
Cold Atmospheric Plasma Modification of Amyloid β

 , , , , , and
, , , , , and 
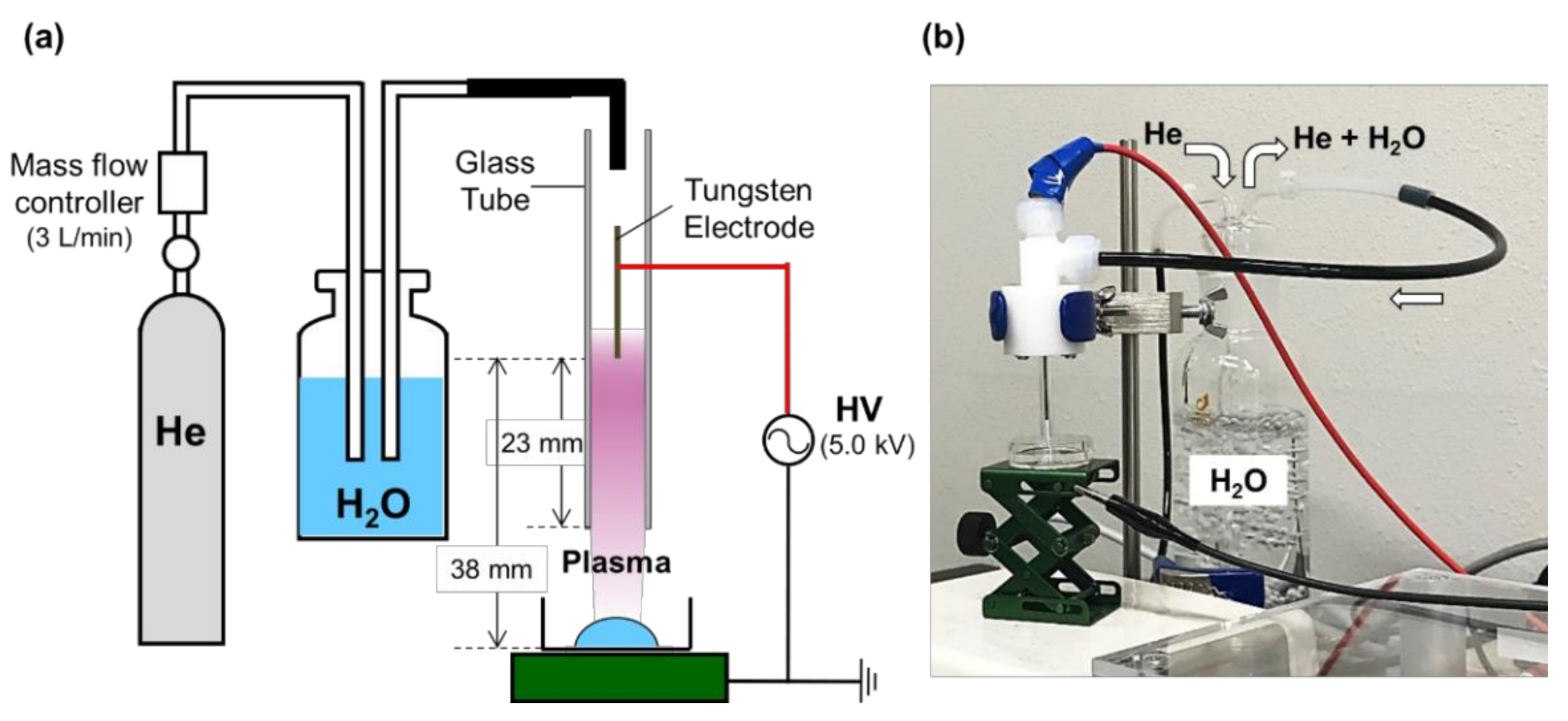{kind=link}
{kind=link}
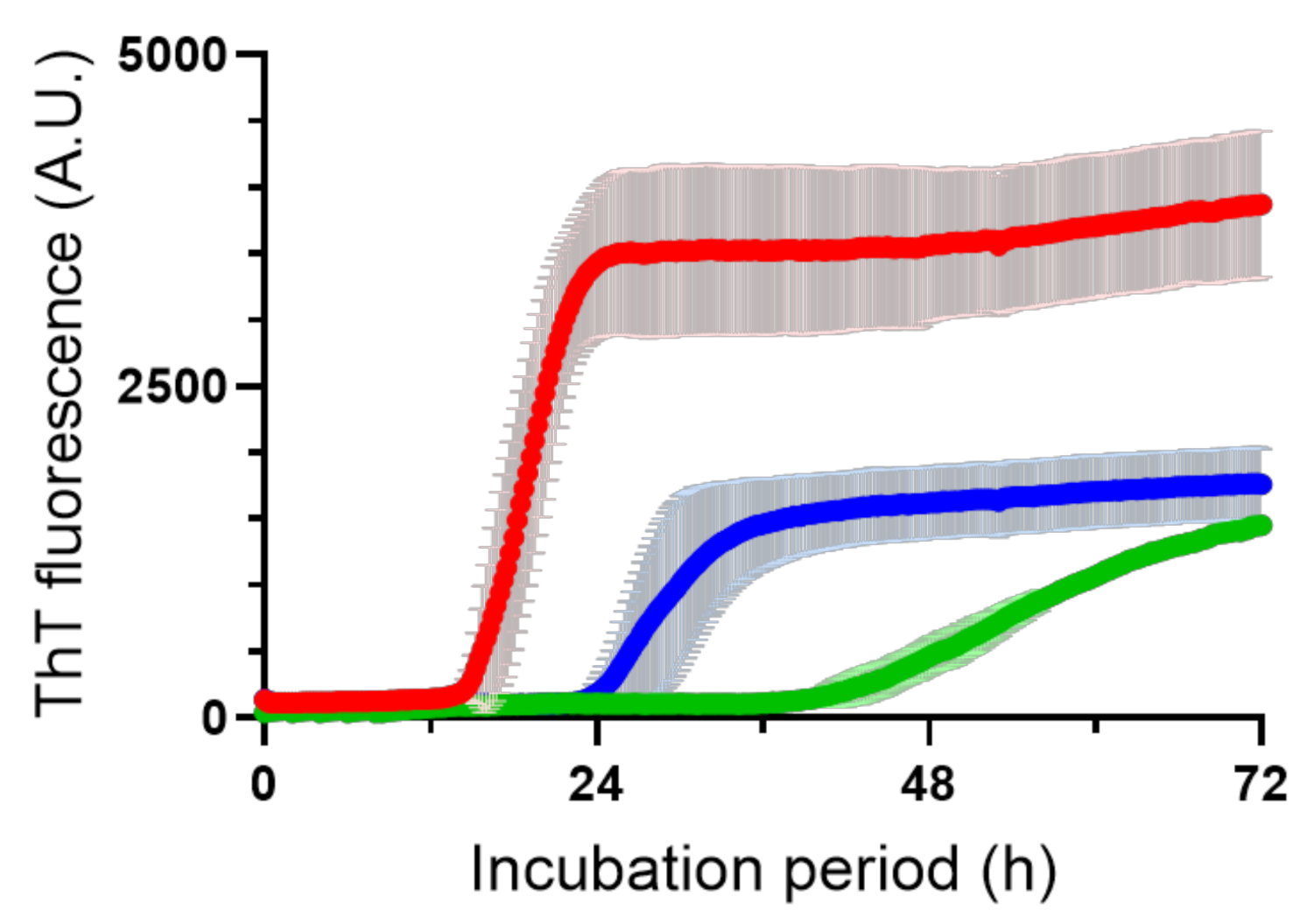{kind=link}
{kind=link}
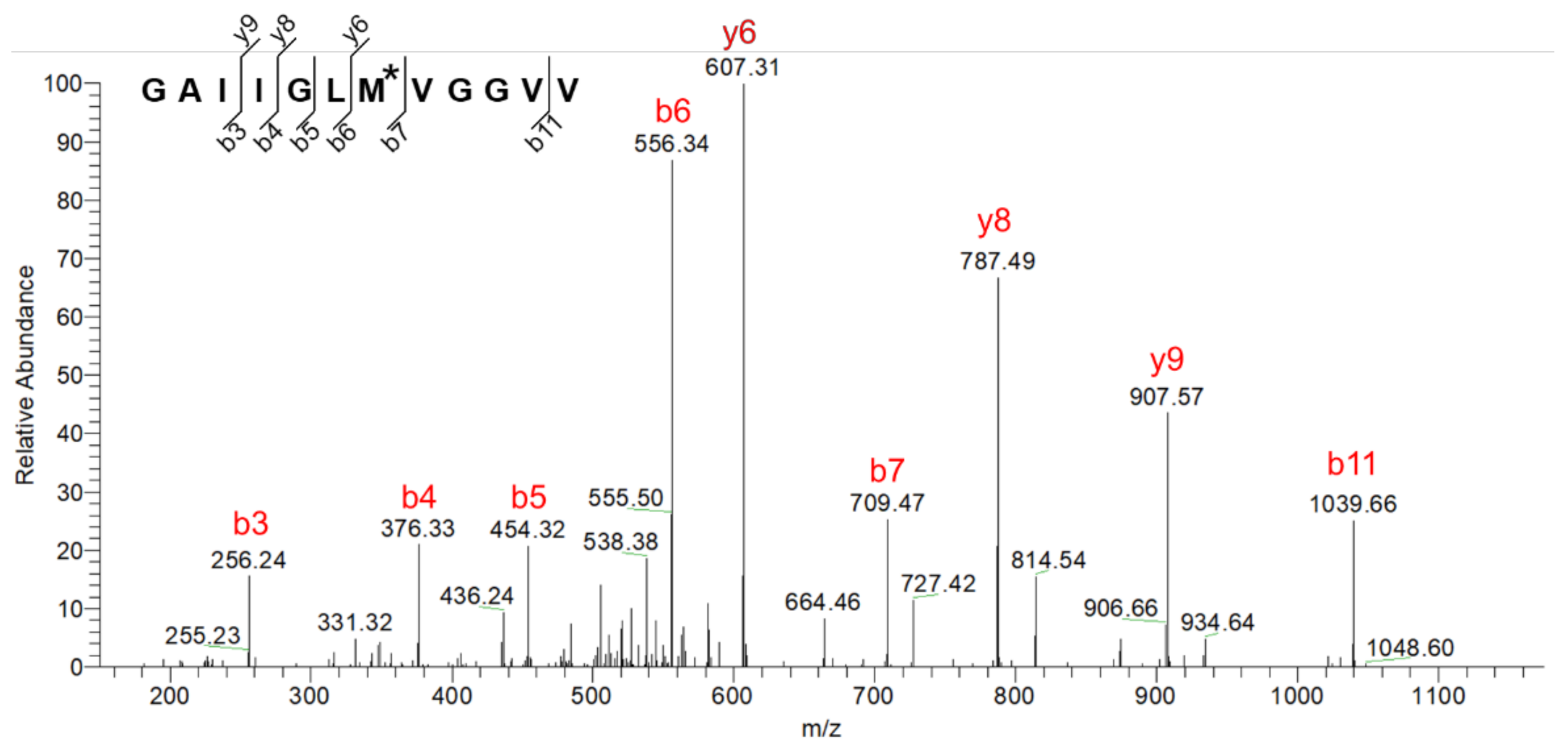{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. NMR Characterization of CAP-Treated Aβ
2.2. CAP Effects on Aβ Fibril Formation
2.3. Characterization of Chemical Modification of Aβ Caused by CAP Treatment
2.4. Reactive Species Responsible for Methionine Oxidation of Aβ
3. Materials and Methods
3.1. Aβ Sample Preparation
3.2. CAP Irradiation
3.3. NMR Experiments
3.4. HPLC Analyses
3.5. MS Analyses
3.6. ThT Assay
3.7. TEM
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bernhardt, T.; Semmler, M.L.; Schäfer, M.; Bekeschus, S.; Emmert, S.; Boeckmann, L. Plasma Medicine: Applications of Cold Atmospheric Pressure Plasma in Dermatology. Oxidative Med. Cell. Longev. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Braný, D.; Dvorská, D.; Halašová, E.; Škovierová, H. Cold Atmospheric Plasma: A Powerful Tool for Modern Medicine. Int. J. Mol. Sci. 2020, 21, 2932. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Wang, G.; Ruan, Q.; Tang, K.; Chu, P.K. Plasma-activated interfaces for biomedical engineering. Bioact. Mater. 2021, 6, 2134–2143. [Google Scholar] [CrossRef] [PubMed]
- Attri, P.; Kumar, N.; Park, J.H.; Yadav, D.K.; Choi, S.; Uhm, H.S.; Kim, I.T.; Choi, E.H.; Lee, W. Influence of reactive species on the modification of biomolecules generated from the soft plasma. Sci. Rep. 2015, 5, srep08221. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Attri, P.; Lee, I.; Oh, J.; Yun, J.-H.; Park, J.H.; Choi, E.H.; Lee, W. Structural and functional analysis of lysozyme after treatment with dielectric barrier discharge plasma and atmospheric pressure plasma jet. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Attri, P.; Han, J.; Choi, S.; Choi, E.H.; Bogaerts, A.; Lee, W. CAP modifies the structure of a model protein from thermophilic bacteria: Mechanisms of CAP-mediated inactivation. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attri, P.; Park, J.-H.; De Backer, J.; Kim, M.; Yun, J.-H.; Heo, Y.; Dewilde, S.; Shiratani, M.; Choi, E.H.; Lee, W.; et al. Structural modification of NADPH oxidase activator (Noxa 1) by oxidative stress: An experimental and computational study. Int. J. Biol. Macromol. 2020, 163, 2405–2414. [Google Scholar] [CrossRef] [PubMed]
- Takai, E.; Kitamura, T.; Kuwabara, J.; Ikawa, S.; Yoshizawa, S.; Shiraki, K.; Kawasaki, H.; Arakawa, R.; Kitano, K. Chemical modification of amino acids by atmospheric-pressure cold plasma in aqueous solution. J. Phys. D Appl. Phys. 2014, 47, 285403. [Google Scholar] [CrossRef]
- Bayliss, D.L.; Walsh, J.L.; Shama, G.; Iza, F.; Kong, M.G. Reduction and degradation of amyloid aggregates by a pulsed radio-frequency cold atmospheric plasma jet. New J. Phys. 2009, 11, 115024. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Sachse, C.; Richter, W.; Xu, C.; Fandrich, M.; Grigorieff, N. Comparison of Alzheimer Aβ (1–40) and Aβ (1–42) amyloid fibrils reveals similar protofilament structures. Proc. Natl. Acad. Sci. USA 2009, 106, 19813–19818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi-Utsumi, M.; Matsuo, K.; Yanagisawa, K.; Gekko, K.; Kato, K. Spectroscopic Characterization of Intermolecular Interaction of Amyloid β Promoted on GM1 Micelles. Int. J. Alzheimer’s Dis. 2010, 2011, 925073. [Google Scholar]
- Matsuzaki, K.; Kato, K.; Yanagisawa, K. Aβ polymerization through interaction with membrane gangliosides. Biochim. Biophys. Acta 2010, 1801, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Tycko, R.; Wickner, R.B. Molecular Structures of Amyloid and Prion Fibrils: Consensus versus Controversy. Acc. Chem. Res. 2013, 46, 1487–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.; Rohou, A.; Lasker, K.; Yadav, J.K.; Schiene-Fischer, C.; Fandrich, M.; Grigorieff, N. Peptide dimer structure in an Aβ(1-42) fibril visualized with cryo-EM. Proc. Natl. Acad. Sci. USA 2015, 112, 11858–11863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe-Nakayama, T.; Ono, K.; Itami, M.; Takahashi, R.; Teplow, D.B.; Yamada, M. High-speed atomic force microscopy reveals structural dynamics of amyloid β1-42 aggregates. Proc. Natl. Acad. Sci. USA 2016, 113, 5835–5840. [Google Scholar] [CrossRef] [Green Version]
- Meier, B.H.; Riek, R.; Böckmann, A. Emerging Structural Understanding of Amyloid Fibrils by Solid-State NMR. Trends Biochem. Sci. 2017, 42, 777–787. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, K.; Kato, K.; Yanagisawa, K. Ganglioside-Mediated Assembly of Amyloid β-Protein: Roles in Alzheimer’s Disease. Prog. Mol. Biol. Transl. Sci. 2018, 156, 413–434. [Google Scholar]
- Yagi-Utsumi, M. NMR Characterization of Conformational Dynamics and Molecular Assemblies of Proteins. Biol. Pharm. Bull. 2019, 42, 867–872. [Google Scholar] [CrossRef] [Green Version]
- Itoh, S.G.; Yagi-Utsumi, M.; Kato, K.; Okumura, H. Effects of a Hydrophilic/Hydrophobic Interface on Amyloid-β Peptides Studied by Molecular Dynamics Simulations and NMR Experiments. J. Phys. Chem. B 2019, 123, 160–169. [Google Scholar] [CrossRef]
- Yagi-Utsumi, M.; Yanaka, S.; Song, C.; Satoh, T.; Yamazaki, C.; Kasahara, H.; Shimazu, T.; Murata, K.; Kato, K. Characterization of amyloid β fibril formation under microgravity conditions. NPJ Microgravity 2020, 6, 17. [Google Scholar] [CrossRef]
- Hiller, K.-O.; Masloch, B.; Gobl, M.; Asmus, K.-D. Mechanism of the OH• Radical Induced Oxidation of Methionine in Aqueous Solution. J. Am. Chem. Soc. 1981, 103, 2734–2743. [Google Scholar] [CrossRef]
- Anzai, K.; Aoki, T.; Koshimizu, S.; Takaya, R.; Tsuchida, K.; Takajo, T. Formation of reactive oxygen species by irradiation of cold atmospheric pressure plasma jet to water depends on the irradiation distance. J. Clin. Biochem. Nutr. 2019, 64, 187–193. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Shao, H.; Zhang, Y.; Li, H.; Menon, N.K.; Neuhaus, E.B.; Brewer, J.M.; Byeon, I.J.; Ray, D.G.; Vitek, M.P.; et al. Solution NMR studies of the Ab(1–40) and Ab(1–42) peptides establish that the Met35 oxidation state affects the mechanism of amyloid formation. J. Am. Chem. Soc. 2004, 126, 1992–2005. [Google Scholar] [CrossRef]
- Gu, M.; Viles, J.H. Methionine oxidation reduces lag-times for amyloid-b(1–40) fiber formation but generates highly fragmented fibers. Biochim. Biophys. Acta 2016, 1864, 1260–1269. [Google Scholar] [CrossRef]
- Razzokov, J.; Yusupov, M.; Bogaerts, A. Oxidation destabilizes toxic amyloid β peptide aggregation. Sci. Rep. 2019, 9, 1–7. [Google Scholar] [CrossRef]
- Sasaki, S.; Kanzaki, M.; Kaneko, T. Calcium influx through TRP channels induced by short-lived reactive species in plasma-irradiated solution. Sci. Rep. 2016, 6, 25728. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, S.; Hokari, Y.; Kumada, A.; Kanzaki, M.; Kaneko, T. Direct plasma stimuli including electrostimulation and OH radical induce transient increase in intracellular Ca2+and uptake of a middle-size membrane-impermeable molecule. Plasma Process. Polym. 2018, 15, 1700077. [Google Scholar] [CrossRef]
- Yoshimura, S.; Aramaki, M.; Otsubo, Y.; Yamashita, A.; Koga, K. Controlling feeding gas temperature of plasma jet with Peltier device for experiments with fission yeast. Jpn. J. Appl. Phys. 2019, 58, SEEG03. [Google Scholar] [CrossRef]
- Palmblad, M.; Westlind-Danielsson, A.; Bergquist, J. Oxidation of methionine 35 attenuates formation of amyloid β-peptide 1–40 oligomers. J. Biol. Chem. 2002, 277, 19506–19510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, A.S.; Bergquist, J.; Volbracht, C.; Paivio, A.; Leist, M.; Lannfelt, L.; Westlind-Danielsson, A. Attenuated amyloid-β aggregation and neurotoxicity owing to methionine oxidation. Neuroreport 2007, 18, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carelli-Alinovi, C.; Misiti, F. Methionine 35 sulphoxide reduces toxicity of Aβ in red blood cell. Eur. J. Clin. Investig. 2017, 47, 314–321. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yagi-Utsumi, M.; Tanaka, T.; Otsubo, Y.; Yamashita, A.; Yoshimura, S.; Nishida, M.; Kato, K. Cold Atmospheric Plasma Modification of Amyloid β. Int. J. Mol. Sci. 2021, 22, 3116. https://doi.org/10.3390/ijms22063116
Yagi-Utsumi M, Tanaka T, Otsubo Y, Yamashita A, Yoshimura S, Nishida M, Kato K. Cold Atmospheric Plasma Modification of Amyloid β. International Journal of Molecular Sciences. 2021; 22(6):3116. https://doi.org/10.3390/ijms22063116
Chicago/Turabian StyleYagi-Utsumi, Maho, Tomohiro Tanaka, Yoko Otsubo, Akira Yamashita, Shinji Yoshimura, Motohiro Nishida, and Koichi Kato. 2021. "Cold Atmospheric Plasma Modification of Amyloid β" International Journal of Molecular Sciences 22, no. 6: 3116. https://doi.org/10.3390/ijms22063116
APA StyleYagi-Utsumi, M., Tanaka, T., Otsubo, Y., Yamashita, A., Yoshimura, S., Nishida, M., & Kato, K. (2021). Cold Atmospheric Plasma Modification of Amyloid β. International Journal of Molecular Sciences, 22(6), 3116. https://doi.org/10.3390/ijms22063116