1047. [Google Scholar] [CrossRef]
- Wang, X. Regulation of Elongation Factor 2 Kinase by P90RSK1 and P70 S6 Kinase. EMBO J. 2001, 20, 4370–4379. [Google Scholar] [CrossRef]
- Christophersen, C.T.; Karlsen, J.; Nielsen, M.O.; Riis, B. Eukaryotic Elongation Factor-2 (EEF-2) Activity in Bovine Mammary Tissue in Relation to Milk Protein Synthesis. J. Dairy Res. 2002, 69, 205–212. [Google Scholar] [CrossRef]
- Luiken, J.J.F.P.; Blommaart, E.F.C.; Boon, L.; van Woerkom, G.M.; Meijer, A.J. Cell Swelling and the Control of Autophagic Proteolysis in Hepatocytes: Involvement of Phosphorylation of Ribosomal Protein S6? Biochem. Soc. Trans. 1994, 22, 508–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blommaart, E.F.; Luiken, J.J.; Blommaart, P.J.; van Woerkom, G.M.; Meijer, A.J. Phosphorylation of Ribosomal Protein S6 Is Inhibitory for Autophagy in Isolated Rat Hepatocytes. J. Biol. Chem. 1995, 270, 2320–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connors, M.T.; Poppi, D.P.; Cant, J.P. Protein Elongation Rates in Tissues of Growing and Adult Sheep. J. Anim. Sci. 2008, 86, 2288–2295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, R.J.; Hellen, C.U.T.; Pestova, T.V. The Mechanism of Eukaryotic Translation Initiation and Principles of Its Regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 113–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orellana, R.A.; Jeyapalan, A.; Escobar, J.; Frank, J.W.; Nguyen, H.V.; Suryawan, A.; Davis, T.A. Amino Acids Augment Muscle Protein Synthesis in Neonatal Pigs during Acute Endotoxemia by Stimulating MTOR-Dependent Translation Initiation. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1416–E1425. [Google Scholar] [CrossRef] [PubMed]
- Lobley, G. Energy Metabolism Reactions in Ruminant Muscle: Responses to Age, Nutrition and Hormonal Status. Reprod. Nutr. Dév. 1990, 30, 13–34. [Google Scholar] [CrossRef] [Green Version]
- Hanigan, M.D.; Baldwin, R.L. A Mechanistic Model of Mammary Gland Metabolism in the Lactating Cow. Agric. Syst. 1994, 45, 369–419. [Google Scholar] [CrossRef]
- Rius, A.G.; Appuhamy, J.A.D.R.N.; Cyriac, J.; Kirovski, D.; Becvar, O.; Escobar, J.; McGilliard, M.L.; Bequette, B.J.; Akers, R.M.; Hanigan, M.D. Regulation of Protein Synthesis in Mammary Glands of Lactating Dairy Cows by Starch and Amino Acids. J. Dairy Sci. 2010, 93, 3114–3127. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Lim, J.-N.; Bok, J.-D.; Kim, J.-H.; Kang, S.-K.; Lee, S.-B.; Hwang, J.-H.; Lee, K.-H.; Kang, H.-S.; Choi, Y.-J.; et al. Association of Protein Expression in Isolated Milk Epithelial Cells and Cis-9, Trans-11 Conjugated Linoleic Acid Proportions in Milk from Dairy Cows: Association of Protein and Cis-9, Trans-11 CLA in Milk. J. Sci. Food Agric. 2014, 94, 1835–1843. [Google Scholar] [CrossRef]
- Wang, T.; Lee, S.B.; Hwang, J.H.; Lim, J.N.; Jung, U.S.; Kim, M.J.; Kang, H.S.; Choi, S.H.; Lee, J.S.; Roh, S.G.; et al. Proteomic Analysis Reveals PGAM1 Altering Cis-9, Trans-11 Conjugated Linoleic Acid Synthesis in Bovine Mammary Gland. Lipids 2015, 50, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially Modified Protein Abundance Index (EmPAI) for Estimation of Absolute Protein Amount in Proteomics by the Number of Sequenced Peptides per Protein. Mol. Cell. Proteom. 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Detected Proteins | ||
|---|---|---|
| Number of protein increased | 51 | |
| Number of protein decreased | 59 | |
| Selected downregulated and upregulated proteins | ▲▼ | |
| HSPD1 (60 kDa heat shock protein, mitochondrial) | HSPD1 | ▲ |
| HSPA1A (Heat shock 70 kDa protein 1A) | HSPA1A | ▲ |
| ATP5B (ATP synthase subunit beta, mitochondrial) | ATP5B | ▼ |
| EEF1A1 (Elongation factor 1 alpha 1) | EEF1A1 | ▼ |
| RPSA (Similar to 40S ribosomal protein SA (fragment)) | RPSA | ▼ |
| ATP synthase subunit alpha, mitochondrial | ATP5A1 | ▼ |
| RPS18 (40S ribosomal protein S18) | RPS18 | ▲ |
| EIF4A1 (Eukaryotic initiation factor 4A-I) | EIF4A1 | ▼ |
| EEF2 (Elongation factor 2) | EEF2 | ▼ |
| RPS25 (40S ribosomal protein S25) | RPS25 | ▲ |
| EEF1G (Elongation factor 1-gamma) | EEF1G | ▼ |
| RPN2 (Dolichyl-diphosphooligosaccharide–protein glycosyltransferase) | RPN2 | ▼ |
| ATP1A2 (Sodium/potassium-transporting ATPase subunit alpha-2) | ATP1A2 | ▼ |
| GPI (Glucose-6-phosphate isomerase) | GPI | ▼ |
| RPL11 (60S ribosomal protein L11) | RPL11 | ▼ |
| RPS2 (40S ribosomal protein S2) | RPS2 | ▼ |
| Detected Pathways * |
|---|
| Apoptosis signaling pathway |
| p53 pathway |
| Glycolysis |
| Pentose phosphate pathway |
| ATP synthesis |
| CCKR signaling map |
| Endothelin signaling pathway |
| FGF signaling pathway |
| Ras pathway |
| EGF receptor signaling pathway |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conejos, J.R.V.; Ghassemi Nejad, J.; Kim, J.-E.; Moon, J.-O.; Lee, J.-S.; Lee, H.-G. Supplementing with L-Tryptophan Increases Medium Protein and Alters Expression of Genes and Proteins Involved in Milk Protein Synthesis and Energy Metabolism in Bovine Mammary Cells. Int. J. Mol. Sci. 2021, 22, 2751. https://doi.org/10.3390/ijms22052751
Conejos JRV, Ghassemi Nejad J, Kim J-E, Moon J-O, Lee J-S, Lee H-G. Supplementing with L-Tryptophan Increases Medium Protein and Alters Expression of Genes and Proteins Involved in Milk Protein Synthesis and Energy Metabolism in Bovine Mammary Cells. International Journal of Molecular Sciences. 2021; 22(5):2751. https://doi.org/10.3390/ijms22052751
Chicago/Turabian StyleConejos, Jay Ronel V., Jalil Ghassemi Nejad, Jung-Eun Kim, Jun-Ok Moon, Jae-Sung Lee, and Hong-Gu Lee. 2021. "Supplementing with L-Tryptophan Increases Medium Protein and Alters Expression of Genes and Proteins Involved in Milk Protein Synthesis and Energy Metabolism in Bovine Mammary Cells" International Journal of Molecular Sciences 22, no. 5: 2751. https://doi.org/10.3390/ijms22052751
APA StyleConejos, J. R. V., Ghassemi Nejad, J., Kim, J. -E., Moon, J. -O., Lee, J. -S., & Lee, H. -G. (2021). Supplementing with L-Tryptophan Increases Medium Protein and Alters Expression of Genes and Proteins Involved in Milk Protein Synthesis and Energy Metabolism in Bovine Mammary Cells. International Journal of Molecular Sciences, 22(5), 2751. https://doi.org/10.3390/ijms22052751