
The Landscape of RNA-Protein Interactions in Plants: Approaches and Current Status


Abstract
:1. Introduction
2. Overview of Methods to Detect RNA-Protein Interactions and Their Application to Plants

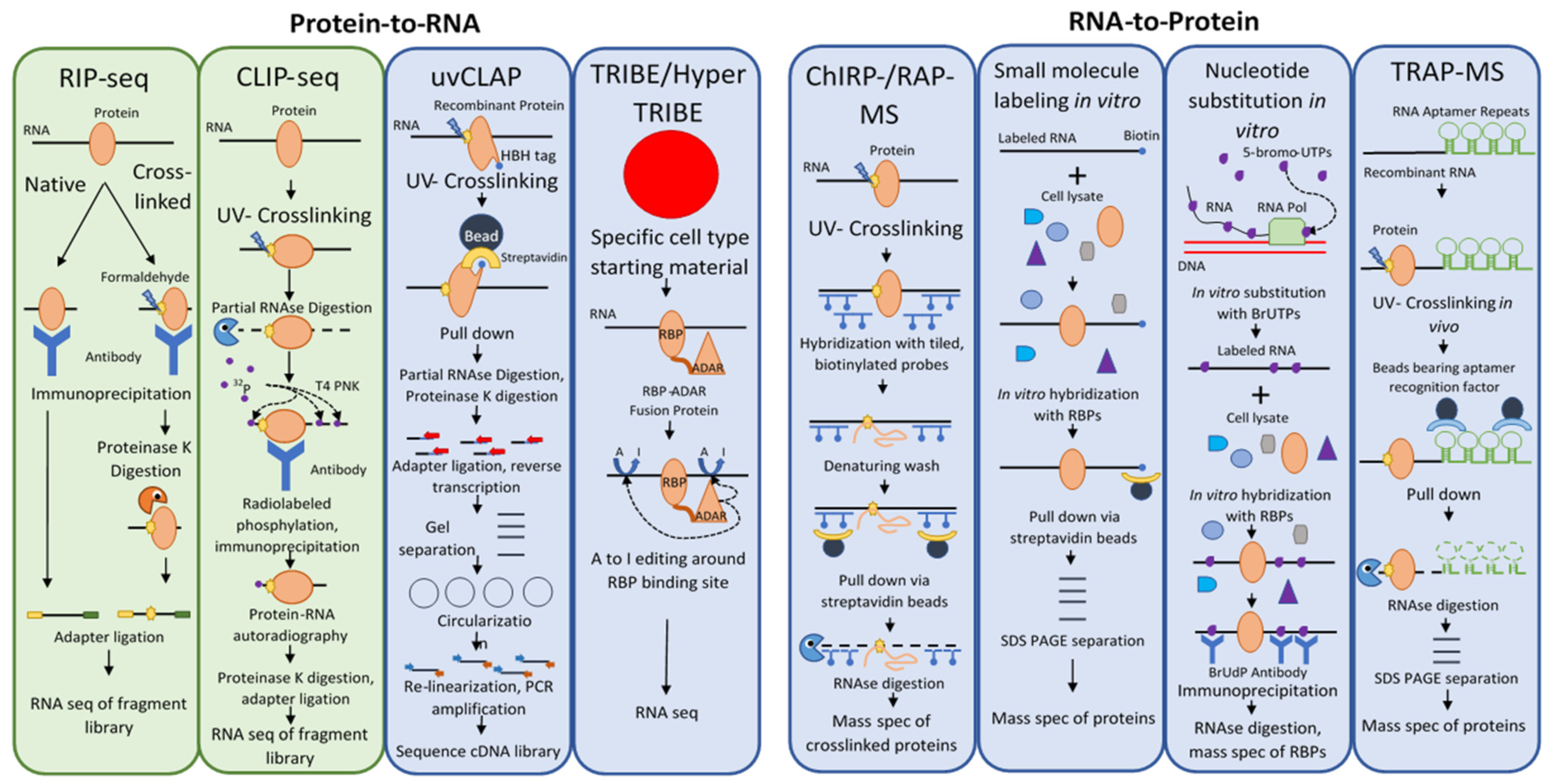
2.1. Methods That Use a Protein Bait to Identify Its RNA Targets (Protein-to-RNA) and RBPs Identified Using These Methods in Plants
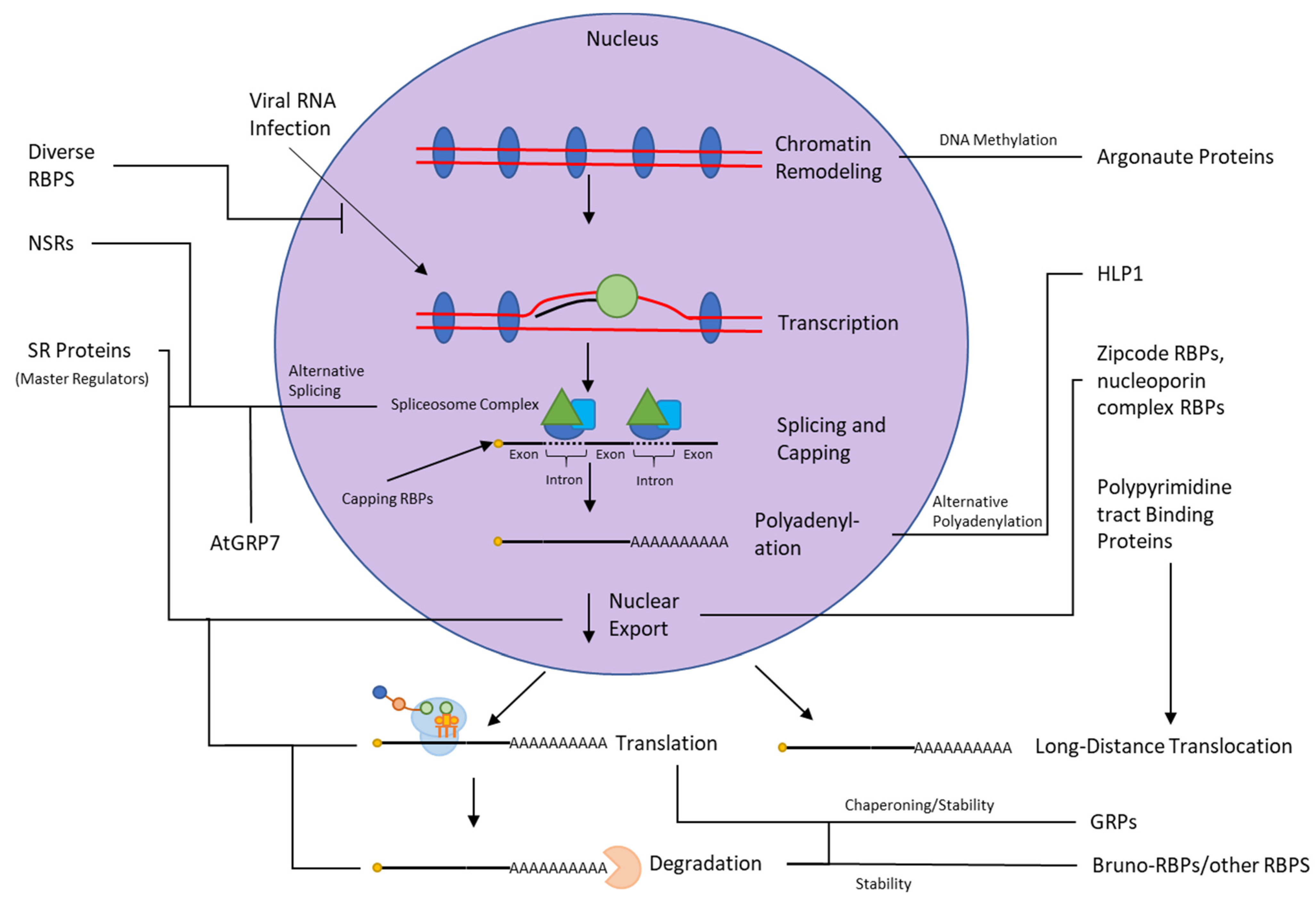
2.1.1. Regulation of RNA Processing
2.1.2. Trafficking and Translocation
2.1.3. Chaperoning
2.1.4. Gene Silencing
2.1.5. Viral RNA Suppression
2.1.6. Other RBPs
2.1.7. Perspective on the Application of Protein-to-RNA Methods in Plants
2.2. Methods That Use an RNA Bait to Identify Binding Proteins (RNA-to-Protein)
2.3. Global RNA-Protein Interactome
3. RNA-Protein Interactions in Plants as Unearthed by Classical Methods
3.1. Regulation of RNA Processing
3.2. Trafficking and Translocation
3.3. Chaperoning
3.4. Stability and Decay
3.5. Gene Silencing
3.6. Viral RNA Suppression
3.7. Other Plant RBPs
4. Manipulation of RBPs Confers Desirable Traits in Plants
5. Conclusions and Perspectives
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marondedze, C. The increasing diversity and complexity of the RNA-binding protein repertoire in plants. Proc. R. Soc. 2020, 287, 20201397. [Google Scholar] [CrossRef] [PubMed]
- Lorkovic, Z. Role of plant RNA-binding proteins in development, stress response and genome organization. Trends Plant Sci. 2009, 14, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Woloshen, V.; Huang, S.; Li, X. RNA-Binding Proteins in Plant Immunity. J. Pathog. 2011, 2011, 278697. [Google Scholar] [CrossRef] [Green Version]
- Huh, S.; Paek, K. Plant RNA binding proteins for control of RNA virus infection. Front. Physiol. 2013, 4, 397. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Kang, H. Emerging Roles of RNA-Binding Proteins in Plant Growth, Development, and Stress Responses. Mol. Cells 2016, 39, 179–185. [Google Scholar]
- Koster, T.; Marondedze, C.; Meyer, K.; Staiger, D. RNA-Binding Proteins Revisited—The Emerging Arabidopsis mRNA Interactome. Trends Plant Sci. 2017, 22, 512–526. [Google Scholar] [CrossRef]
- Dedow, L.; Bailey-Serres, J. Searching for a Match: Structure, Function and Application of Sequence-Specific RNA-Binding Proteins. Plant Cell Physiol. 2019, 60, 1927–1938. [Google Scholar] [CrossRef] [PubMed]
- Marondedze, C.; Thomas, L.; Serrano, N.; Lilley, K.; Gehring, C. The RNA-binding protein repertoire of Arabidopsis thaliana. Sci. Rep. 2016, 6, 29766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castello, A.; Hentze, M.; Preiss, T. Metabolic Enzymes Enjoying New Partnerships as RNA-Binding Proteins. Trends Endocrinol. Metab. 2015, 26, 746–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hentz, M.; Preiss, T. The REM phase of gene regulation. Trends Biochem. Sci. 2010, 35, 423–426. [Google Scholar] [CrossRef]
- Vermel, M.; Guermann, B.; Delage, L.; Grienenberger, J.; Maréchal-Drouard, L.; Gualberto, J. A family of RRM-type RNA-binding proteins specific to plant mitochondria. PNAS 2002, 99, 5866–5871. [Google Scholar] [CrossRef] [Green Version]
- Staiger, D.; Zecca, L.; Wieczorek Kirk, D.; Apel, K.; Eckstein, L. The circadian clock regulated RNA-binding protein AtGRP7 autoregulates its expression by influencing alternative splicing of its own pre-mRNA. Plant J. 2003, 33, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, I. Regulation and function of SOC1, a flowering pathway integrator. J. Exp. Bot. 2010, 61, 2247–2254. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Miles, W. Beyond CLIP: Advances and opportunities to measure RBP–RNA and RNA–RNA interactions. Nucleic Acids Res. 2019, 47, 5490–5501. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Hsu, Y.; Zhu, S.; Wierzbicki, A.; Pontes, O.; Pikaard, C.; Liu, H.; Wang, C.; Jin, H.; Zhu, J. An effector of RNA-directed DNA methylation in Arabidopsis is an ARGONAUTE 4- and RNA-binding protein. Cell 2009, 137, 498–508. [Google Scholar] [CrossRef] [Green Version]
- Gagliardi, M.; Matarazzo, M. RIP: RNA Immunoprecipitation. Methods Mol. Biol. 2016, 1480, 73–86. [Google Scholar] [PubMed]
- Zambelli, F.; Pavesi, G. RIP-Seq Data Analysis to Determine RNA-protein Associations. Methods Mol. Biol. 2015, 1269, 293–303. [Google Scholar] [PubMed]
- Niranjanakumari, S.; Lasda, E.; Brazas, R.; Garcia-Blanco, M. Reversible cross-linking combined with immunoprecipitation to study RNA-protein interactions in vivo. Methods 2002, 26, 182–190. [Google Scholar] [CrossRef]
- Ule, J.; Jensen, K.; Ruggiu, M.; Mele, A.; Ule, A.; Darnell, R. CLIP Identifies Nova-Regulated RNA Networks in the Brain. Science 2003, 302, 1212–1215. [Google Scholar] [CrossRef]
