
Is the Membrane Lipid Matrix a Key Target for Action of Pharmacologically Active Plant Saponins?


Abstract
:1. Introduction
2. Results and Discussion
2.1. The Effect of Saponins and Related Compounds on the Ion Permeability of Lipid Bilayers
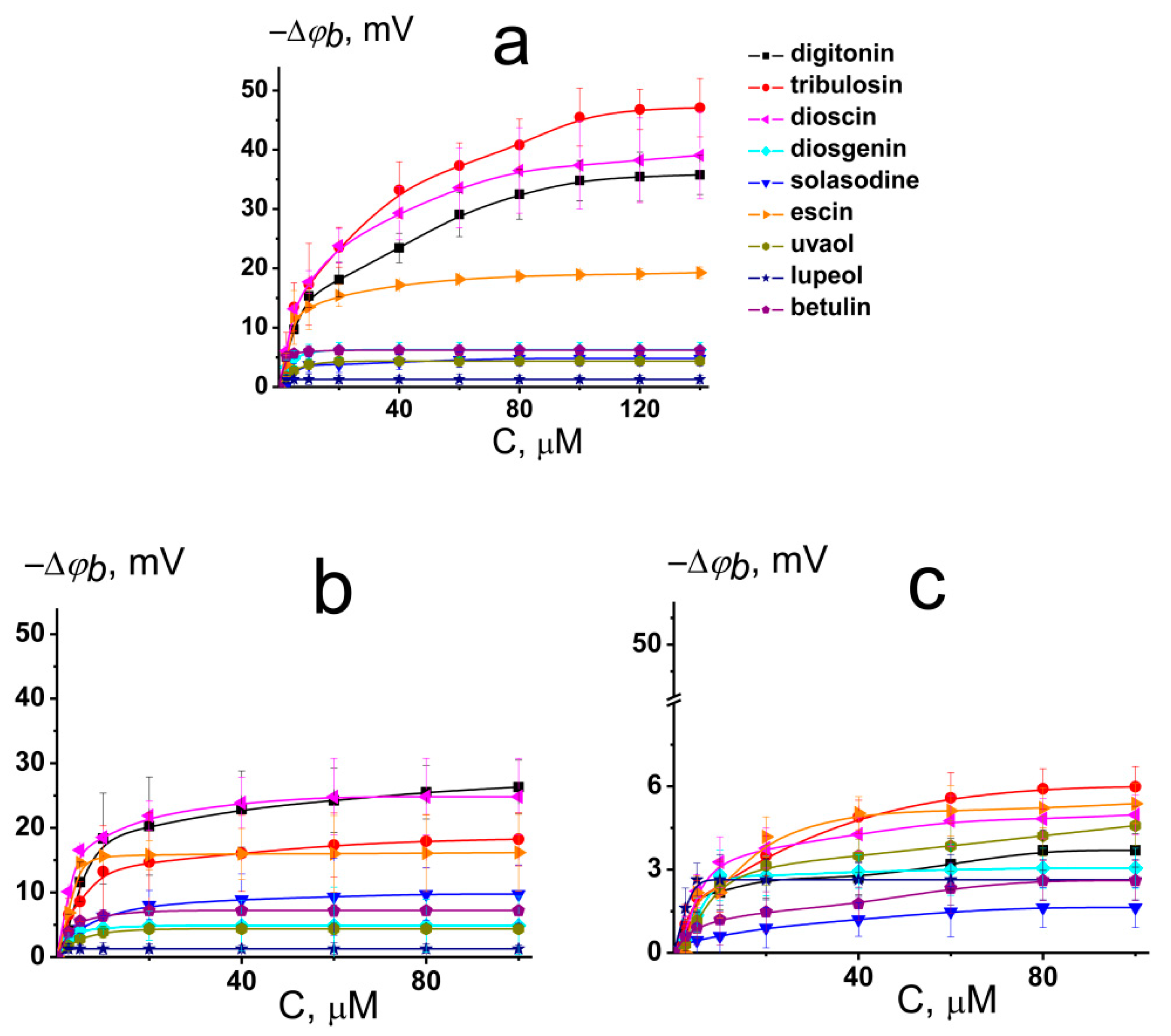
2.2. The Influence of Saponins and Related Compounds on the Boundary Potential of Model Lipid Membranes
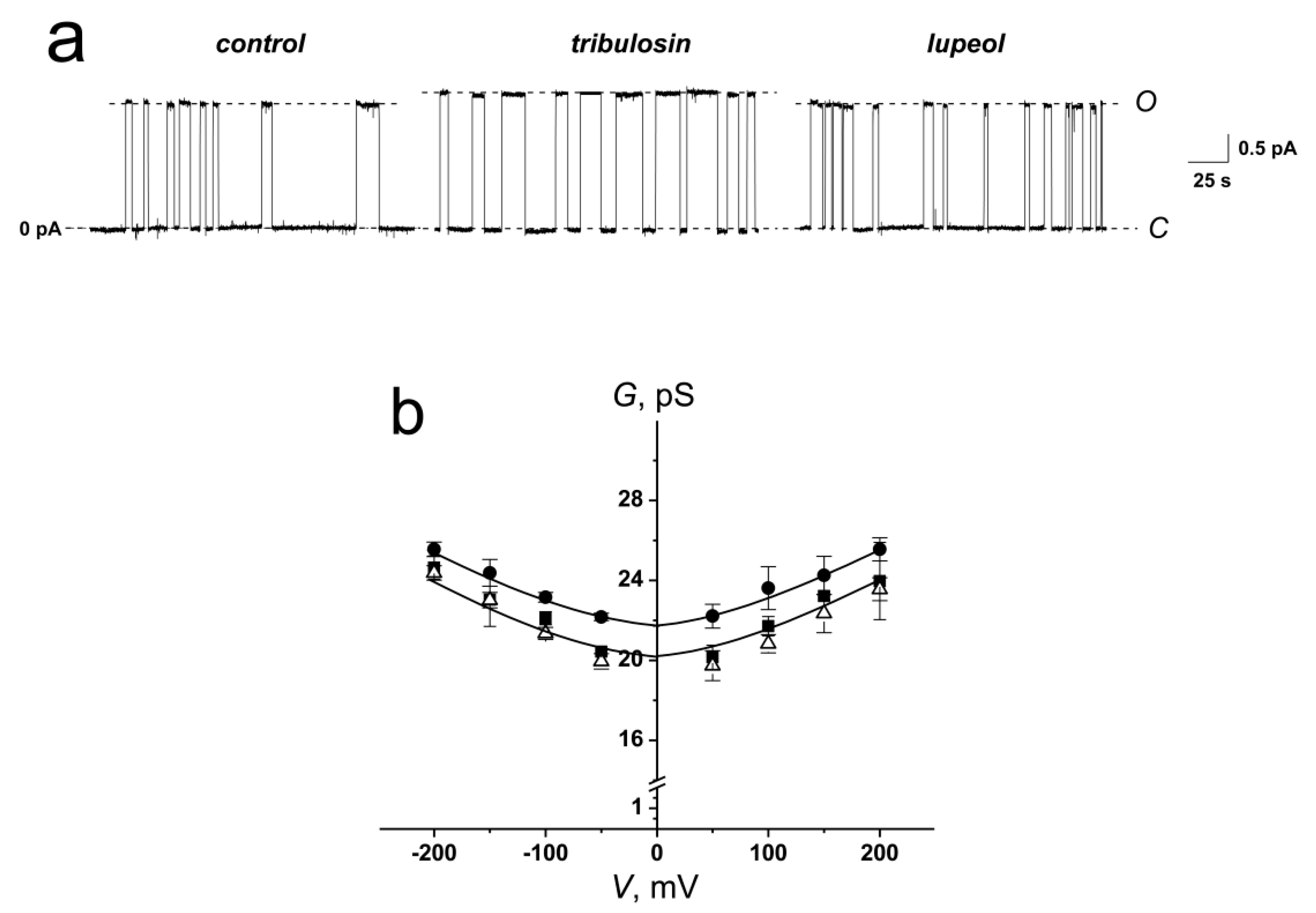
2.3. The Effects of Saponins and Related Compounds on the Voltage-Sensitive ion Channels Formed by Antimicrobial Peptide
2.4. The Effects of Saponins and Related Compounds on Lipid Melting
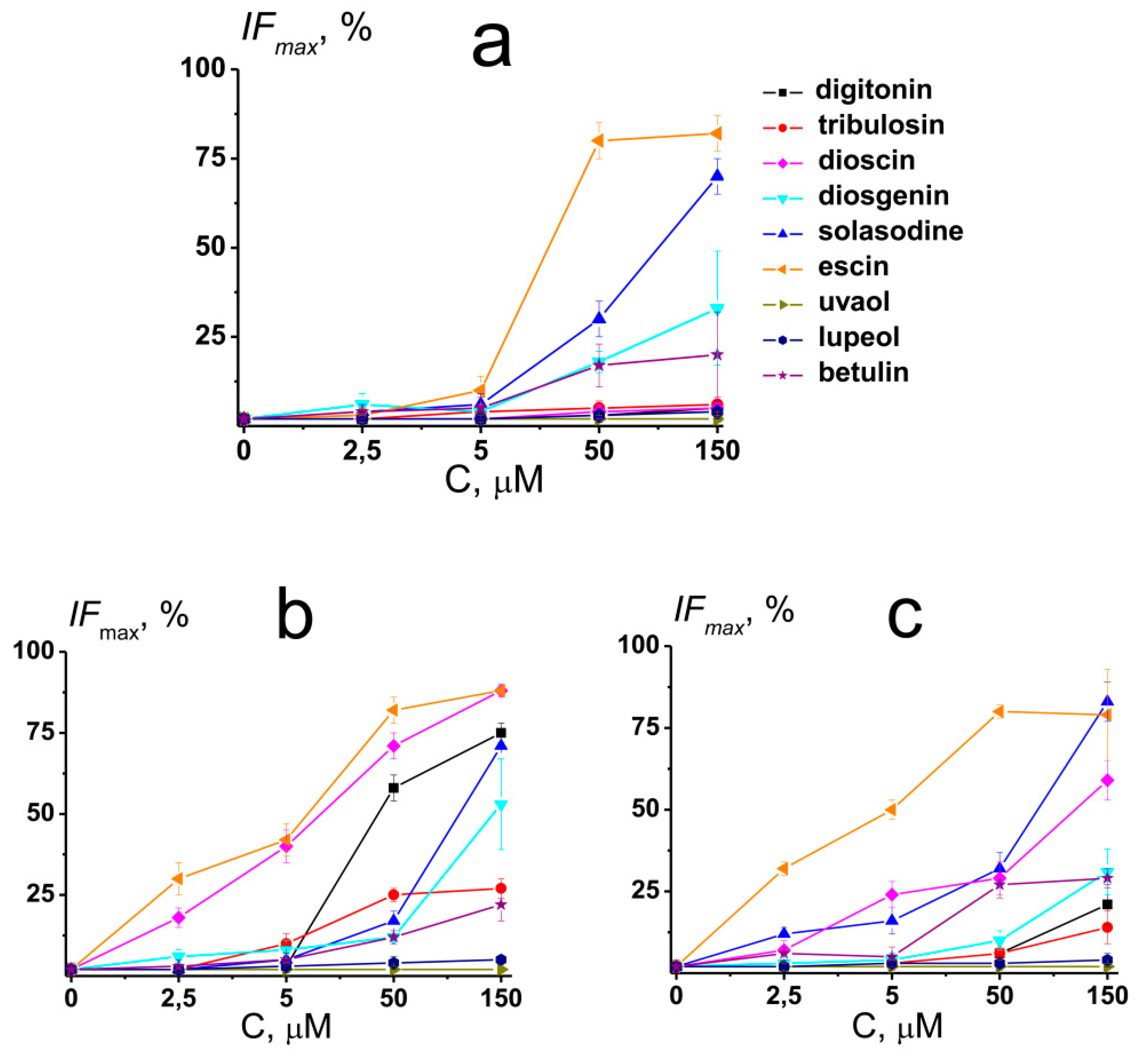
2.5. The Effect of Saponins and Related Compounds on the Calcein Leakage
2.6. The Influence of Saponins and Related Compounds on the Mechanosensitive ion Channels Induced by Various Antifungals
3. Materials and Methods
3.1. Chemical Reagents
3.2. Electrophysiological Method for Measuring the Membrane Boundary Potential
3.3. Reconstitution of Ion Channels into Lipid Bilayers
3.4. Calcein Release Assay
3.5. Differential Scanning Microcalorimetry of Liposomal Suspensions
3.6. Correlation Analysis between the Altered Membrane Parameter and Octanol/Water Partition Coefficient of Tested Compounds
4. Conclusions
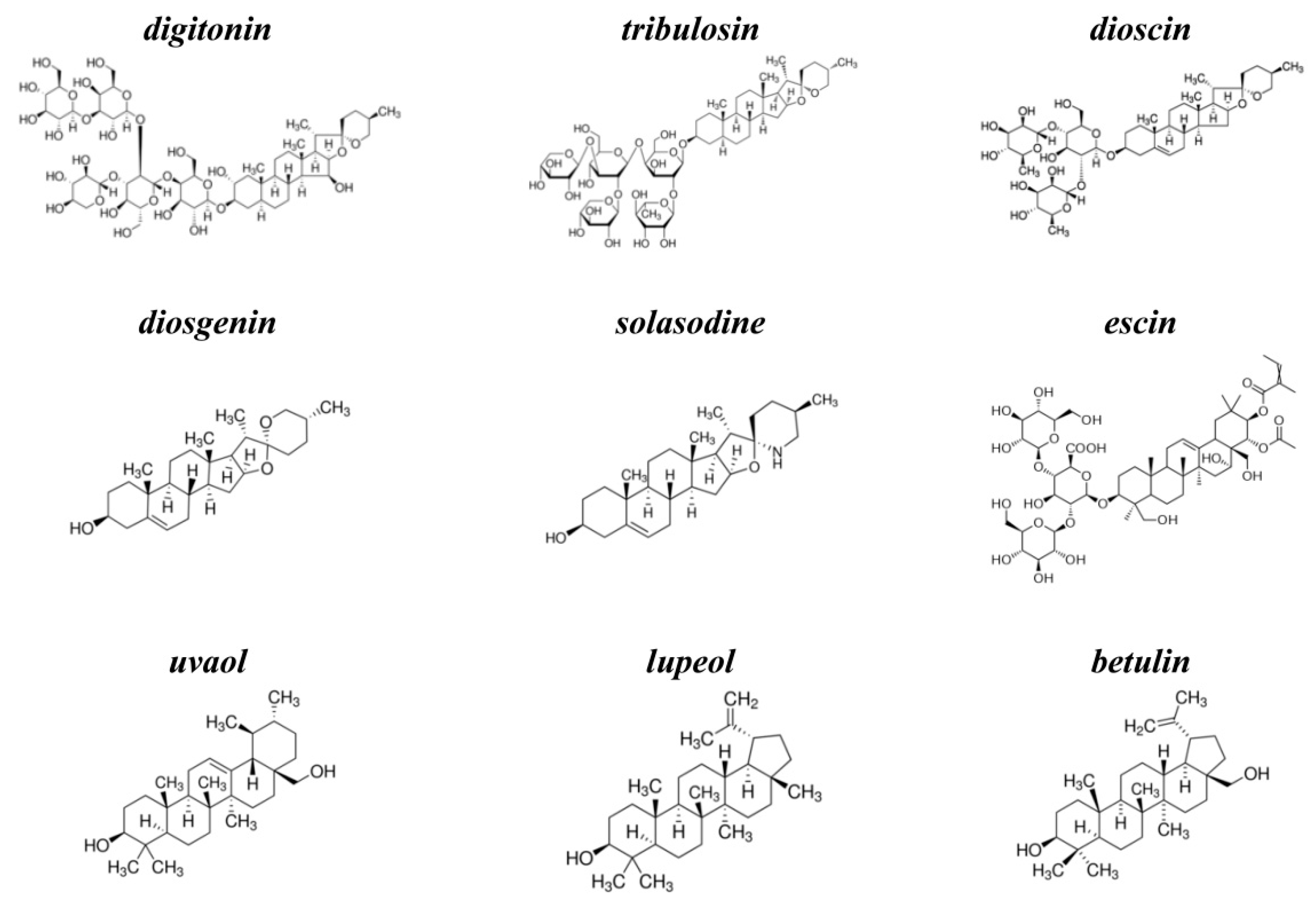
- The ability of saponins and related compounds to induce transmembrane pores depends on the agent type and membrane lipid composition. Cholesterol enhances the pore-forming and detergent-like activity of the compounds. All tested agents of steroid nature (digitonin, tribulosin, dioscin, diosgenin, and solasodine), and only one triterpenic sapogenin (betulin) among four studied triterpens are characterized by the ability to form pores in CHOL-enriched bilayers.
- Saponins (digitonin, tribulosin, dioscin, and escin) might decrease the membrane boundary potential. Sapogenins (diosgenin, uvaol, lupeol, and betulin) and nonglycosylated alkaloid solasodine are not characterized by this ability. Moreover, saponins can regulate the fluxes through ion channels, in particular, through cation-selective gramicidin A pores, by changing the transmembrane distribution of the electric potential. The dependence of the dipole-modifying ability on sapogenin glycosylation, the lack of the saponin effects on plasmanylcholine membranes as well as the absence of the correlation between the changes in the dipole potential and the lipophilicity of the compounds, indicate that saponins affect the dipole potential by altering the membrane hydration layer.
- The ability of saponins and related compounds to affect the phase behavior of membrane-forming lipids depends on the structure of lipophilic part of molecule (steroid or triterpenic). Steroids (digitonin, tribulosin, dioscin, diosgenin, and solasodine) are characterized by more pronounced effects in CHOL-free DPPC bilayers than in CHOL-enriched DPPC membranes. Triterpenes (uvaol, lupeol, and betulin) show an inverse relationship. Triterpenic saponin escin has no effect on lipid melting independent of the vesicle composition. The dependence of the effects on the structure of the lipophilic core of molecule and the presence of CHOL in the bilayer might indicate that tested agents affect lipid melting mainly through immersion into membrane and disordering of the lipids. The influence of saponins on the elastic properties of the membrane determines the possibility of regulating mechanosensitive ion channels, such as asymmetric peptide-lipid pores formed by syringomycin E.
- The effects of saponins and related compounds on the membrane permeability for the fluorescent marker dramatically depend on the agent structure and the liposome composition. The sapogenins (diosgenin, uvaol, lupeol, and betulin) and nonglycosylated alkaloid solasodine, increase permeability of POPC, POPC/Chol, and POPG vesicles for calcein in the same manner. Steroid saponins (digitonin, tribulosin, and dioscin) are characterized by the greatest efficiency in POPC/CHOL bilayers, while triterpenic saponin escin is almost equally effective in POPC/Chol and POPG membranes. The ability of saponins and related compounds to disengage the fluorescent marker from lipid vesicles might be determined by their abilities to disorder membrane lipids and induce the positive membrane curvature stress. The formation of the micellar-like structures in the presence of the tested chemicals might also facilitate ion and calcein transport through the membranes.
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yan, L.L.; Zhang, Y.J.; Gao, W.Y.; Man, S.L.; Wang, Y. In vitro and in vivo anticancer activity of steroid saponins of Paris polyphylla var. Yunnanensis. Exp. Oncol. 2009, 31, 27–32. [Google Scholar]
- Yassin, A.M.; El-Deeb, N.M.; Metwaly, A.M.; Fawal, G.F.; Radwan, M.M.; Hafez, E.E. Induction of apoptosis in human cancer cells through extrinsic and intrinsic pathways by balanites Aegyptiaca furostanol saponins and saponin-coated silvernanoparticles. Appl. Biochem. Biotechnol. 2017, 182, 1675–1693. [Google Scholar] [CrossRef]
- Gestetner, B.; Assa, Y.; Henis, Y.; Tencer, Y.; Rotman, M.; Birk, Y.; Bondi, A. Interaction of leucerne saponins with steroids. Biochim. Biophys. Acta 1972, 270, 181–187. [Google Scholar] [CrossRef]
- Vinarova, L.; Vinarov, