
Antenna Protein Clustering In Vitro Unveiled by Fluorescence Correlation Spectroscopy

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of pH and Detergent on LHCII Fluorescence Quenching: Bulk Measurements
2.2. FCS Unveiled Sizes of LHCII Clusters
2.3. Low pH Induced LHCII Large Particle Clusters
2.4. MEMFCS Analysis Highlighted LHCII Cluster Size Heterogeneity at Low pH
2.5. Aggregates Out of Physiological Range Induced by In Vitro Fluorescence quenching
2.6. Assessment of the Red-Shifted Fluorescence in the Samples
3. Discussion
4. Materials and Methods
4.1. LHCII Purification
4.2. Bulk Fluorescence Kinetic Measurements
4.3. Fluorescence Correlation Spectroscopy
4.4. FCS Data Analysis—Fitting of Autocorrelation Functions
4.5. FCS Data Analysis—Characterization of the Oligomers by the Maximum Entropy Method Algorithm (MEMFCS)
4.6. FCS Data Analysis—Estimation of the Size of LHCII Particles
4.7. Standard Confocal Microscopy Imaging of LHCII Particles
4.8. Low-Temperature Fluorescence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Z.; Yan, H.; Wang, K.; Kuang, T.; Zhang, J.; Gui, L.; An, X.; Chang, W. Crystal structure of spinach major light-harvesting complex at 2.72 A resolution. Nature 2004, 428, 287–292. [Google Scholar] [CrossRef]
- Standfuss, R.; van Scheltinga, A.C.T.; Lamborghini, M.; Kuhlbrandt, W. Mechanisms of photoprotection and nonphotochemical quenching in pea light-harvesting complex at 2.5 A resolution. EMBO J. 2005, 24, 919–928. [Google Scholar] [CrossRef] [Green Version]
- Kana, R.; Vass, I. Thermoimaging as a tool for studying light-induced heating of leaves Correlation of heat dissipation with the efficiency of photosystem II photochemistry and non-photochemical quenching. Environ. Exp. Bot. 2008, 64, 90–96. [Google Scholar] [CrossRef]
- Holt, N.E.; Fleming, G.R.; Niyogi, K.K. Toward an understanding of the mechanism of nonphotochemical quenching in green plants. Biochemistry 2004, 43, 8281–8289. [Google Scholar] [CrossRef]
- Goss, R.; Lepetit, B. Biodiversity of NPQ. J. Plant Physiol. 2015, 172, 13–32. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Ruban, A.V.; Rees, D.; Pascal, A.A.; Noctor, G.; Young, A.J. Control of the light-harvesting function of chloroplast membranes by aggregation of the LHCII chlorophyll-protein complex. FEBS Lett. 1991, 292, 1–4. [Google Scholar] [PubMed] [Green Version]
- Horton, P.; Wentworth, M.; Ruban, A. Control of the light harvesting function of chloroplast membranes: The LHCII-aggregation model for non-photochemical quenching. FEBS Lett. 2005, 579, 4201–4206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.P.; Gilmore, A.M.; Caffarri, S.; Bassi, R.; Golan, T.; Kramer, D.; Niyogi, K.K. Regulation of photosynthetic light harvesting involves intrathylakoid lumen pH sensing by the PsbS protein. J. Biol. Chem. 2004, 279, 22866–22874. [Google Scholar] [CrossRef] [Green Version]
- Ahn, T.K.; Avenson, T.J.; Ballottari, M.; Cheng, Y.C.; Niyogi, K.K.; Bassi, R.; Fleming, G.R. Architecture of a charge-transfer state regulating light harvesting in a plant antenna protein. Science 2008, 320, 794–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belgio, E.; Duffy, C.D.; Ruban, A.V. Switching light harvesting complex II into photoprotective state involves the lumen-facing apoprotein loop. Phys. Chem. Chem. Phys. 2013, 15, 12253–12261. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V.; Horton, P. Mechanism of DeltapH-Dependent Dissipation of Absorbed Excitation Energy by Photosynthetic Membranes. 1. Spectroscopic Analysis of Isolated Light-Harvesting Complexes. Biochim. Biophys. Acta 1992, 1102, 30–38. [Google Scholar] [CrossRef]
- Ruban, A.V.; Rees, D.; Pascal, A.A.; Horton, P. Mechanism of DeltapH-Dependent Dissipation of Absorbed Excitation Energy by Photosynthetic Membranes. 2. The Relationship Between LHCII Aggregation Invitro and qE in Isolated Thylakoids. Biochim. Biophys. Acta 1992, 1102, 39–44. [Google Scholar] [CrossRef]
- Johnson, M.P.; Ruban, A.V. Restoration of rapidly reversible photoprotective energy dissipation in the absence of PsbS protein by enhanced DeltapH. J. Biol. Chem. 2011, 286, 19973–19981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuthanova Trskova, E.; Belgio, E.; Yeates, A.M.; Sobotka, R.; Ruban, A.V.; Kana, R. Antenna proton sensitivity determines photosynthetic light harvesting strategy. J. Exp. Bot. 2018, 69, 4483–4493. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Nawrocki, W.J.; Liu, X.; Polukhina, I.; van Stokkum, I.H.M.; Croce, R. pH dependence, kinetics and light-harvesting regulation of nonphotochemical quenching in Chlamydomonas. Proc Natl. Acad. Sci. USA 2019, 116, 8320–8325. [Google Scholar] [CrossRef] [Green Version]
- Siffel, P.; Vacha, F. Aggregation of the light-harvesting complex in intact leaves of tobacco plants stressed by CO2 deficit. Photochem. Photobiol. 1998, 67, 304–311. [Google Scholar] [CrossRef]
- Tang, Y.L.; Wen, X.G.; Lu, Q.T.; Yang, Z.P.; Cheng, Z.K.; Lu, C.M. Heat stress induces an aggregation of the light-harvesting complex of photosystem II in spinach plants. Plant Physiol. 2007, 143, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Miloslavina, Y.; Wehner, A.; Lambrev, P.H.; Wientjes, E.; Reus, M.; Garab, G.; Croce, R.; Holzwarth, A.R. Far-red fluorescence: A direct spectroscopic marker for LHCII oligomer formation in non-photochemical quenching. FEBS Lett. 2008, 582, 3625–3631. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.P.; Ruban, A.V. Photoprotective energy dissipation in higher plants involves alteration of the excited state energy of the emitting chlorophyll(s) in the light harvesting antenna II (LHCII). J. Biol. Chem. 2009, 284, 23592–23601. [Google Scholar] [CrossRef] [Green Version]
- Janik, E.; Bednarska, J.; Zubik, M.; Puzio, M.; Luchowski, R.; Grudzinski, W.; Mazur, R.; Garstka, M.; Maksymiec, W.; Kulik, A.; et al. Molecular architecture of plant thylakoids under physiological and light stress conditions: A study of lipid-light-harvesting complex II model membranes. Plant Cell 2013, 25, 2155–2170. [Google Scholar] [CrossRef] [Green Version]
- Natali, A.; Gruber, J.M.; Dietzel, L.; Stuart, M.C.; van Grondelle, R.; Croce, R. Light-harvesting Complexes (LHCs) Cluster Spontaneously in Membrane Environment Leading to Shortening of Their Excited State Lifetimes. J. Biol. Chem. 2016, 291, 16730–16739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruban, A.V.; Horton, P.; Robert, B. Resonance Raman spectroscopy of the photosystem II light- harvesting complex of green plants: A comparison of trimeric and aggregated. Biochemistry 1995, 34, 2333–2337. [Google Scholar] [CrossRef] [PubMed]
- Barzda, V.; Peterman, E.J.G.; van Grondelle, R.; Van Amerongen, H. The influence of aggregation on triplet formation in light- harvesting chlorophyll a/b pigment-protein complex II of green plants. Biochemistry 1998, 37, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Van Oort, B.; van Hoek, A.; Ruban, A.V.; van Amerongen, H. Aggregation of light-harvesting complex II leads to formation of efficient excitation energy traps in monomeric and trimeric complexes. FEBS Lett. 2007, 581, 3528–3532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrou, K.; Belgio, E.; Ruban, A.V. pH sensitivity of chlorophyll fluorescence quenching is determined by the detergent/protein ratio and the state of LHCII aggregation. Biochim. Biophys. Acta 2014, 1837, 1533–1539. [Google Scholar] [CrossRef] [Green Version]
- Schaller, S.; Richter, K.; Wilhelm, C.; Goss, R. Influence of pH, Mg2+, and lipid composition on the aggregation state of the diatom FCP in comparison to the LHCII of vascular plants. Photosynth. Res. 2014, 119, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Kana, R.; Kotabova, E.; Sobotka, R.; Prasil, O. Non-photochemical quenching in cryptophyte alga Rhodomonas salina is located in chlorophyll a/c antennae. PLoS ONE 2012, 7, e29700. [Google Scholar] [CrossRef]
- Lambrev, P.H.; Schmitt, F.J.; Kussin, S.; Schoengen, M.; Varkonyi, Z.; Eichler, H.J.; Garab, G.; Renger, G. Functional domain size in aggregates of light-harvesting complex II and thylakoid membranes. Biochim. Biophys. Acta 2011, 1807, 1022–1031. [Google Scholar] [CrossRef] [Green Version]
- Magdaong, N.M.; Enriquez, M.M.; LaFountain, A.M.; Rafka, L.; Frank, H.A. Effect of protein aggregation on the spectroscopic properties and excited state kinetics of the LHCII pigment-protein complex from green plants. Photosynth. Res. 2013, 118, 259–276. [Google Scholar] [CrossRef]
- Enriquez, M.M.; Akhtar, P.; Zhang, C.; Garab, G.; Lambrev, P.H.; Tan, H.S. Energy transfer dynamics in trimers and aggregates of light-harvesting complex II probed by 2D electronic spectroscopy. J. Chem. Phys. 2015, 142, 212432. [Google Scholar] [CrossRef] [Green Version]
- Kramer, D.M.; Avenson, T.J.; Edwards, G.E. Dynamic flexibility in the light reactions of photosynthesis governed by both electron and proton transfer reactions. Trends Plant Sci. 2004, 9, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Joliot, P.; Johnson, G.N. Regulation of cyclic and linear electron flow in higher plants. Proc. Natl. Acad. Sci. USA 2011, 108, 13317–13322. [Google Scholar] [CrossRef] [Green Version]
- Jarvi, S.; Gollan, P.J.; Aro, E.M. Understanding the roles of the thylakoid lumen in photosynthesis regulation. Front. Plant Sci. 2013, 4, 434. [Google Scholar] [CrossRef] [Green Version]
- Kramer, D.M.; Sacksteder, C.A.; Cruz, J.A. How acidic is the lumen? Photosynth. Res. 1999, 60, 151–163. [Google Scholar] [CrossRef]
- Kramer, D.M.; Cruz, J.A.; Kanazawa, A. Balancing the central roles of the thylakoid proton gradient. Trends Plant Sci. 2003, 8, 27–32. [Google Scholar] [CrossRef]
- Perozeni, F.; Cazzaniga, S.; Ballottari, M. In vitro and in vivo investigation of chlorophyll binding sites involved in non-photochemical quenching in Chlamydomonas reinhardtii. Plant Cell Environ. 2019, 42, 2522–2535. [Google Scholar] [CrossRef] [Green Version]
- Levenberg, K. A method for the solution of certain non-linear problems in least squares. Quart. Appl. Math. 1944, 2, 164–168. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, P.; Garai, K.; Balaji, J.; Periasamy, N.; Maiti, S. Measuring size distribution in highly heterogeneous systems with fluorescence correlation spectroscopy. Biophys. J. 2003, 84, 1977–1984. [Google Scholar] [CrossRef] [Green Version]
- Goins, A.B.; Sanabria, H.; Waxham, M.N. Macromolecular crowding and size effects on probe microviscosity. Biophys. J. 2008, 95, 5362–5373. [Google Scholar] [CrossRef] [Green Version]
- David, L.; Prado, M.; Arteni, A.A.; Elmlund, D.A.; Blankenship, R.E.; Adir, N. Structural studies show energy transfer within stabilized phycobilisomes independent of the mode of rod-core assembly. Biochim. Biophys. Acta 2014, 1837, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwai, M.; Pack, C.G.; Takenaka, Y.; Sako, Y.; Nakano, A. Photosystem II antenna phosphorylation-dependent protein diffusion determined by fluorescence correlation spectroscopy. Sci. Rep. 2013, 3, 2833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janik, E.; Bednarska, J.; Sowinski, K.; Luchowski, R.; Zubik, M.; Grudzinski, W.; Gruszecki, W.I. Light-induced formation of dimeric LHCII. Photosynth. Res. 2017, 132, 265–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kana, R.; Steinbach, G.; Sobotka, R.; Vamosi, G.; Komenda, J. Fast Diffusion of the Unassembled PetC1-GFP Protein in the Cyanobacterial Thylakoid Membrane. Life 2020, 11, 15. [Google Scholar] [CrossRef]
- Jennings, R.C.; Garlaschi, F.M.; Zucchelli, G. Light-induced fluorescence quenching in the light-harvesting chlorophyll a/b protein complex. Photosynth. Res. 1991, 27, 57–64. [Google Scholar] [CrossRef]
- Ostroumov, E.E.; Gotze, J.P.; Reus, M.; Lambrev, P.H.; Holzwarth, A.R. Characterization of fluorescent chlorophyll charge-transfer states as intermediates in the excited state quenching of light-harvesting complex II. Photosynth. Res. 2020, 144, 171–193. [Google Scholar] [CrossRef]
- Gruszecki, W.I.; Zubik, M.; Luchowski, R.; Grudzinski, W.; Gospodarek, M.; Szurkowski, J.; Gryczynski, Z.; Gryczynski, I. Investigation of the molecular mechanism of the blue-light-specific excitation energy quenching in the plant antenna complex LHCII. J. Plant Physiol. 2011, 168, 409–414. [Google Scholar] [CrossRef]
- Ruban, A.V.; Young, A.; Horton, P. Modulation of chlorophyll fluorescence quenching in isolated light harvesting complex of Photosystem II. Biochim. Biophys. Acta 1994, 1186, 123–127. [Google Scholar] [CrossRef]
- Schaller, S.; Latowski, D.; Jemiola-Rzeminska, M.; Dawood, A.; Wilhelm, C.; Strzalka, K.; Goss, R. Regulation of LHCII aggregation by different thylakoid membrane lipids. BBA Bioenerg. 2011, 1807, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, B.; Goswami, M.; Nag, S.; Maiti, S. Spontaneous formation of a protein corona prevents the loss of quantum dot fluorescence in physiological buffers. Chem. Phys. Lett. 2007, 445, 217–220. [Google Scholar] [CrossRef]
- Sahoo, B.; Nag, S.; Sengupta, P.; Maiti, S. On the Stability of the Soluble Amyloid Aggregates. Biophys. J. 2009, 97, 1454–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, N.; Verma, S.D.; Singh, M.K.; Sen, S. Fluorescence Correlation Spectroscopy: An Efficient Tool for Measuring Size, Size-Distribution and Polydispersity of Microemulsion Droplets in Solution. Anal. Chem. 2011, 83, 7736–7744. [Google Scholar] [CrossRef]
- Wentworth, M.; Ruban, A.V.; Horton, P. Kinetic analysis of nonphotochemical quenching of chlorophyll fluorescence. 2. Isolated light-harvesting complexes. Biochemistry 2001, 40, 9902–9908. [Google Scholar] [CrossRef] [PubMed]
- Danaei, M.; Dehghankhold, M.; Ataei, S.; Hasanzadeh Davarani, F.; Javanmard, R.; Dokhani, A.; Khorasani, S.; Mozafari, M.R. Impact of Particle Size and Polydispersity Index on the Clinical Applications of Lipidic Nanocarrier Systems. Pharmaceutics 2018, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- Gruszecki, W.I.; Grudzinski, W.; Gospodarek, M.; Patyra, M.; Maksymiec, W. Xanthophyll-induced aggregation of LHCII as a switch between light-harvesting and energy dissipation systems. Biochim. Biophys. Acta 2006, 1757, 1504–1511. [Google Scholar] [CrossRef] [Green Version]
- Barzda, V.; Vengris, M.; Valkunas, L.; van Grondelle, R.; Van Amerongen, H. Generation of fluorescence quenchers from the triplet states of chlorophylls in the major light-harvesting complex II from green plants. Biochemistry 2000, 39, 10468–10477. [Google Scholar] [CrossRef] [PubMed]
- Barzda, V.; de Grauw, C.J.; Gerritsen, H.C.; Kleima, F.J.; Van Amerongen, H.; van Grondelle, R.; Vroom, J. Fluorescence lifetime heterogeneity in aggregates of LHCII revealed by time-resolved microscopy. Biophys. J. 2001, 81, 538–546. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.X.; Chia, D.; Csizmok, V.; Farber, P.; Forman-Kay, J.D.; Gradinaru, C.C. The Effect of Intrachain Electrostatic Repulsion on Conformational Disorder and Dynamics of the Sic1 Protein. J. Phys. Chem. B 2014, 118, 4088–4097. [Google Scholar] [CrossRef]
- Vrandecic, K.; Ratsep, M.; Wilk, L.; Rusevich, L.; Golub, M.; Reppert, M.; Irrgang, K.D.; Kuhlbrandt, W.; Pieper, J. Protein dynamics tunes excited state positions in light-harvesting complex II. J. Phys. Chem. B 2015, 119, 3920–3930. [Google Scholar] [CrossRef]
- Kandula, H.N.; Jee, A.Y.; Granick, S. Robustness of FCS (Fluorescence Correlation Spectroscopy) with Quenchers Present. J. Phys. Chem. A 2019, 123, 10184–10189. [Google Scholar] [CrossRef] [Green Version]
- Dittrich, P.S.; Schwille, P. Photobleaching and stabilization of fluorophores used for single-molecule analysis with one- and two-photon excitation. Appl. Phys. B Lasers Opt. 2001, 73, 829–837. [Google Scholar] [CrossRef]
- Zhang, Z.; Yomo, D.; Gradinaru, C. Choosing the right fluorophore for single-molecule fluorescence studies in a lipid environment. BBA Biomembr. 2017, 1859, 1242–1253. [Google Scholar] [CrossRef]
- Gendron, P.O.; Avaltroni, F.; Wilkinson, K.J. Diffusion coefficients of several rhodamine derivatives as determined by pulsed field gradient-nuclear magnetic resonance and fluorescence correlation spectroscopy. J. Fluoresc. 2008, 18, 1093–1101. [Google Scholar] [CrossRef]
- Caffarri, S.; Tibiletti, T.; Jennings, R.C.; Santabarbara, S. A comparison between plant photosystem I and photosystem II architecture and functioning. Curr. Protein Pept. Sci. 2014, 15, 296–331. [Google Scholar] [CrossRef]
- Ruban, A.V.; Rees, D.; Noctor, G.D.; Young, A.; Horton, P. Long-wavelength chlorophyll species are associated with amplification of high-energy-state excitation quenching in higher plants. Biochim. Biophys. Acta 1991, 1059, 355–360. [Google Scholar] [CrossRef]
- Caffarri, S.; Kouril, R.; Kereiche, S.; Boekema, E.J.; Croce, R. Functional architecture of higher plant photosystem II supercomplexes. EMBO J. 2009, 28, 3052–3063. [Google Scholar] [CrossRef] [Green Version]
- Su, X.; Ma, J.; Wei, X.; Cao, P.; Zhu, D.; Chang, W.; Liu, Z.; Zhang, X.; Li, M. Structure and assembly mechanism of plant C2S2M2-type PSII-LHCII supercomplex. Science 2017, 357, 815–820. [Google Scholar] [CrossRef] [Green Version]
- Van Haeringen, B.; Dekker, J.P.; Bloemendal, M.; Rögner, M.; van Grondelle, R.; Van Amerongen, H. Simultaneous measurement of electric birefringence and dichroism. A study on photosystem 1 particles. Biophys. J. 1994, 67, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Dekker, J.P.; van Roon, H.; Boekem, E.J. Heptameric association of light-harvesting complex II trimers in partially solubilized photosystem II membranes. FEBS Lett. 1999, 449, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Ruban, A.V. Light harvesting control in plants. FEBS Lett. 2018, 592, 3030–3039. [Google Scholar] [CrossRef]
- Ruban, A.V.; Johnson, M.P.; Duffy, C.D. The photoprotective molecular switch in the photosystem II antenna. Biochim. Biophys. Acta 2012, 1817, 167–181. [Google Scholar] [CrossRef] [Green Version]
- Belgio, E.; Johnson, M.P.; Juric, S.; Ruban, A.V. Higher plant photosystem II light-harvesting antenna, not the reaction center, determines the excited-state lifetime-both the maximum and the nonphotochemically quenched. Biophys. J. 2012, 102, 2761–2771. [Google Scholar] [CrossRef] [Green Version]
- Ware, M.A.; Giovagnetti, V.; Belgio, E.; Ruban, A.V. PsbS protein modulates non-photochemical chlorophyll fluorescence quenching in membranes depleted of photosystems. J. Photoch. Photobio. B 2015, 152, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Zaks, J.; Amarnath, K.; Sylak-Glassman, E.J.; Fleming, G.R. Models and measurements of energy-dependent quenching. Photosynth. Res. 2013, 116, 389–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daum, B.; Nicastro, D.; Il, J.A.; McIntosh, J.R.; Kuhlbrandt, W. Arrangement of Photosystem II and ATP Synthase in Chloroplast Membranes of Spinach and Pea. Plant Cell 2010, 22, 1299–1312. [Google Scholar] [CrossRef] [Green Version]
- Kirchhoff, H. Chloroplast ultrastructure in plants. New Phytol. 2019, 223, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Wood, W.H.J.; Barnett, S.F.H.; Flannery, S.; Hunter, C.N.; Johnson, M.P. Dynamic Thylakoid Stacking Is Regulated by LHCII Phosphorylation but Not Its interaction with PSI. Plant Physiol. 2019, 180, 2152–2166. [Google Scholar] [CrossRef] [Green Version]
- Belgio, E.; Ungerer, P.; Ruban, A.V. Light-harvesting superstructures of green plant chloroplasts lacking photosystems. Plant Cell Environ. 2015, 38, 2035–2047. [Google Scholar] [CrossRef]
- Tietz, S.; Leuenberger, M.; Hohner, R.; Olson, A.H.; Fleming, G.R.; Kirchhoff, H. A proteoliposome-based system reveals how lipids control photosynthetic light harvesting. J. Biol. Chem. 2020, 295, 1857–1866. [Google Scholar] [CrossRef] [Green Version]
- Chmeliov, J.; Gelzinis, A.; Songaila, E.; Augulis, R.; Duffy, C.D.; Ruban, A.V.; Valkunas, L. The nature of self-regulation in photosynthetic light-harvesting antenna. Nat. Plants 2016, 2, 16045. [Google Scholar] [CrossRef]
- Ruban, A.V.; Horton, P. The xanthophyll cycle modulates the kinetics of nonphotochemical energy dissipation in isolated light-harvesting complexes, intact chloroplasts, and leaves of spinach. Plant Physiol. 1999, 119, 531–542. [Google Scholar] [CrossRef] [Green Version]
- Wentworth, M.; Ruban, A.V.; Horton, P. Chlorophyll fluorescence quenching in isolated light harvesting complexes induced by zeaxanthin. FEBS Lett. 2000, 471, 71–74. [Google Scholar] [CrossRef] [Green Version]
- Banks, D.S.; Tressler, C.; Peters, R.D.; Hofling, F.; Fradin, C. Characterizing anomalous diffusion in crowded polymer solutions and gels over five decades in time with variable-lengthscale fluorescence correlation spectroscopy. Soft Matter. 2016, 12, 4190–4203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieger, J.W.; Langowski, J. QuickFit 3.0: A Data Evaluation Application for Biophysics. 2015. Available online: http://www.dkfz.de/Macromol/quickfit/ (accessed on 7 January 2019).
- Dertinger, T.; Pacheco, V.; von der Hocht, I.; Hartmann, R.; Gregor, I.; Enderlein, J. Two-focus fluorescence correlation spectroscopy: A new tool for accurate and absolute diffusion measurements. Chem. Phys. Chem. 2007, 8, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Belgio, E.; Kapitonova, E.; Chmeliov, J.; Duffy, C.D.P.; Ungerer, P.; Valkunas, L.; Ruban, A.V. Economic photoprotection in photosystem II that retains a complete light-harvesting system with slow energy traps. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
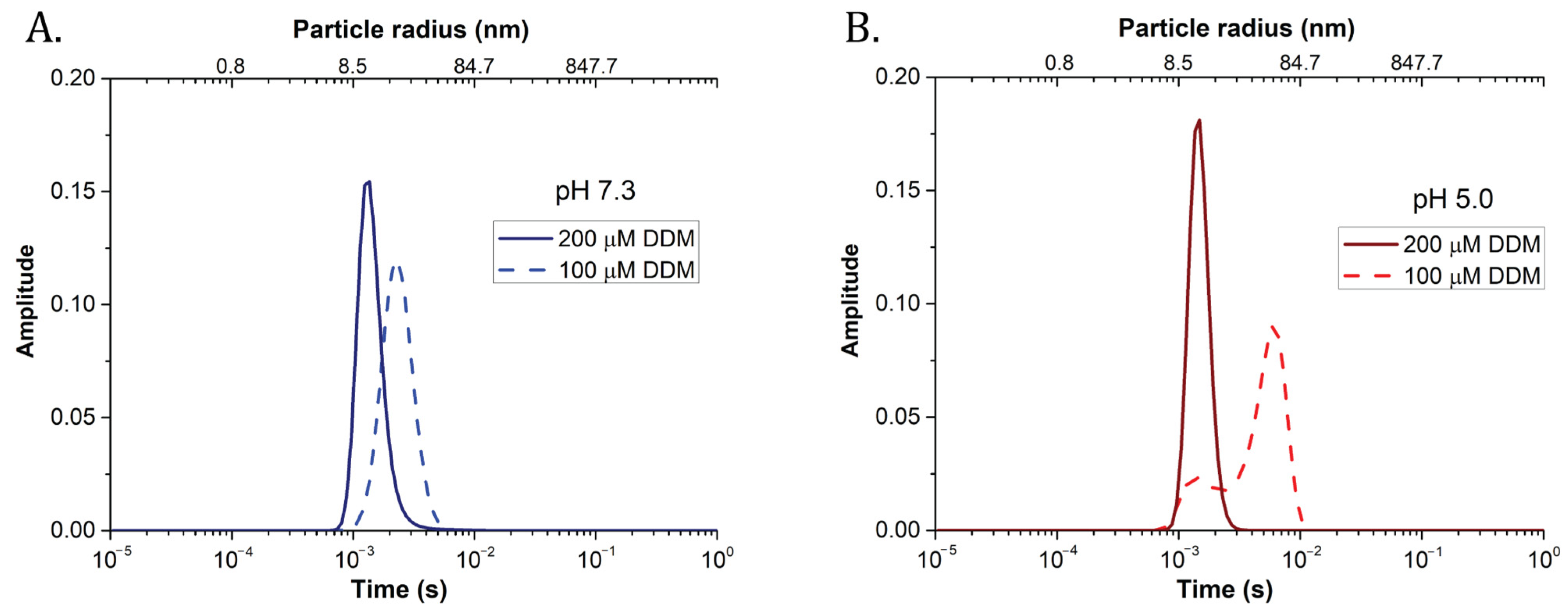{kind=link}
{kind=link}
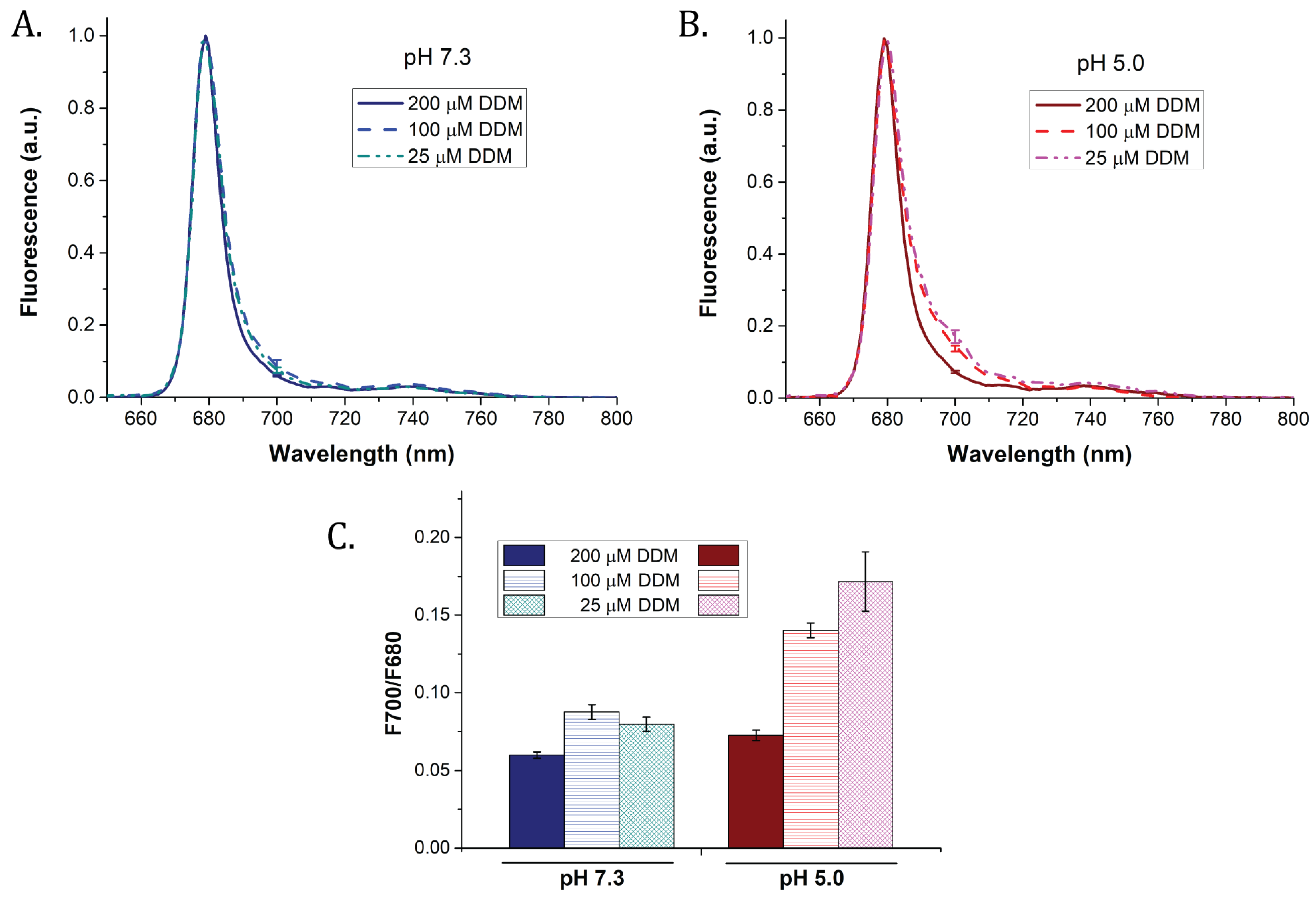{kind=link}
| pH | DDM Concentration | FQ | KD |
|---|---|---|---|
| 7.3 | 200 µM | 12.4 ± 1.2% | 0.1 |
| 100 µM | 43.7 ± 0.5% | 0.7 | |
| 25 µM | 61.7 ± 1.7% | 1.4 | |
| 5.0 | 200 µM | 20.1 ± 3.5% | 0.2 |
| 100 µM | 70.9 ± 1.2% | 2 | |
| 25 µM | 75.6 ± 0.7% | 3 |
| DDM Concentration | pH | Diffusion Time (ms) | Radius (nm) | Total Count Rate (KHz) | N |
|---|---|---|---|---|---|
| 200 μM | pH 7.3 | 1.18 ± 0.10 | 9.2 ± 0.8 | 11.48 ± 0.21 | 35.0 ± 0.7 |
| pH 5.0 | 1.10 ± 0.09 | 7.9 ± 0.0 | 11.67 ± 0.17 | 36.3 ± 3.2 | |
| 100 μM | pH 7.3 | 1.83 ± 0.18 | 13.4 ± 1.4 | 8.16 ± 0.19 | 23.5 ± 3.0 |
| pH 5.0 | 3.56 ± 0.53 | 30.1 ± 1.5 | 4.11 ± 0.05 | 4.7 ± 1.2 | |
| 25 μM | pH 7.3 | 1.80 ± 0.28 | 15.4 ± 2.4 | 4.69 ± 0.32 | 18.3 ± 2.1 |
| pH 5.0 | 7.60 ± 3.05 | 65.8 ± 27.0 | 1.56 ± 0.14 | 0.5 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crepin, A.; Cunill-Semanat, E.; Kuthanová Trsková, E.; Belgio, E.; Kaňa, R. Antenna Protein Clustering In Vitro Unveiled by Fluorescence Correlation Spectroscopy. Int. J. Mol. Sci. 2021, 22, 2969. https://doi.org/10.3390/ijms22062969
Crepin A, Cunill-Semanat E, Kuthanová Trsková E, Belgio E, Kaňa R. Antenna Protein Clustering In Vitro Unveiled by Fluorescence Correlation Spectroscopy. International Journal of Molecular Sciences. 2021; 22(6):2969. https://doi.org/10.3390/ijms22062969
Chicago/Turabian StyleCrepin, Aurélie, Edel Cunill-Semanat, Eliška Kuthanová Trsková, Erica Belgio, and Radek Kaňa. 2021. "Antenna Protein Clustering In Vitro Unveiled by Fluorescence Correlation Spectroscopy" International Journal of Molecular Sciences 22, no. 6: 2969. https://doi.org/10.3390/ijms22062969
APA StyleCrepin, A., Cunill-Semanat, E., Kuthanová Trsková, E., Belgio, E., & Kaňa, R. (2021). Antenna Protein Clustering In Vitro Unveiled by Fluorescence Correlation Spectroscopy. International Journal of Molecular Sciences, 22(6), 2969. https://doi.org/10.3390/ijms22062969