- Licatalosi, D.; Mele, A.; Fak, J.; Ule, J.; Kayikci, M.; Chi, S.; Clark, T.; Schweitzer, A.; Blume, J.; Wang, X.; et al. HITS-CLIP yields genome-wide insights into brain alternative RNA processing. Nature 2008, 456, 464–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafner, M.; Landthaler, M.; Burger, L.; Khorshid, M.; Hausser, J.; Berninger, P.; Rothballer, A.; Ascano, M.; Jungkamp, A.; Munschauer, M.; et al. Transcriptome-wide Identification of RNA-Binding Protein and MicroRNA Target Sites by PAR-CLIP. Cell 2010, 141, 129–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, J.; Zarnack, K.; Rot, G.; Curk, T.; Kayikci, M.; Zupan, B.; Turner, D.; Luscombe, N.; Ule, J. iCLIP reveals the function of hnRNP particles in splicing at individual nucleotide resolution. Nat. Struct. Mol. Biol. 2014, 17, 909–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nostrand, E.; Pratt, G.; Shishkin, A.; Gelboin-Burkhart, C.; Fang, M.; Sundararaman, B.; Blue, S.; Nguyen, T.; Surka, C.; Elkins, K.; et al. Robust transcriptome-wide discovery of RNA-binding protein binding sites with enhanced CLIP (eCLIP). Nat. Methods 2016, 13, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Kargapolova, Y.; Levin, M.; Lackner, K.; Danckwardt, S. sCLIP––An integrated platform to study RNA–protein interactomes in biomedical research: Identification of CSTF2tau in alternative processing of small nuclear RNAs. Nucleic Acids Res. 2017, 45, 6074–6086. [Google Scholar] [CrossRef]
- Zarnegar, B.; Flynn, R.; Shen, Y.; Do, B.; Chang, H.; Khavari, P. irCLIP platform for efficient characterization of protein–RNA interactions. Nat. Methods 2015, 13, 489–492. [Google Scholar] [CrossRef]
- Wang, T.; Xie, Y.; Xiao, G. dCLIP: A computational approach for comparative CLIP-seq analyses. Genome Biol. 2014, 15, R11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramanathan, M.; Porter, D.; Khavari, P. Methods to study RNA–protein interactions. Nat. Methods 2019, 16, 225–234. [Google Scholar] [CrossRef]
- Darnell, R. HITS-CLIP: Panoramic views of protein-RNA regulation in living cells. Wiley Interdiscip. Rev. RNA 2010, 1, 266–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maticzka, D.; Ilik, I.; Aktas, T.; Backofen, R.; Akhtar, A. uvCLAP is a fast and non-radioactive method to identify in vivo targets of RNA-binding proteins. Nat. Commun. 2018, 9, 1142. [Google Scholar] [CrossRef] [Green Version]
- McMahon, A.; Rahman, R.; Jin, H.; Shen, J.; Fieldsend, A.; Luo, W.; Rosbash, M. TRIBE: Hijacking an RNA-Editing Enzyme to Identify Cell-Specific Targets of RNA-Binding Proteins. Cell 2016, 165, 742–753. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Rahman, R.; Rosbash, M. Mechanistic implications of enhanced editing by a HyperTRIBE RNA-binding protein. RNA 2018, 24, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Xing, D.; Wang, Y.; Hamilton, M.; Ben-Hur, A.; Reddy, A. Transcriptome-Wide Identification of RNA Targets of Arabidopsis SERINE/ARGININE-RICH45 Uncovers the Unexpected Roles of This RNA Binding Protein in RNA Processing. Plant Cell 2015, 27, 3294–3308. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, Y.; Powers, J.; Gowda, N.; Zhang, C.; Ibrahim, H.; Ball, H.; Chen, S.; Lu, H.; Mount, S. Transcriptome analyses reveal SR45 to be a neutral splicing regulator and a suppressor of innate immunity in Arabidopsis thaliana. BMC Genom. 2017, 18, 772. [Google Scholar] [CrossRef] [PubMed]
- Albaqami, M.; Laluk, K.; Reddy, A. The Arabidopsis splicing regulator SR45 confers salt tolerance in a splice isoform-dependent manner. Plant Mol. Biol. 2019, 100, 379–390. [Google Scholar] [CrossRef]
- Park, H.; You, Y.; Lee, A.; Jung, H.; Jo, S.; Oh, N.; Kim, H.; Lee, H.; Kim, J.; Kim, Y.; et al. OsFKBP20-1b interacts with the splicing factor OsSR45 and participates in the environmental stress response at the post-transcriptional level in rice. Plant J. 2020, 102, 992–1007. [Google Scholar] [CrossRef] [PubMed]
- Francisco-Mangilet, A.; Karlsson, P.; Kim, M.; Eo, H.; Oh, S.; Kim, J.; Kulcheski, F.; Park, S.; Manavella, P. THO2, a core member of the THO/TREX complex, is required for microRNA production in Arabidopsis. Plant J. 2015, 82, 1018–1029. [Google Scholar] [CrossRef]
- Streitner, C.; Danisman, S.; Wehrle, F.; Schoning, J.; Alfano, J.; Staiger, D. The small glycine-rich RNA binding protein AtGRP7 promotes floral transition in Arabidopsis thaliana. Plant J. 2008, 56, 239–250. [Google Scholar] [CrossRef]
- Meyer, K.; Koster, T.; Nolte, C.; Weinholdt, C.; Lewinski, M.; Grosse, I.; Staiger, D. Adaptation of iCLIP to plants determines the binding landscape of the clockregulated RNA-binding protein AtGRP7. Genome Biol. 2017, 18, 204. [Google Scholar] [CrossRef] [Green Version]
- Bardou, F.; Ariel, F.; Simpson, C.; Romero-Barrios, N.; Laporte, P.; Balzergue, S.; Brown, J.; Crespi, M. Long Noncoding RNA Modulates Alternative Splicing Regulators in Arabidopsis. Dev. Cell 2014, 30, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Bazin, J.; Romero, N.; Rigo, R.; Charon, C.; Blein, T.; Ariel, F.; Crespi, M. Nuclear Speckle RNA Binding Proteins Remodel Alternative Splicing and the Non-coding Arabidopsis Transcriptome to Regulate a Cross-Talk Between Auxin and Immune Responses. Front. Plant Sci. 2018, 9, 1209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gu, L.; Hou, Y.; Wang, L.; Deng, X.; Hang, R.; Chen, D.; Zhang, X.; Zhang, Y.; Liu, C.; et al. Integrative genome-wide analysis reveals HLP1, a novel RNA-binding protein, regulates plant flowering by targeting alternative polyadenylation. Cell Res. 2015, 25, 864–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Q.; Cui, Y.; Zhang, G.; Zhang, H.; Wang, X.; Yang, Z. The Arabidopsis pentatricopeptide repeat protein PDM1 is associated with the intergenic sequence of S11-rpoA for rpoA monocistronic RNA cleavage. Chin. Sci. Bull. 2012, 57, 3452–3459. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Dorozhenk, K.; Zhang, L.; Fukuda, M.; Washida, H.; Kumamaru, T.; Okita, T. Zipcode RNA-Binding Proteins and Membrane Trafficking Proteins Cooperate to Transport Glutelin mRNAs in Rice Endosperm. Plant Cell 2020, 32, 2566–2581. [Google Scholar] [CrossRef]
- Tian, L.; Chou, H.; Zhang, L.; Okita, T. Targeted Endoplasmic Reticulum Localization of Storage Protein mRNAs Requires the RNA-Binding Protein RBP-L. Plant Physiol. 2019, 179, 1111–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juntawong, P.; Sorenson, R.; Bailey-Serres, J. Cold shock protein 1 chaperones mRNAs during translation in Arabidopsis thaliana. Plant J. 2013, 74, 1016–1028. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicki, A.; Ream, T.; Haag, J.; Pikaard, C. RNA Polymerase V transcription guides ARGONAUTE4 to chromatin. Nat. Genet. 2009, 41, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Rowley, J.; Bohmdorfer, G.; Wierzbicki, A. A SWI/SNF Chromatin-Remodeling Complex Acts in Noncoding RNA-Mediated Transcriptional Silencing. Mol. Cell 2013, 49, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Rowley, J.; Bohmdorfer, G.; Wierzbicki, A. Analysis of long non-coding RNAs produced by a specialized RNA Polymerase in Arabidopsis thaliana. Methods 2013, 63, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Marmisolle, F.; García, M.; Reyes, C. RNA-binding protein immunoprecipitation as a tool to investigate plant miRNA processing interference by regulatory proteins of diverse origin. Plant Methods 2018, 14, 9. [Google Scholar] [CrossRef] [Green Version]
- Schmid, L.; Ohler, L.; Mohlmann, T.; Brachmann, A.; Muino, J.; Leister, D.; Meurer, J.; Manavski, N. PUMPKIN, the Sole Plastid UMP Kinase, Associates with Group II Introns and Alters Their Metabolism. Plant Physiol. 2019, 179, 248–264. [Google Scholar] [CrossRef] [Green Version]
- Libault, M.; Pingault, L.; Zogli, P.; Schiefelbein, J. Plant Systems Biology at the Single-Cell Level. Trends Plant Sci. 2017, 22, 949–960. [Google Scholar] [CrossRef]
- Martin, L.; Nicolas, P.; Matas, A.; Shinozaki, Y.; Catalá, C.; Rose, J. Laser microdissection of tomato fruit cell and tissue types for transcriptome profiling. Nat. Protoc. 2016, 11, 2376–2388. [Google Scholar] [CrossRef] [PubMed]
- Engreitz, J.; Pandya-Jones, A.; McDonel, P.; Shishkin, A.; Sirokman, K.; Surka, C.; Kadri, S.; Xing, J.; Goren, A.; Lander, E.; et al. The Xist lncRNA Exploits Three-Dimensional Genome Architecture to Spread across the X Chromosome. Science 2013, 341, 1237973. [Google Scholar] [CrossRef] [Green Version]
- McHugh, C.; Chen, C.; Chow, A.; Surka, C.; Tran, C.; McDonel, P.; Pandya-Jones, A.; Blanco, M.; Burghard, C.; Moradian, A.; et al. The Xist lncRNA interacts directly with SHARP to silence transcription through HDAC3. Nature 2015, 521, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Qu, K.; Zhong, F.; Artandi, S.; Chang, H. Genomic Maps of Long Noncoding RNA Occupancy Reveal Principles of RNA-Chromatin Interactions. Mol. Cell 2011, 44, 667–678. [Google Scholar] [CrossRef] [Green Version]
- Chu, C.; Zhang, Q.; Heard, E.; Chang, H. Systematic Discovery of Xist RNA Binding Proteins. Cell 2015, 161, 404–416. [Google Scholar] [CrossRef] [Green Version]
- Foley, S.; Gosai, S.; Wang, D.; Selamoglu, N.; Sollitti, A.; Koster, T.; Steffen, A.; Lyons, E.; Daldal, F.; Garcia, B.; et al. A Global View of RNA-Protein Interactions Identifies Post-transcriptional Regulators of Root Hair Cell Fate. Dev. Cell 2017, 41, 204–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Sun, W.; Shi, T.; Lu, P.; Zhuang, M.; Liu, J. Capturing RNA–protein interaction via CRUIS. Nucleic Acids Res. 2020, 48, e52. [Google Scholar] [CrossRef]
- Gemmill, D.; D’souza, S.; Meier-Stephenson, V.; Patel, T. Current approaches for RNA-labelling to identify RNA-binding proteins. Biochem. Cell. Biol. 2020, 98, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Johansson, H.; Liljas, L.; Uhlenbeck, O. RNA Recognition by the MS2 Phage Coat Protein. Semin. Virol. 1997, 8, 176–185. [Google Scholar] [CrossRef]
- Lim, F.; Downey, T.; Peabody, D. Translational Repression and Specific RNA Binding by the Coat Protein of the Pseudomonas Phage PP7. J. Biol. Chem. 2001, 276, 22507–22513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Tan, W.; Zu, Y. Aptamers: Versatile molecular recognition probes for cancer detection. Analyst 2016, 141, 403. [Google Scholar] [CrossRef] [Green Version]
- Mallikaratchy, P. Evolution of Complex Target SELEX to Identify Aptamers against Mammalian Cell-Surface Antigens. Molecules 2017, 22, 215. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Zhao, B.S.; Myers, S.; Carr, S.; He, C.; Ting, A. RNA–protein interaction mapping via MS2- or Cas13-based APEX targeting. PNAS 2020, 117, 22068–22079. [Google Scholar] [CrossRef]
- Mukherjee, J.; Hermesh, O.; Eliscovich, C.; Nalpas, N.; Franz-Wachtel, M.; Macek, B.; Jansen, R.P. β-Actin mRNA interactome mapping by proximity biotinylation. PNAS 2019, 116, 12863–12872. [Google Scholar] [CrossRef] [Green Version]
- Kaewsapsak, P.; Schechner, D.; Mallard, W.; Rinn, J.; Ting, A. Live-cell mapping of organelle-associated RNAs via proximity biotinylation combined with protein-RNA crosslinking. eLife 2017, 6, e29224. [Google Scholar] [CrossRef]
- Yi, W.; Li, J.; Zhu, X.; Wang, X.; Fan, L.; Sun, W.; Liao, L.; Zhang, J.; Ye, J.; Chen, F.; et al. CRISPR-assisted detection of RNA–protein interactions in living cells. Nat. Methods 2020, 17, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Slavkovic, S.; Churcher, Z.; Johnson, P. Nanomolar binding affinity of quinine-based antimalarial compounds by the cocaine-binding aptamer. Bioorg. Med. Chem. 2018, 1, 5427–5434. [Google Scholar] [CrossRef] [PubMed]
- Debiais, M.; Lelievre, A.; Smietana, M.; Muller, S. Splitting aptamers and nucleic acid enzymes for the development of advanced biosensors. Nucleic Acids Res. 2020, 48, 3400–3422. [Google Scholar] [CrossRef]
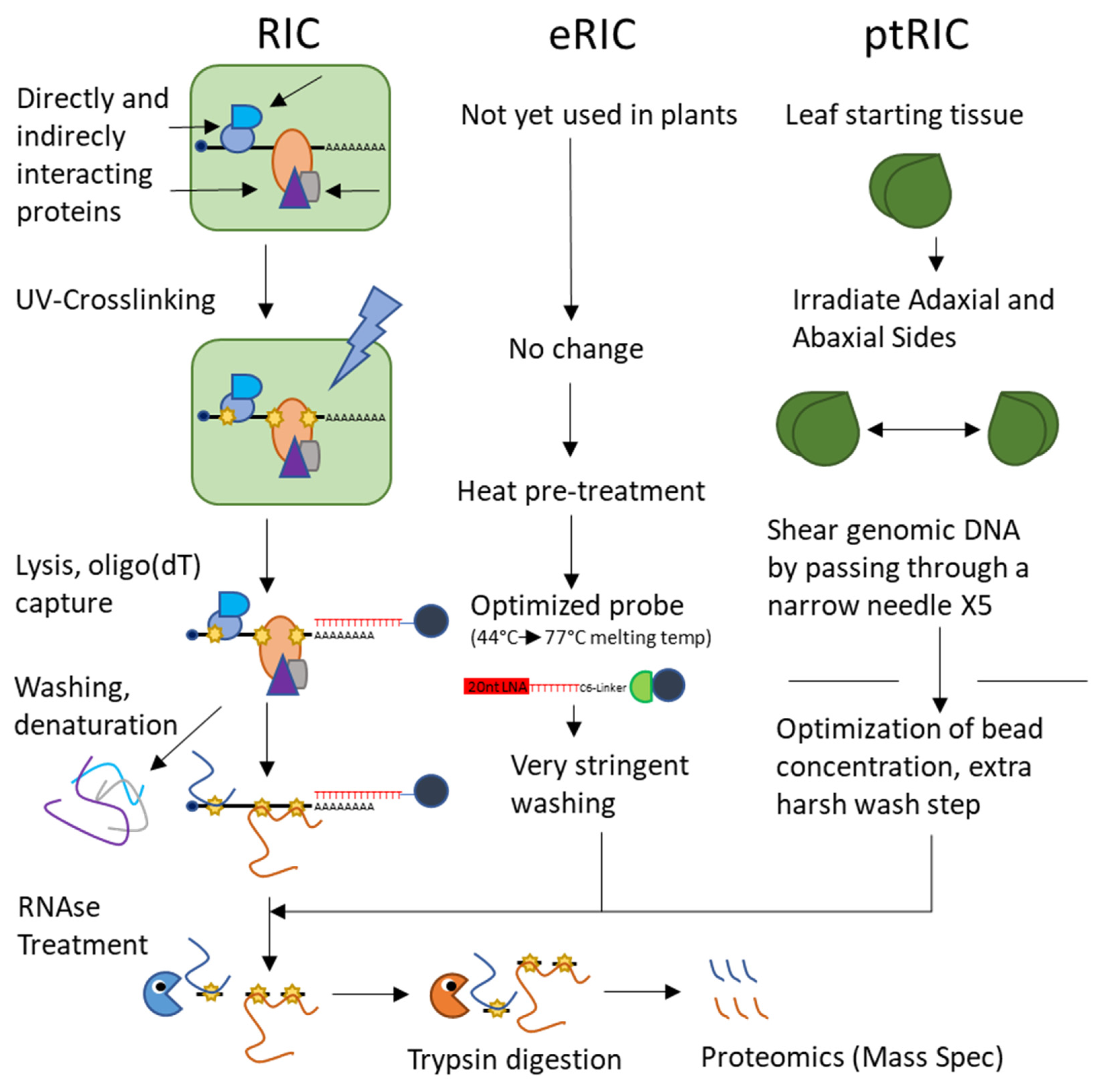
- Bach-Pages, M.; Castello, A.; Preston, G. Plant RNA Interactome Capture: Revealing the Plant RBPome. Trends Plant Sci. 2017, 22, 449–451. [Google Scholar] [CrossRef]
- Reichel, M.; Liao, Y.; Rettel, M.; Ragan, C.; Evers, M.; Alleaume, A.; Horos, R.; Hentze, M.; Preiss, T.; Millar, A. In Planta Determination of the mRNA-Binding Proteome of Arabidopsis Etiolated Seedlings. Plant Cell 2016, 28, 2435–2452. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Boonen, K.; Ferrari, P.; Schoofs, L.; Janssens, E.; van Noort, V.; Rolland, F.; Geuten, K. UV crosslinked mRNA-binding proteins captured from leaf mesophyll protoplasts. Plant Methods 2016, 12, 42. [Google Scholar] [CrossRef] [Green Version]
- Marondedze, C.; Thomas, L.; Gehring, C.; Lilley, K. Changes in the Arabidopsis RNA-binding proteome reveal novel stress response mechanisms. BMC Plant Biol. 2019, 19, 139. [Google Scholar] [CrossRef]
- Marondedze, C.; Thomas, L.; Lilley, K.; Gehring, C. Drought Stress Causes Specific Changes to the Spliceosome and Stress Granule Components. Front. Mol. Biosci. 2020, 6, 163. [Google Scholar] [CrossRef] [Green Version]
- Perez-Perri, J.; Rogell, B.; Schwarzi, T.; Stein, F.; Zhou, Y.; Rettel, M.; Brosig, A.; Hentze, M. Discovery of RNA-binding proteins and characterization of their dynamic responses by enhanced RNA interactome capture. Nat. Commun. 2018, 9, 4408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bach-Pages, M.; Homma, F.; Kourelis, J.; Kaschani, F.; Mohammed, S.; Kaiser, M.; van der Hoorn, R.; Castello, A.; Preston, G. Discovering the RNA-Binding Proteome of Plant Leaves with an Improved RNA Interactome Capture Method. Biomolecules 2020, 10, 661. [Google Scholar] [CrossRef]
- Queiroz, R.; Smith, T.; Villanueva, E.; Marti-Solano, M.; Monit, M.; Pizzinga, M.; Mirea, D.; Ramakrishna, M.; Harvey, R.; Dezi, V.; et al. Comprehensive identification of RNA–protein interactions in any organism using orthogonal organic phase separation (OOPS). Nat. Biotechnol. 2019, 37, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, C.; Jia, X.; Wang, W.; Yin, H. Comparative analysis of RNA-binding proteomes under Arabidopsis thaliana-Pst DC3000-PAMP interaction by orthogonal organic phase separation. Int. J. Biol. Macromol. 2020, 160, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Hocine, S.; Singer, R.; Grunwald, D. RNA Processing and Export. Cold Spring Harb. Perspect. Biol. 2010, 2, a000752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, K.; Patil, D.; Zhou, J.; Zinoviev, A.; Skabkin, M.; Elemento, O.; Pestova, T.; Qian, S.; Jaffrey, S. 5′ UTR m6A Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, A.S.N.; Marquez, Y.; Kalyna, M.; Barta, A. Complexity of the alternative splicing landscape in plants. Plant Cell 2013, 25, 3657–3683. [Google Scholar] [CrossRef] [Green Version]
- Shang, X.; Cao, Y.; Ma, L. Alternative Splicing in Plant Genes: A Means of Regulating the Environmental Fitness of Plants. Int. J. Mol. Sci. 2017, 18, 432. [Google Scholar] [CrossRef]
- Hunt, A. RNA regulatory elements and polyadenylation in plants. Front. Plant Sci. 2012, 2, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, A.; Xing, D.; Li, Q. Plant polyadenylation factors: Conservation and variety in the polyadenylation complex in plants. BMC Genom. 2012, 13, 641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saéz-Vásquez, J.; Delseny, M. Ribosome Biogenesis in Plants: From Functional 45S Ribosomal DNA Organization to Ribosome Assembly Factors. Plant Cell 2019, 31, 1945–1967. [Google Scholar] [CrossRef] [Green Version]
- Campalans, A.; Kondorosi, A.; Crespi, M. Enod40, a Short Open Reading Frame–Containing mRNA, Induces Cytoplasmic Localization of a Nuclear RNA Binding Protein in Medicago truncatula. Plant Cell 2004, 16, 1047–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucero, L.; Bazin, J.; Melo, J.; Ibanez, F.; Crespi, M.; Ariel, F. Evolution of the Small Family of Alternative Splicing Modulators Nuclear Speckle RNA-Binding Proteins in Plants. Genes 2020, 11, 207. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Xu, S.; Wiermer, M.; Zhang, Y.; Li, X. The cyclin L homolog MOS12 and the MOS4-associated complex are required for the proper splicing of plant resistance genes. Plant J. 2012, 70, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Palma, K.; Zhao, Q.; Cheng, Y.; Bi, D.; Monaghan, J.; Cheng, W.; Zhang, Y.; Li, X. Regulation of plant innate immunity by three proteins in a complex conserved across the plant and animal kingdoms. Genes Dev. 2007, 21, 1484–1493. [Google Scholar] [CrossRef] [Green Version]
- Monaghan, J.; Xu, F.; Gao, M.; Zhao, Q.; Palma, K.; Long, C.; Chen, S.; Zhang, Y.; Li, X. Two Prp19-Like U-Box Proteins in the MOS4-Associated Complex Play Redundant Roles in Plant Innate Immunity. PLoS Pathog. 2009, 5, e1000526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staiger, D. RNA-Binding Proteins and Circadian Rhythms in Arabidopsis thaliana. Philos. Trans. R. Soc. Lond. 2001, 356, 1755–1759. [Google Scholar] [CrossRef] [Green Version]
- Schoning, J.; Streitner, C.; Page, D.; Hennig, S.; Uchida, K.; Wolf, E.; Furuya, M.; Staiger, D. Auto-regulation of the circadian slave oscillator component AtGRP7 and regulation of its targets is impaired by a single RNA recognition motif point mutation. Plant J. 2007, 52, 1119–1130. [Google Scholar] [CrossRef] [Green Version]
- Steffen, A.; Elgner, M.; Staiger, D. Regulation of Flowering Time by the RNA-Binding Proteins AtGRP7 and AtGRP8. Plant Cell Physiol. 2019, 60, 2040–2050. [Google Scholar] [CrossRef]
- Streitner, C.; Hennig, L.; Korneli, C.; Staiger, D. Global transcript profiling of transgenic plants constitutively overexpressing the RNA-binding protein AtGRP7. BMC Plant Biol. 2010, 10, 221. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.; Guo, M.; Jeong, B.; Tian, F.; Elthon, T.; Cerny, R.; Staiger, D.; Alfano, J. A type III effector ADP-ribosylates RNA-binding proteins and quells plant immunity. Nature 2007, 447. [Google Scholar] [CrossRef]
- Zuo, Y.; Feng, F.; Qi, W.; Song, R. Dek42 encodes an RNA-binding protein that affects alternative pre-mRNA splicing and maize kernel development. J. Integr. Plant Biol. 2019, 61, 728–748. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Manley, J. Alternative polyadenylation of mRNA precursors. Nat. Rev. Mol. Cell Biol. 2017, 18, 18–30. [Google Scholar] [CrossRef]
- Xing, D.; Li, Q. Alternative polyadenylation and gene expression regulation in plants. Wires RNA 2011, 2, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Hornyik, C.; Terzi, L.; Simpson, G. The Spen Family Protein FPA Controls Alternative Cleavage and Polyadenylation of RNA. Dev. Cell 2010, 18, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Sonmez, C.; Baurle, I.; Magusin, A.; Dreos, R.; Laubinger, S.; Weigel, D.; Dean, C. RNA 3′ processing functions of Arabidopsis FCA and FPA limit intergenic transcription. PNAS 2011, 108, 8508–8513. [Google Scholar] [CrossRef] [Green Version]
- Lyons, R.; Iwase, A.; Gansewig, T.; Sherstnev, A.; Duc, C.; Barton, G.; Hanada, K.; Higuchi-Takeuchi, M.; Matsui, M.; Sugimoto, K.; et al. The RNA-binding protein FPA regulates flg22-triggered defense responses and transcription factor activity by alternative polyadenylation. Sci. Rep. 2013, 3, 2866. [Google Scholar] [CrossRef]
- Crofts, A.; Crofts, N.; Whitelegge, J.; Okita, T. Isolation and identification of cytoskeleton-associated prolamine mRNA binding proteins from developing rice seeds. Planta 2010, 231, 1261–1276. [Google Scholar] [CrossRef]
- Tian, L.; Chou, H.; Fukuda, M.; Kumamaru, T.; Okita, T. mRNA Localization in Plant Cells. Plant Physiol. 2020, 182, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, L.; Chou, H.; Zhang, L.; Hwang, S.; Starkenburg, S.; Doroshenk, K.; Kumamaru, T.; Okita, T. RNA-Binding Protein RBP-P Is Required for Glutelin and Prolamine mRNA Localization in Rice Endosperm Cells. Plant Cell 2018, 30, 2529–2552. [Google Scholar] [CrossRef] [Green Version]
- Monaghan, J.; Germain, H.; Weihmann, T.; Li, X. Dissecting plant defence signal transduction: Modifiers of snc1 in Arabidopsis. Can. J. Plant Pathol. 2010, 32, 35–42. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, Y.; Bi, D.; Palma, K.; Li, X. MOS2, a Protein Containing G-Patch and KOW Motifs, Is Essential for Innate Immunity in Arabidopsis thaliana. Curr. Biol. 2005, 15, 1936–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, X. A Putative Nucleoporin 96 is Required for Both Basal Defense and Constitutive Resistance Responses Mediated by suppressor of npr1-1, constitutive 1. Plant Cell 2005, 17, 1306–1316. [Google Scholar] [CrossRef] [Green Version]
- Germain, H.; Qu, N.; Cheng, Y.; Lee, E.; Huang, Y.; Dong, O.; Gannon, P.; Huang, S.; Ding, P.; Li, Y.; et al. MOS11, A New Component in the mRNA Export Pathway. PLoS Genet. 2010, 6, e1001250. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Liu, S.; Tang, D. HPR1, a component of the THO/TREX complex, plays an important role in disease resistance and senescence in Arabidopsis. Plant J. 2012, 69, 831–843. [Google Scholar] [CrossRef]
- Parry, G.; Ward, S.; Cernac, A.; Dharmasiri, S.; Estelle, M. The Arabidopsis Suppressor of Auxin Resistance Proteins are Nucleoporins with an Important Role in Hormone Signaling and Development. Plant Cell 2006, 18, 1590–1603. [Google Scholar] [CrossRef] [Green Version]
- Lucas, W.; Yoo, B.; Kragler, F. RNA as a long-distance information macromolecule in plants. Nat. Rev. 2001, 2, 849–887. [Google Scholar] [CrossRef]
- Li, P.; Ham, B.; Lucas, W. CmRBP50 Protein Phosphorylation Is Essential for Assembly of a Stable Phloem-mobile High-affinity Ribonucleoprotein Complex. J. Biol. Chem. 2011, 286, 23142–23149. [Google Scholar] [CrossRef] [Green Version]
- Ham, B.; Brandom, J.; Xoconostle-Cázares, B.; Ringgold, V.; Lough, T.; Lucas, W. A Polypyrimidine Tract Binding Protein, Pumpkin RBP50, Forms the Basis of a Phloem-Mobile Ribonucleoprotein Complex. Plant Cell 2009, 21, 197–215. [Google Scholar] [CrossRef] [Green Version]
- Czolpinska, M.; Rurek, M. Plant Glycine-Rich Proteins in Stress Response: An Emerging, Still Prospective Story. Front. Plant Sci. 2018, 9, 302. [Google Scholar] [CrossRef]
- Wang, B.; Wang, G.; Shen, F.; Zhu, S. A Glycine-Rich RNA-Binding Protein, CsGR-RBP3, Is Involved in Defense Responses against Cold Stress in Harvested Cucumber (Cucumis sativus L.) Fruit. Front. Plant Sci. 2018, 9, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Kim, J.; Kang, H. Cold-inducible zinc finger-containing glycine-rich RNA-binding protein contributes to the enhancement of freezing tolerance in Arabidopsis thaliana. Plant J. 2005, 42, 890–900. [Google Scholar] [CrossRef]
- Kwak, K.; Kim, Y.; Kang, H. Characterization of transgenic Arabidopsis plants overexpressing GR-RBP4 under high salinity, dehydration, or cold stress. J. Exp. Bot. 2005, 56, 3007–3016. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yu, M.; Zou, C.; Lu, C.; Yu, D.; Cheng, H.; Jiang, P.; Feng, X.; Zhang, Y.; Wang, Q.; et al. Genome-wide comparative analysis of RNA binding Glycine-rich protein family genes between Gossypium arboreum and Gossypium raimondii. PLoS ONE 2019, 14, e0218938. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Cheng, Y.; Gao, M.; Li, M.; Xu, X. Molecular Characterization, Expression Pattern and Function Analysis of Glycine-Rich Protein Genes Under Stresses in Chinese Cabbage (Brassica rapa L. ssp. pekinensis). Front. Genet. 2020, 11, 774. [Google Scholar] [CrossRef] [PubMed]
- Melencion, S.; Chi, Y.; Pham, T.; Paeng, S.; Wi, S.; Lee, C.; Ryu, S.; Koo, S.; Lee, S. RNA Chaperone Function of a Universal Stress Protein in Arabidopsis Confers Enhanced Cold Stress Tolerance in Plants. Int. J. Mol. Sci. 2017, 18, 2546. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Abbasi, N.; Choi, S. Bruno-like proteins modulate flowering time via 3′ UTR-dependent decay of SOC1 mRNA. New Phytol. 2013, 198, 747–756. [Google Scholar] [CrossRef]
- Pfalz, J.; Bayraktar, O.; Prikryl, J.; Barkan, A. Site-specific binding of a PPR protein defines and stabilizes 50 and 30 mRNA termini in chloroplasts. EMBO J. 2009, 28, 2042–2052. [Google Scholar] [CrossRef]
- Barkan, A.; Small, I. Pentatricopeptide Repeat Proteins in Plants. Annu. Rev. Plant Biol. 2014, 65, 415–442. [Google Scholar] [CrossRef]
- Ruwe, H.; Wang, G.; Gusewski, S.; Schmitz-Linneweber, C. Systematic analysis of plant mitochondrial and chloroplast small RNAs suggests organelle-specific mRNA stabilization mechanisms. Nucleic Acids Res. 2016, 44. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Lee, K.; Lee, K.; Jung, S.; Jeon, Y.; Pai, H.; Kang, H. A nuclear-encoded chloroplast-targeted S1 RNA-binding domain protein affects chloroplast rRNA processing and is crucial for the normal growth of Arabidopsis thaliana. Plant J. 2015, 83, 277–289. [Google Scholar] [CrossRef]
- Dinh, S.; Park, S.; Han, J.; Kang, H. A Chloroplast-targeted S1 RNA-binding Domain Protein Plays a Role in Arabidopsis Response to Diverse Abiotic Stresses. J. Plant Biol. 2019, 62, 74–81. [Google Scholar] [CrossRef]
- Muthuramalingam, M.; Wang, Y.; Mahalingam, R. Interacting protein partners of Arabidopsis RNA-binding protein AtRBP45b. Plant Biol. 2016, 19, 327–334. [Google Scholar] [CrossRef]
- Park, S.; Jeong, J.; Redillas, M.; Jung, H.; Bang, S.; Kim, Y.; Kim, J. Transgenic overexpression of UIP1, an interactor of the 30 untranslated region of the Rubisco small subunit mRNA, increases rice tolerance to drought. Plant Biotechnol. Rep. 2013, 7, 83–90. [Google Scholar] [CrossRef]
- Brodersen, P.; Sakvarelidze-Achard, L.; Bruun-Rasmussen, M.; Dunoyer, P.; Yamamoto, Y.; Sieburth, L.; Voinnet, O. Widespread Translational Inhibition by Plant miRNAs and siRNAs. Science 2008, 320, 1185–1189. [Google Scholar] [CrossRef]
- Ren, G.; Chen, X.; Yu, B. Uridylation of miRNAs by HEN1 SUPPRESSOR1 in Arabidopsis. Curr. Biol. 2012, 22, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yu, Y.; Zhai, J.; Ramachandran, V.; Dinh, T.; Meyers, B.; Mo, B.; Chen, X. The Arabidopsis Nucleotidyl Transferase HESO1 Uridylates Unmethylated Small RNAs to Trigger Their Degradation. Curr. Biol. 2012, 22, 689–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, S.; Paek, K. Role of Arabidopsis Pumilio RNA binding protein 5 in virus infection. Plant Signal. Behav. 2013, 8, e23975. [Google Scholar] [CrossRef] [Green Version]
- Fujisaki, K.; Ishikawa, M. Identification of an Arabidopsis thaliana protein that binds to tomato mosaic virus genomic RNA and inhibits its multiplication. Virology 2008, 380, 402–411. [Google Scholar] [CrossRef] [Green Version]
- Jakubiec, A.; Yang, S.; Chua, N. Arabidopsis DRB4 protein in antiviral defense against Turnip yellow mosaic virus infection. Plant J. 2012, 69, 14–25. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Kim, K.; Shin, R.; Park, J.; Shin, Y.; Paek, K. Pathogenesis-related protein 10 isolated from hot pepper functions as a ribonuclease in an antiviral pathway. Plant J. 2004, 37, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Kim, M.; Kang, H. An ABA-regulated putative RNA-binding protein affects seed germination of Arabidopsis under ABA or abiotic stress conditions. J. Plant Physiol. 2013, 170, 179–184. [Google Scholar] [CrossRef]
- Gu, L.; Jung, H.; Kwak, K.; Dinh, S.; Kim, Y.; Kang, H. An RRM-containing mei2-like MCT1 plays a negative role in the seed germination and seedling growth of Arabidopsis thaliana in the presence of ABA. Plant Physiol. Biochem. 2016, 109, 273–279. [Google Scholar] [CrossRef]
- Tan, Y.; Qin, Y.; Li, Y.; Li, M.; Ma, F. Overexpression of MpGR-RBP1, a glycine-rich RNA-binding protein gene from Malus prunifolia (Willd.) Borkh., confers salt stress tolerance and protects against oxidative stress in Arabidopsis. Plant Cell Tissue Organ Cult. 2014, 119, 635–646. [Google Scholar] [CrossRef]
- Ben Saad, R.; Ben Halima, N.; Ghorbel, M.; Zouari, N.; Ben Romdhane, W.; Guiderdoni, E.; Al-Doss, A.; Hassairi, A. AlSRG1, a novel gene encoding an RRM-type RNA-binding protein (RBP) from Aeluropus littoralis, confers salt and drought tolerance in transgenic tobacco. Environ. Exp. Bot. 2018, 150, 25–36. [Google Scholar] [CrossRef]
- Kim, H.; Lee, J.; Kim, A.; Park, S.; Ma, S.; Lee, S.; Joung, Y. Heterologous expression of an RNA-binding protein affects flowering time as well as other developmental processes in Solanaceae. Mol. Breed. 2016, 36, 71. [Google Scholar] [CrossRef]
- Moshelion, M.; Altman, A. Current challenges and future perspectives of plant and agricultural biotechnology. Trends Biotechnol. 2015, 33, 337–341. [Google Scholar] [CrossRef]
- Yang, D.; Kwak, K.; Kim, M.; Park, S.; Yang, K.; Kang, H. Expression of Arabidopsis glycine-rich RNA-binding protein AtGRP2 or AtGRP7 improves grain yield of rice (Oryza sativa) under drought stress conditions. Plant Stress 2013, 214, 106–112. [Google Scholar] [CrossRef]
- Wang, S.; Wang, R.; Liang, D.; Ma, F.; Shu, H. Molecular characterization and expression analysis of a glycine-rich RNA-binding protein gene from Malus hupehensis Rehd. Mol. Biol. Rep. 2012, 39, 4145–4153. [Google Scholar] [CrossRef]
- Castiglioni, P.; Warner, D.; Bensen, R.; Anstrom, D.; Harrison, J.; Stoecker, M.; Abad, M.; Kumar, G.; Salvador, S.; D’Ordine, R.; et al. Bacterial RNA Chaperones Confer Abiotic Stress Tolerance in Plants and Improved Grain Yield in Maize under Water-Limited Conditions. Plant Physiol. 2008, 147, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemali, K.; Bonin, C.; Dohleman, F.; Stephens, M.; Reeves, W.; Nelson, D.; Castiglioni, P.; Whitsel, J.; Sammons, B.; Silady, R.; et al. Physiological responses related to increased grain yield under drought in the first biotechnology-derived drought-tolerant maize. Plant Cell Environ. 2015, 38, 1866–1880. [Google Scholar] [CrossRef] [Green Version]
- Téllez, S.; Kanhonou, R.; Bellés, C.; Serrano, R.; Alepuz, P.; Ros, R. RNA-Binding Proteins as Targets to Improve Salt Stress Tolerance in Crops. Agronomy 2020, 10, 250. [Google Scholar] [CrossRef] [Green Version]
- Ambrosone, A.; Batelli, G.; Nurcato, R.; Aurilia, V.; Punzo, P.; Bangarusamy, D.; Ruberti, I.; Sassi, M.; Leone, A.; Costa, A.; et al. The Arabidopsis RNA-Binding Protein AtRGGA Regulates Tolerance to Salt and Drought Stress. Plant Physiol. 2015, 168, 292–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.; Wang, N.; Xue, Y.; Guan, Q.; van Nocker, S.; Liu, C.; Ma, F. Overexpression of the RNA binding protein MhYTP1 in transgenic apple enhances drought tolerance and WUE by improving ABA level under drought condition. Plant Sci. 2019, 280, 397–407. [Google Scholar] [CrossRef]
- Mergner, J.; Frejno, M.; List, M.; Papacek, M.; Chen, X.; Chaudhary, A.; Samaras, P.; Richter, S.; Shikata, H.; Messerer, M.; et al. Mass-spectrometry-based draft of the Arabidopsis proteome. Nature 2020, 579, 409–414. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Pros | Cons | Plant Refs |
|---|---|---|---|
| Protein-to-RNA | |||
| RIP-seq | No genetic trans., reversible crosslinking, well established in plants, no radiolabeling | Uses antibody–antigen interaction, non-specific crosslinking, large amounts of starting material, no info on RBP site | He et al., 2009, Streitner et al., 2012; Yin et al., 2012; Rowley et al., 2013; Bardou et al., 2014; Francisco-Mangilet et al., 2015; Xing et al., 2015; Bazin et al., 2018; Marmisolle et al., 2018; Schmid et al., 2019; Tian et al., 2019 |
| CLIP-seq | No genetic trans., provides info. on the RBP binding site, well-established in plants | Uses antibody–antigen interaction, uses radiolabeling, large amounts of starting material | Meyer et al., 2017; Zhang et al., 2015 |
| HITS-CLIP | Increased coverage | As CLIP-seq | Zhang et al., 2015 |
| PAR-CLIP | More efficient UV-crosslinking | As CLIP-seq, favors certain RBP-RNA interactions | None |
| iCLIP | Increases precision of RBP site prediction | As CLIP-seq | Meyer et al., 2017 |
| dCLIP | Permits comparisons across all CLIP exps. | As CLIP-seq | None |
| uvCLAP | Tight binding affinity, uniform pulldown efficiency, quantify background, no radiolabeling, no antibodies | Not in plants, needs genetic trans., may alter RNA–protein interactions, no info on RBP site, large amounts of starting material | None |
| TRIBE/HyperTRIBE | No pull down, small amounts of starting material, no radiolabeling, no antibodies | Not in plants, needs genetic trans., editing occurs in a wide range around the binding site, no info on RBP site | None |
| RNA-to-Protein | |||
| ChIRP-MS/RAP-MS | High affinity interaction, no genetic trans., no radiolabeling, no antibodies | Not in plants, no info on RBP site, large amounts of starting material | None |
| RNA Small Molecule Labeling | No genetic trans., no radiolabeling, no antibodies | In vitro only | None |
| RNA Nucleotide Substition | No genetic trans., no radiolabeling, no antibodies | In vitro only | None |
| RNA Aptamer Pulldown | High affinity interaction, many aptamers, no radiolabeling, no antibodies | Not in plants, needs genetic trans., no info on RBP site, may alter RNA–protein interactions, large amounts of starting material, may be prone to aggregation | None |
| RBP | Plant System | Method | Number of RNA Targets | References |
|---|---|---|---|---|
| AGO4 | Arabidopsis thaliana | RIP | 2 | Wierzbicki et al., 2009 |
| AtGRP7 | Arabidopsis thaliana | RIP-seq/iCLIP | 452/858 | Streitner et al., 2012; Meyer et al., 2017 |
| AtNSRa | Arabidopsis thaliana | RIP-seq | >2000 | Bardou et al., 2014; Bazin et al., 2018 |
| AtNSRb | Arabidopsis thaliana | RIP-seq | >2000 | Bardou et al., 2014; Bazin et al., 2018 |
| CPsV 24K (viral) | Nicotiana benthamiana | RIP | 2 | Marmisolle et al., 2018 |
| CPsV 24K (viral) | Nicotiana benthamiana | RIP | 2 | Marmisolle et al., 2018 |
| CSP1 | Arabidopsis thaliana | RIP-chip | >6000 | Juntawong et al., 2013 |
| IDN2 | Arabidopsis thaliana | RIP | 1 | Zhu et al., 2013 |
| FCA | Arabidopsis thaliana | RIP | 1 | Tian et al., 2019 |
| HLP1 | Arabidopsis thaliana | HITS-CLIP | >5000 | Zhang et al., 2015 |
| KTF1 | Arabidopsis thaliana | RIP | 1 | He et al., 2009 |
| NSF | Oryza sativa | RIP | ? | Tian et al., 2020 |
| PUMPKIN | Arabidopsis thaliana | RIP-seq | 5 | Schmid et al., 2019 |
| PDM1 | Arabidopsis thaliana | RIP | 1 | Yin et al., 2012 |
| Rab5a | Oryza sativa | RIP | ? | Tian et al., 2020 |
| RBP-L | Oryza sativa | RIP | ? | Tian et al., 2020 |
| RBP-P | Oryza sativa | RIP | ? | Tian et al., 2020 |
| SR45 | Arabidopsis thaliana | RIP-seq | >4000/>1800 | Xing et al., 2015; Zhang et al., 2017 |
| THO2 | Arabidopsis thaliana | RIP | 6 | Francisco-Mangilet et al., 2015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burjoski, V.; Reddy, A.S.N. The Landscape of RNA-Protein Interactions in Plants: Approaches and Current Status. Int. J. Mol. Sci. 2021, 22, 2845. https://doi.org/10.3390/ijms22062845
Burjoski V, Reddy ASN. The Landscape of RNA-Protein Interactions in Plants: Approaches and Current Status. International Journal of Molecular Sciences. 2021; 22(6):2845. https://doi.org/10.3390/ijms22062845
Chicago/Turabian StyleBurjoski, Vesper, and Anireddy S. N. Reddy. 2021. "The Landscape of RNA-Protein Interactions in Plants: Approaches and Current Status" International Journal of Molecular Sciences 22, no. 6: 2845. https://doi.org/10.3390/ijms22062845
APA StyleBurjoski, V., & Reddy, A. S. N. (2021). The Landscape of RNA-Protein Interactions in Plants: Approaches and Current Status. International Journal of Molecular Sciences, 22(6), 2845. https://doi.org/10.3390/ijms22062845