
Muscle Damage in Systemic Sclerosis and CXCL10: The Potential Therapeutic Role of PDE5 Inhibition

 , ,
, , 
 and
and
Abstract
:1. Introduction
2. Results
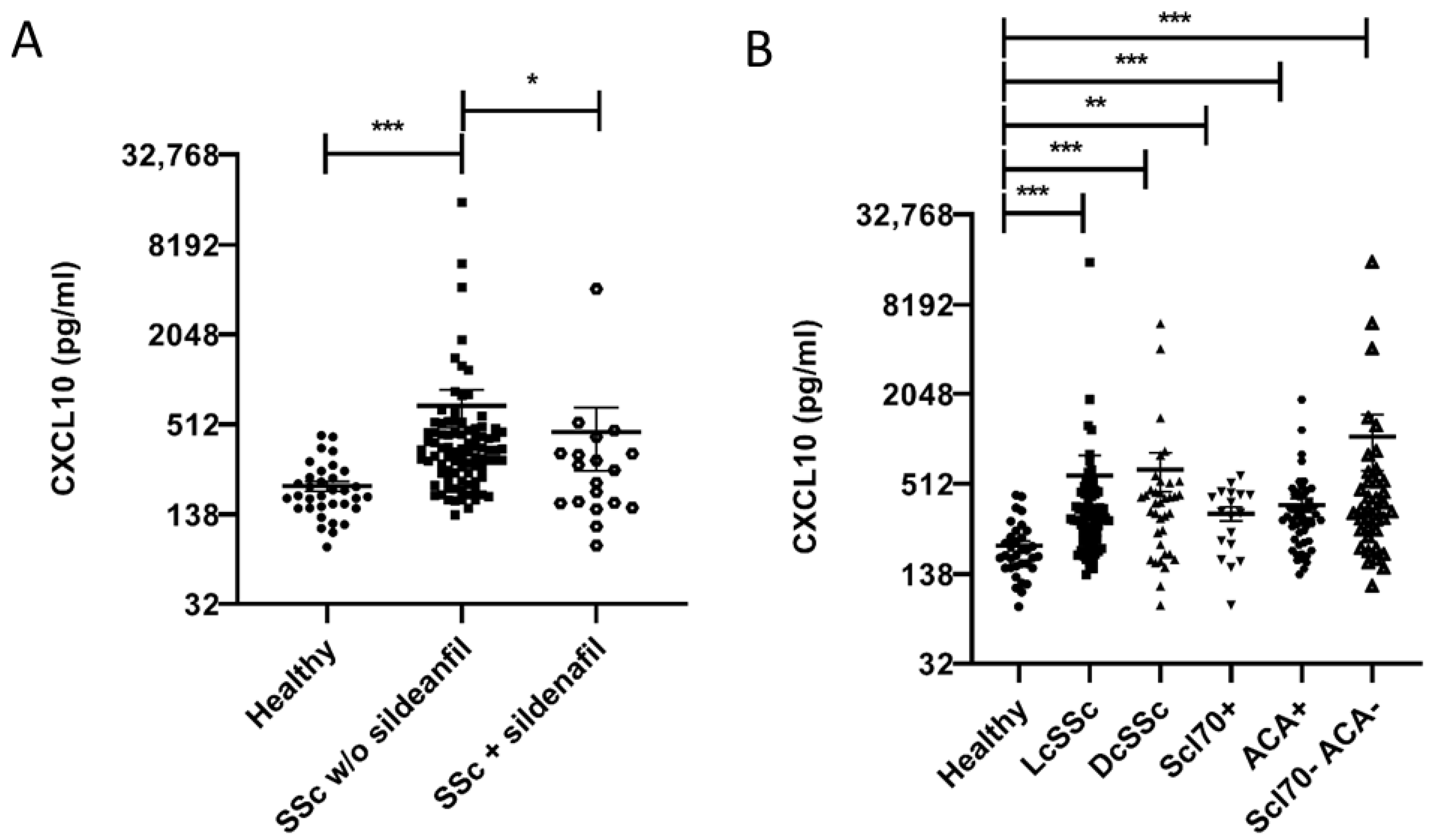
2.1. CXCL10 Serum Level Is Higher in SSc vs. Healthy Subjects and Correlates Inversely with Sildenafil Treatment
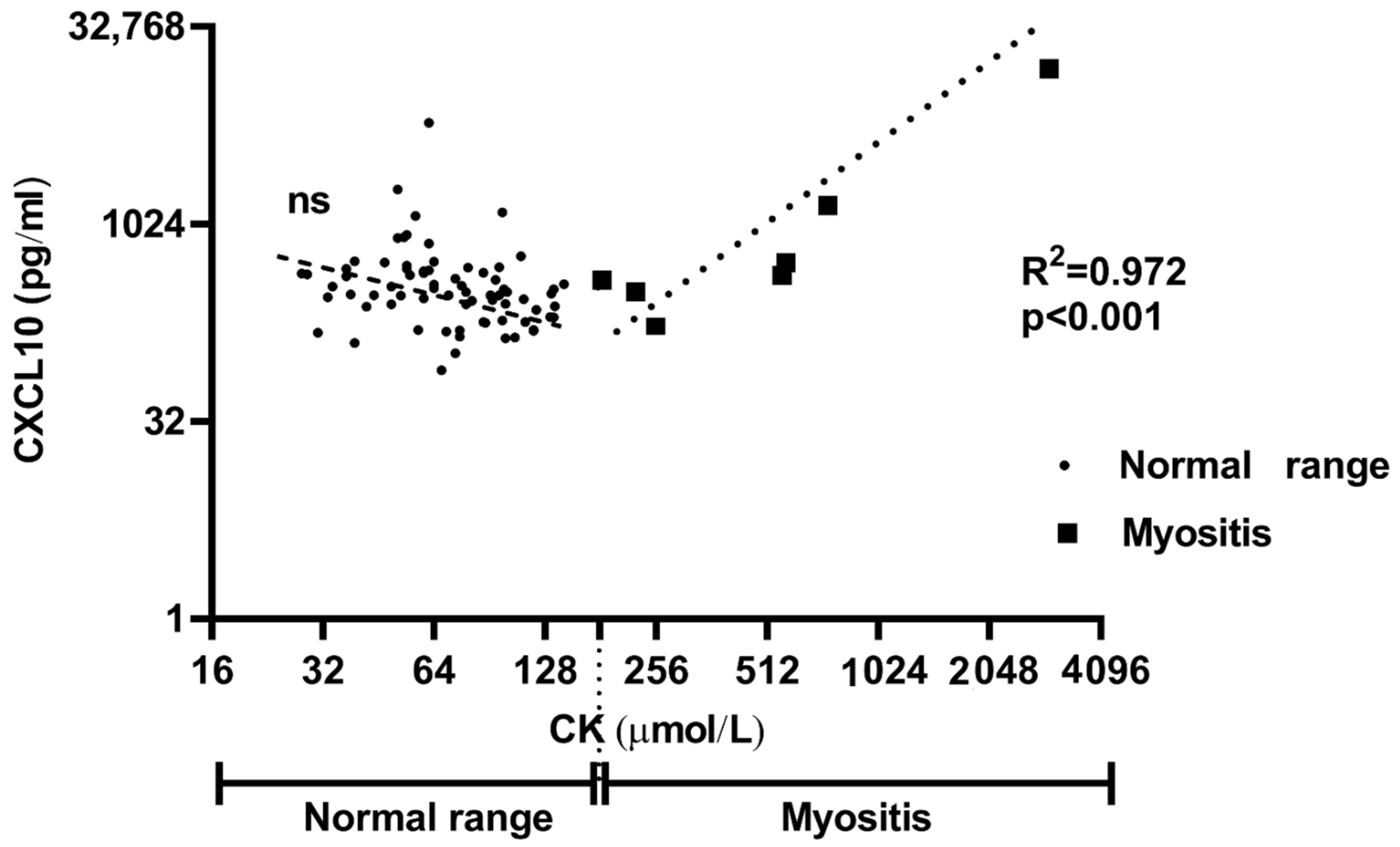
2.2. Patients with Skeletal Muscle Damage Have Higher CXCL10 Sera Levels
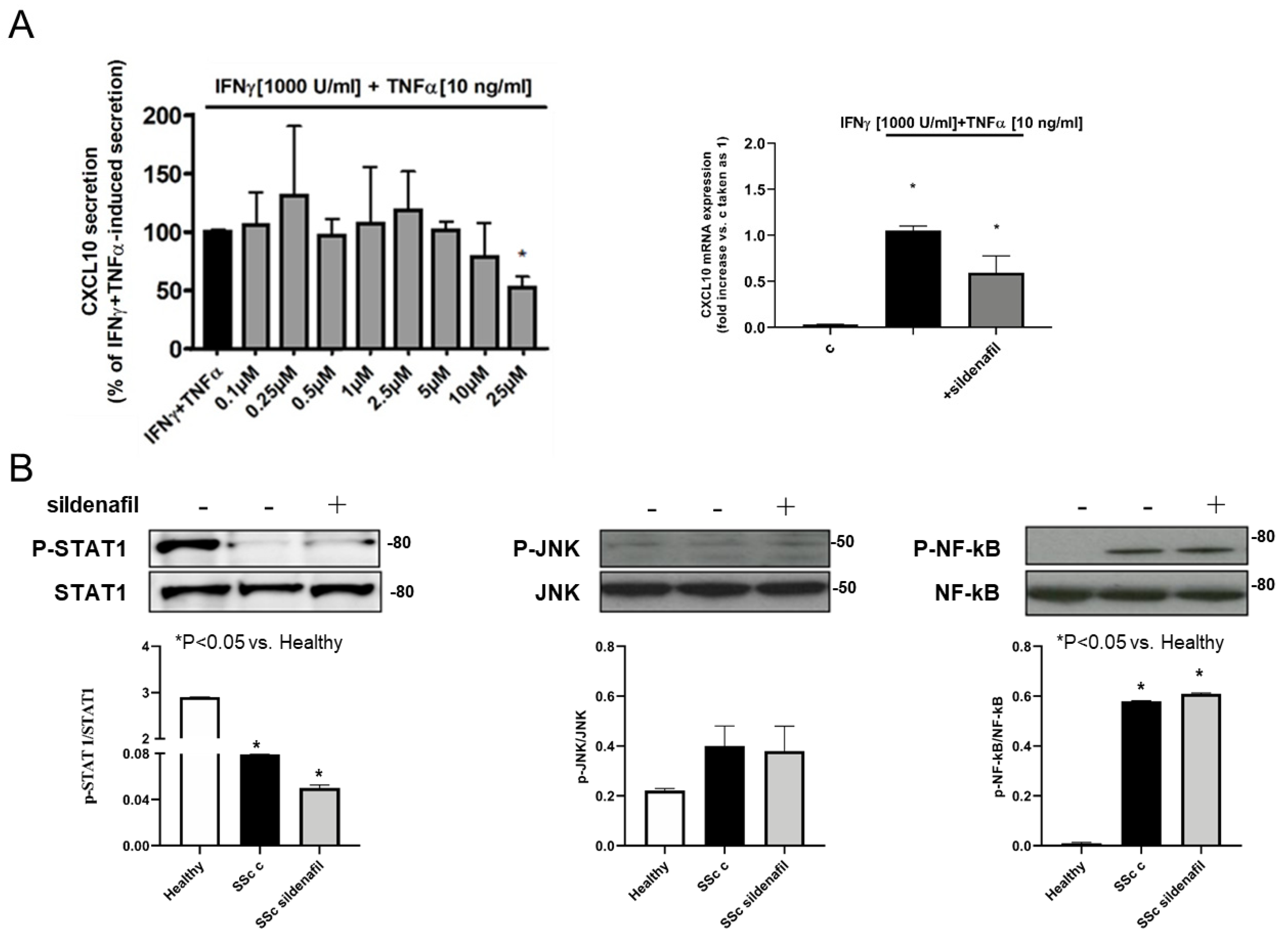
2.3. Sildenafil Suppresses Cytokine-Induced CXCL10 Secretion in Human Skeletal Muscle Cells, with No Effect on Healthy Human Fibroblasts
2.4. Sildenafil Inhibited STAT1, NF-kB and JNK Activation by Inflammatory Stimuli in Human Myocytes and Cardiomyocytes, Not in Human Normal Fibroblasts
2.5. SSc Fibroblast CXCL10 Secretion and STAT1, JNK and NF-kB Phosphorylation Levels Are Unaffected by Sildenafil Treatment
2.6. Sildenafil Did Not Affect Viability of Human Skeletal Muscle Cells, Normal or SSc Fibroblasts
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Chemicals
4.3. Cell Cultures
4.4. Cytokine Secretion Assay
4.5. Elisa Assays
4.6. Western Blot Analysis
4.7. RNA Extraction, Reverse Transcription and Real-Time Quantitative PCR
4.8. Cell Viability
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rabquer, B.J.; Tsou, P.S.; Hou, Y.; Thirunavukkarasu, E.; Haines, G.K., 3rd; Impens, A.J.; Phillips, K.; Kahaleh, B.; Seibold, J.R.; Koch, A.E. Dysregulated expression of MIG/CXCL9, IP-10/CXCL10 and CXCL16 and their receptors in systemic sclerosis. Arthritis Res. Ther. 2011, 13, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, H.; Shimada, Y.; Hasegawa, M.; Takehara, K.; Sato, S. Serum levels of a Th1 chemoattractant IP-10 and Th2 chemoattractants, TARC and MDC, are elevated in patients with systemic sclerosis. J. Dermatol. Sci. 2014, 35, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Kong, K.O.; Tan, A.W.; Thong, B.Y.; Lian, T.Y.; Cheng, Y.K.; Teh, C.L.; Koh, E.T.; Chng, H.H.; Law, W.G.; Lau, T.C.; et al. Enhanced expression of interferon-inducible protein-10 correlates with disease activity and clinical manifestations in systemic lupus erythematosus. Clin. Exp. Immunol. 2009, 156, 134–140. [Google Scholar] [CrossRef]
- Martini, G.; Zulian, F.; Calabrese, F.; Bortoli, M.; Facco, M.; Cabrelle, A.; Valente, M.; Zacchello, F.; Agostini, C. CXCR3/CXCL10 expression in the synovium of children with juvenile idiopathic arthritis. Arthritis Res. Ther. 2005, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- De Paepe, B.; Creus, K.K.; De Bleecker, J.L. Chemokine profile of different inflammatory myopathies reflects humoral versus cytotoxic immune responses. Ann N. Y. Acad. Sci. 2007, 1109, 441–453. [Google Scholar] [CrossRef]
- Crescioli, C.; Squecco, R.; Cosmi, L.; Sottili, M.; Gelmini, S.; Borgogni, E.; Sarchielli, E.; Scolletta, S.; Francini, F.; Annunziato, F.; et al. Immunosuppression in cardiac graft rejection: A human in vitro model to study the potential use of new immunomodulatory drugs. Exp. Cell Res. 2008, 314, 1337–1350. [Google Scholar] [CrossRef] [PubMed]
- Sottili, M.; Cosmi, L.; Borgogni, E.; Sarchielli, E.; Maggi, L.; Francalanci, M.; Vannelli, G.B.; Ronconi, E.; Adorini, L.; Annunziato, F.; et al. Immunomodulatory effects of BXL-01-0029, a less hypercalcemic vitamin D analogue, in human cardiomyocytes and T cells. Exp. Cell Res. 2009, 315, 264–273. [Google Scholar] [CrossRef]
- Lacotte, S.; Brun, S.; Muller, S.; Dumortier, H. CXCR3, inflammation, and autoimmune diseases. Ann. N. Y. Acad. Sci. 2009, 1173, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Scolletta, S.; Colletti, M.; Di Luigi, L.; Crescioli, C. Vitamin D receptor agonists target CXCL10: New therapeutic tools for resolution of inflammation. Mediat. Inflamm. 2013, 876319. [Google Scholar] [CrossRef] [Green Version]
- Crescioli, C.; Corinaldesi, C.; Riccieri, V.; Raparelli, V.; Vasile, M.; Del Galdo, F.; Valesini, G.; Lenzi, A.; Basili, S.; Antinozzi, C. Association of circulating CXCL10 and CXCL11 with systemic sclerosis. Ann. Rheum. Dis. 2018, 77, 1845–1846. [Google Scholar] [CrossRef] [Green Version]
- Cossu, M.; van Bon, L.; Preti, C.; Rossato, M.; Beretta, L.; Radstake, T.R.D.J. Earliest Phase of Systemic Sclerosis Typified by Increased Levels of Inflammatory Proteins in the Serum. Arthritis Rheumatol. 2017, 69, 2359–2369. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Mayes, M.D.; Tan, F.K.; Wu, M.; Reveille, J.D.; Harper, B.E.; Draeger, H.T.; Gonzalez, E.B.; Assassi, S. Correlation of interferon-inducible chemokine plasma levels with disease severity in systemic sclerosis. Arthritis Rheum. 2013, 65, 226–235. [Google Scholar] [CrossRef] [Green Version]
- Medsger, T.A., Jr.; Bombardieri, S.; Czirjak, L.; Scorza, R.; Della Rossa, A.; Bencivelli, W. Assessment of disease severity and prognosis. Clin. Exp. Rheumatol. 2003, 21, S42–S46. [Google Scholar] [PubMed]
- Ranque, B.; Authier, F.J.; Berezne, A.; Guillevin, L.; Mouthon, L. Systemic sclerosis-associated myopathy. Ann. N. Y. Acad. Sci. 2007, 1108, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M. Biomarkers in systemic sclerosis: Their potential to predict clinical courses. J. Dermatol. 2016, 43, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Parks, J.L.; Taylor, M.H.; Parks, L.P.; Silver, R.M. Systemic sclerosis and the heart. Rheum. Dis. Clin. 2014, 40, 87–102. [Google Scholar] [CrossRef]
- Di Luigi, L.; Corinaldesi, C.; Colletti, M.; Scolletta, S.; Antinozzi, C.; Vannelli, G.B.; Giannetta, E.; Gianfrilli, D.; Isidori, A.M.; Migliaccio, S.; et al. Phosphodiesterase Type 5 Inhibitor Sildenafil Decreases the Proinflammatory Chemokine CXCL10 in Human Cardiomyocytes and in Subjects with Diabetic Cardiomyopathy. Inflammation 2016, 39, 1238–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altara, R.; Mallat, Z.; Booz, G.W.; Zouein, F.A. The CXCL10/CXCR3 Axis and Cardiac Inflammation: Implications for Immunotherapy to Treat Infectious and Noninfectious Diseases of the Heart. J. Immunol. Res. 2016, 4396368. [Google Scholar] [CrossRef] [Green Version]
- Bellando Randone, S.; George, J.; Mazzotta, C.; Guiducci, S.; Furst, D.E.; Mor, A.; Matucci, C.M. Angiostatic and Angiogenic Chemokines in Systemic Sclerosis: An Overview. J. Scleroderma Relat. Disord. 2017, 2, 1–10. [Google Scholar] [CrossRef]
- Goldstein, I.; Lue, T.F.; Padma-Nathan, H.; Rosen, R.C.; Steers, W.D.; Wicker, P.A. Oral sildenafil in the treatment of erectile dysfunction. Sildenafil Study Group. N. Engl. J. Med. 1998, 338, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Galiè, N.; Ghofrani, H.A.; Torbicki, A.; Barst, R.J.; Rubin, L.J.; Badesch, D.; Fleming, T.; Parpia, T.; Burgess, G.; Branzi, A.; et al. Sildenafil Use in Pulmonary Arterial Hypertension (SUPER) Study Group. Sildenafil citrate therapy for pulmonary arterial hypertension. N. Engl. J. Med. 2005, 353, 2148–2157. [Google Scholar] [CrossRef] [Green Version]
- Schwab, N.; Waschbisch, A.; Wrobel, B.; Lochmuller, H.; Sommer, C.; Wiendl, H. Human myoblasts modulate the function of antigen-presenting cells. J. Neuroimmunol. 2008, 200, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.T.; Chen, B.P.; Oertel, P.; Buchmeier, M.J.; Armstrong, D.; Hamilton, T.A.; Lane, T.E. The T cell chemoattractant IFN-inducible protein 10 is essential in host defense against viral-induced neurologic disease. J. Immunol. 2000, 165, 2327–2330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farber, J.M. Mig and IP-10: CXC chemokines that target lymphocytes. J. Leukoc. Biol. 1997, 61, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Campanella, G.S.; Tager, A.M.; El Khoury, J.K.; Thomas, S.Y.; Abrazinski, T.A.; Manice, L.A.; Colvin, R.A.; Luster, A.D. Chemokine receptor CXCR3 and its ligands CXCL9 and CXCL10 are required for the development of murine cerebral malaria. Proc. Natl. Acad. Sci. USA 2008, 105, 4814–4819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crescioli, C.; Buonamano, A.; Scolletta, S.; Sottili, M.; Francalanci, M.; Giomarelli, P.; Biagioli, B.; Lisi, G.; Pradella, F.; Serio, M.; et al. Predictive role of pretransplant serum CXCL10 for cardiac acute rejection. Transplantation 2009, 87, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, P.; Crescioli, C. CXCL10: A candidate biomarker in transplantation. Clin. Chim. Acta. 2012, 413, 1364–1373. [Google Scholar] [CrossRef] [PubMed]
- van den Borne, P.; Quax, P.H.; Hoefer, I.E.; Pasterkamp, G. The multifaceted functions of CXCL10 in cardiovascular disease. Biomed Res. Int. 2014, 893106. [Google Scholar] [CrossRef]
- Scolletta, S.; Buonamano, A.; Sottili, M.; Giomarelli, P.; Biagioli, B.; Vannelli, G.B.; Serio, M.; Romagnani, P.; Crescioli, C. CXCL10 release in cardiopulmonary bypass: An in vivo and in vitro study. Biomed. Aging Pathol. 2012, 2, 187–194. [Google Scholar] [CrossRef]
- Campbell, J.D.; Gangur, V.; Simons, F.E.; HayGlass, K.T. Allergic humans are hyporesponsive to a CXCR3-ligand-mediated Th1 immunity-promoting loop. FASEB J. 2004, 18, 329–331. [Google Scholar] [CrossRef]
- Crescioli, C.; Sottili, M.; Bonini, P.; Cosmi, L.; Chiarugi, P.; Romagnani, P.; Vannelli, G.B.; Colletti, M.; Isidori, A.M.; Serio, M.; et al. Inflammatory response in human skeletal muscle cells: CXCL10 as a potential therapeutic target. Eur. J. Cell Biol. 2012, 91, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Gallo, J.; Raska, M.; Kriegova, E.; Goodman, S.B. Inflammation and its resolution and the musculoskeletal system. J. Orthop. Transl. 2017, 10, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Buckley, C.D.; Barone, F.; Nayar, S.; Bénézech, C.; Caamaño, J. Stromal cells in chronic inflammation and tertiary lymphoid organ formation. Annu. Rev. Immunol. 2015, 33, 715–745. [Google Scholar] [CrossRef]
- Eloranta, M.L.; Franck-Larsson, K.; Lövgren, T.; Kalamajski, S.; Rönnblom, A.; Rubin, K.; Alm, G.V.; Rönnblom, L. Type I interferon system activation and association with disease manifestations in systemic sclerosis. Ann. Rheum. Dis. 2010, 69, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Crescioli, C. Vitamin D receptor agonists: Suitable candidates as novel therapeutic options in autoimmune inflammatory myopathy. BioMed Res. Int. 2014, 949730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deyhle, M.R.; Hafen, P.S.; Parmley, J.; Preece, C.N.; Robison, M.; Sorensen, J.R.; Jackson, B.; Eggett, D.L.; Hancock, C.R.; Hyldahl, R.D. CXCL10 increases in human skeletal muscle following damage but is not necessary for muscle regeneration. Physiol. Rep. 2018, 6, e13689. [Google Scholar] [CrossRef]
- Follansbee, W.P.; Zerbe, T.R.; Medsger, T.A., Jr. Cardiac and skeletal muscle disease in systemic sclerosis (scleroderma): A high risk association. Am. Heart J. 1993, 125, 194–203. [Google Scholar] [CrossRef]
- West, S.G.; Killian, P.J.; Lawless, O.J. Association of myositis and myocarditis in progressive systemic sclerosis. Arthritis Rheum. 1981, 24, 662–668. [Google Scholar] [CrossRef]
- Walker, U.A.; Clements, P.J.; Allanore, Y.; Distler, O.; Oddis, C.V.; Khanna, D.; Furst, D.E. Muscle involvement in systemic sclerosis: Points to consider in clinical trials. Rheumatology 2017, 56, v38–v44. [Google Scholar] [CrossRef] [Green Version]
- Tay, T.; Ferdowsi, N.; Baron, M.; Stevens, W.; Hudson, M.; Proudman, S.M.; Nikpour, M. Damage Index Working Group of the Scleroderma Clinical Trials Consortium. Measures of disease status in systemic sclerosis: A systematic review. Semin. Arthritis Rheum. 2017, 46, 473–487. [Google Scholar] [CrossRef]
- Varma, A.; Das, A.; Hoke, N.N.; Durrant, D.E.; Salloum, F.N.; Kukreja, R.C. Anti-inflammatory and cardioprotective effects of tadalafil in diabetic mice. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Menniti, F.S.; Ren, J.; Coskran, T.M.; Liu, J.; Morton, D.; Sietsma, D.K.; Som, A.; Stephenson, D.T.; Tate, B.A.; Finklestein, S.P. Phosphodiesterase 5A inhibitors improve functional recovery after stroke in rats: Optimized dosing regimen with implications for mechanism. J. Pharm. Exp. Ther. 2009, 331, 842–850. [Google Scholar] [CrossRef]
- Pifarré, P.; Gutierrez-Mecinas, M.; Prado, J.; Usero, L.; Roura-Mir, C.; Giralt, M.; Hidalgo, J.; García, A. Phosphodiesterase 5 inhibition at disease onset prevents experimental autoimmune encephalomyelitis progression through immunoregulatory and neuroprotective actions. Exp. Neurol. 2014, 251, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Puzzo, D.; Staniszewski, A.; Deng, S.X.; Privitera, L.; Leznik, E.; Liu, S.; Zhang, H.; Feng, Y.; Palmeri, A.; Landry, D.W.; et al. Phosphodiesterase 5 inhibition improves synaptic function, memory, and amyloid-beta load in an Alzheimer’s disease mouse model. J. Neurosci. 2009, 29, 8075–8086. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Durrant, D.; Salloum, F.N.; Xi, L.; Kukreja, R.C. PDE5 inhibitors as therapeutics for heart disease, diabetes and cancer. Pharmacol. Ther. 2015, 147, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Corinaldesi, C.; Di Luigi, L.; Lenzi, A.; Crescioli, C. Phosphodiesterase type 5 inhibitors: Back and forward from cardiac indications. J. Endocrinol. Investig. 2016, 39, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenoy, P.; Agarwal, V. Phosphodiesterase inhibitors in the management of autoimmune disease. Autoimmun. Rev. 2010, 9, 511–515. [Google Scholar] [CrossRef]
- Valentini, G.; Medsger, T.A., Jr.; Silman, A.J.; Bombardieri, S. Conclusion and identification of the core set of variables to be used in clinical investigations. Clin. Exp. Rheumatol. 2003, 21, S47–S48. [Google Scholar] [PubMed]
- LeRoy, A.; Avvedimento, E.V.; Krieg, T. Scleroderma. N. Engl J. Med. 2009, 360, 1989–2003. [Google Scholar] [CrossRef]
- Clements, P.J.; Lachenbruch, P.A.; Seibold, J.R.; Zee, B.; Steen, V.D.; Brennan, P.; Silman, A.J.; Allegar, N.; Varga, J.; Massa, M.; et al. Skin thickness score in systemic sclerosis: An assessment of interobserver variability in 3 independent studies. J. Rheumatol. 1993, 20, 1892–1896. [Google Scholar] [PubMed]
- Valentini, G.; Della Rossa, A.; Bombardieri, S.; Bencivelli, W.; Silman, A.J.; D’Angelo, S.; Cerinic, M.M.; Belch, J.F.; Black, C.M.; Bruhlmann, P.; et al. European multicentre study to define disease activity criteria for systemic sclerosis. II. Identification of disease activity variables and development of preliminary activity indexes. Ann. Rheum. Dis. 2001, 60, 592–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyer, C.; Distler, J.H.; Allanore, Y.; Aringer, M.; Avouac, J.; Czirják, L.; Cutolo, M.; Damjanov, N.; Del Galdo, F.; Fligelstone, K.; et al. EUSTAR Biobanking Group. (2011). EUSTAR biobanking: Recommendations for the collection, storage and distribution of biospecimens in scleroderma research. Ann. Rheum. Dis. 2011, 70, 1178–1182. [Google Scholar] [CrossRef] [PubMed]
- Del Galdo, F.; Sotgia, F.; de Almeida, C.J.; Jasmin, J.F.; Musick, M.; Lisanti, M.P.; Jiménez, S.A. Decreased expression of caveolin 1 in patients with systemic sclerosis: Crucial role in the pathogenesis of tissue fibrosis. Arthritis Rheum. 2008, 58, 2854–2865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillespie, J.; Ross, R.L.; Corinaldesi, C.; Esteves, F.; Derrett-Smith, E.; McDermott, M.F.; Doody, G.M.; Denton, C.P.; Emery, P.; Del Galdo, F. Transforming Growth Factor β Activation Primes Canonical Wnt Signaling Through Down-Regulation of Axin-2. Arthritis Rheumatol. 2018, 70, 932–942. [Google Scholar] [CrossRef] [PubMed]
- Mannelli, M.; Gelmini, S.; Arnaldi, G.; Becherini, L.; Bemporad, D.; Crescioli, C.; Pazzagli, M.; Mantero, F.; Serio, M.; Orlando, C. Telomerase activity is significantly enhanced in malignant adrenocortical tumors in comparison to benign adrenocortical adenomas. J. Clin. Endocrinol. Metab. 2000, 85, 468–470. [Google Scholar] [CrossRef] [PubMed]
- Mannelli, M.; Ferruzzi, P.; Luciani, P.; Crescioli, C.; Buci, L.; Corona, G.; Serio, M.; Peri, A. Cushing’s syndrome in a patient with bilateral macronodular adrenal hyperplasia responding to cisapride: An in vivo and in vitro study. J. Clin. Endocrinol. Metab. 2003, 88, 4616–4622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peri, A.; Luciani, P.; Conforti, B.; Baglioni-Peri, S.; Cioppi, F.; Crescioli, C.; Ferruzzi, P.; Gelmini, S.; Arnaldi, G.; Nesi, G.; et al. Variable expression of the transcription factors cAMP response element-binding protein and inducible cAMP early repressor in the normal adrenal cortex and in adrenocortical adenomas and carcinomas. J. Clin. Endocrinol. Metab. 2001, 86, 5443–5449. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | All n = 116 | Sildenafil n = 17 | No Sildenafil n = 99 | p |
|---|---|---|---|---|
| Male gender | 18 (15.5) | 4 (23.5) | 14 (15.5) | ns |
| Age, years | 57.2 (12.8) | 52.8 (11.9) | 57.9 (12.8) | ns |
| Disease duration from RP, years | 17.2 (14.6) | 20.1 (14.8) | 16.7 (14.6) | ns |
| Disease duration from 1st non RP, years | 10.2 (8.2) | 12.9 (6.8) | 9.8 (8.4) | ns |
| Disease subset (DcSSc) | 38 (32.8) | 8 (47.1) | 30 (30.3) | ns |
| ANA + | 113 (97.4) | 16 (94.1) | 97 (98) | ns |
| ACA + | 53 (45.7) | 8 (47.1) | 45 (45.5) | ns |
| Anti-topoisomerase I + | 18 (15.5) | 4 (23.5) | 14 (14.1) | ns |
| mRss | 2 (0–34) | 4 (0–16) | 2 (0–34) | ns |
| Raynaud’s Phenomenon | 110 (94.8) | 17 (100) | 93 | ns |
| Digital ulcers | 18 (15.5) | 8 (47.1) | 10 (10.2) | 0.0008 |
| Telangectasias | 90 (77.6) | 15 (88.2) | 75 (77.3) | ns |
| Synovitis | 9 (7.8) | 1 (5.9) | 8 (8.2) | ns |
| Flexion contractures | 52 (44.8) | 13 (76.5) | 39 (39.4) | 0.0071 |
| Tendon friction rubs | 5 (4.3) | 1 (5.9) | 4 (4) | ns |
| Proximal muscle weakness | 8 (6.9) | 0 (0) | 8 (8.1) | ns |
| Serum CK elevation | 5 (4.3) | 0 (0) | 5 (5) | ns |
| Reflux/dysphagia | 89 (76.7) | 12 (70.6) | 77 (77.8) | ns |
| Early satiety/vomiting | 31 (26.7) | 6 (35.3) | 25 (25) | ns |
| Diarrhoea/constipation/bloating | 32 (27.6) | 8 (47.1) | 24 (24.2) | ns |
| Dyspnoea | 69 (59.5) | 11 (64.7) | 56 (56.6) | ns |
| Chest HRCT fibrosis | 36 (50) | 7 (58.3) | 29 (48.3) | ns |
| Restrictive defect (FVC, DLCO) | 29(25.9) | 6 (42.9) | 23 (24) | ns |
| Confirmed PAH (by RHC) | 8 (6.9) | 4 (23.5) | 4 (4) | 0.0157 |
| Palpitations | 27 (23.3) | 7 (41.2) | 20 (20.2) | ns |
| Conduction defects | 1 (1) | 0 (0) | 1 (1.1) | ns |
| SV arrhythmias | 2 (1.9) | 1 (7.7) | 1 (1.1) | ns |
| V arrhythmias | 1 (1) | 0 (0) | 1 (1.1) | ns |
| Diastolic dysfunction | 59 (54.6) | 7 (41.2) | 52 (57.1) | ns |
| Reduced ejection fraction | 8 (7.1) | 1 (9.1) | 7 (7.4) | ns |
| Arterial hypertension | 17 (14.7) | 3 (17.7) | 14 (14.3) | ns |
| Renal crisis | 1 (0.9) | 0 (0) | 1 (1) | ns |
| SSc capillary pattern | 56 (90.3) | 11 (100) | 28 (54.9) | 0.0046 |
| - Early | 13 (20.9) | 1 (9.1) | 12 (23.5) | ns |
| - Active | 21 (33.9) | 4 (36.4) | 17 (33.3) | ns |
| - Late | 22 (35.5) | 6 (54.6) | 16 (31.4) | ns |
| EScSG-AI | 1.5 (0–7.5) | 1.5 (0–4) | 1 (0–7.5) | ns |
| Sev_general | 0 (0–3) | 0 (0–2) | 0 (0–3) | ns |
| Sev_peripheral vascular | 1 (1–3) | 2 (0–3) | 1 (0–3) | 0.0059 |
| Sev_skin | 1 (0–3) | 1 (0–2) | 1 (0–3) | ns |
| Sev_joint/tendon | 0 (0–4) | 1 (0–2) | 0 (0–4) | 0.03 |
| Sev_muscle | 0 (0–3) | 1 (0–2) | 0 (0–3) | <0.0001 |
| Sev_GI tract | 1 (0–2) | 1 (0–2) | 1 (0–2) | ns |
| Sev_lung | 2 (0–4) | 1 (0–2) | 2 (0–4) | 0.0003 |
| Sev_heart | 0 (0–3) | 1 (0–2) | 0 (0–3) | <0.0001 |
| Sev_kidney | 0 (0–1) | 1 (0–2) | 0 (0–1) | <0.0001 |
| Sev_total | 6 (2–19) | 9 (2–12) | 6 (2–19) | 0.047 |
| Immunosuppressive/anti-rheumatic therapy | 46 (39.7) | 5 (29.4) | 41 (35.3) | ns |
| Sildenafil | 17 (14.7) | 17 | 0 | <0.0001 |
| Bosentan | 4 | 3 | 1 | 0.0097 |
| Prostanoids | 30 | 7 | 23 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corinaldesi, C.; Ross, R.L.; Abignano, G.; Antinozzi, C.; Marampon, F.; di Luigi, L.; Buch, M.H.; Riccieri, V.; Lenzi, A.; Crescioli, C.; et al. Muscle Damage in Systemic Sclerosis and CXCL10: The Potential Therapeutic Role of PDE5 Inhibition. Int. J. Mol. Sci. 2021, 22, 2894. https://doi.org/10.3390/ijms22062894
Corinaldesi C, Ross RL, Abignano G, Antinozzi C, Marampon F, di Luigi L, Buch MH, Riccieri V, Lenzi A, Crescioli C, et al. Muscle Damage in Systemic Sclerosis and CXCL10: The Potential Therapeutic Role of PDE5 Inhibition. International Journal of Molecular Sciences. 2021; 22(6):2894. https://doi.org/10.3390/ijms22062894
Chicago/Turabian StyleCorinaldesi, Clarissa, Rebecca L. Ross, Giuseppina Abignano, Cristina Antinozzi, Francesco Marampon, Luigi di Luigi, Maya H. Buch, Valeria Riccieri, Andrea Lenzi, Clara Crescioli, and et al. 2021. "Muscle Damage in Systemic Sclerosis and CXCL10: The Potential Therapeutic Role of PDE5 Inhibition" International Journal of Molecular Sciences 22, no. 6: 2894. https://doi.org/10.3390/ijms22062894
APA StyleCorinaldesi, C., Ross, R. L., Abignano, G., Antinozzi, C., Marampon, F., di Luigi, L., Buch, M. H., Riccieri, V., Lenzi, A., Crescioli, C., & Del Galdo, F. (2021). Muscle Damage in Systemic Sclerosis and CXCL10: The Potential Therapeutic Role of PDE5 Inhibition. International Journal of Molecular Sciences, 22(6), 2894. https://doi.org/10.3390/ijms22062894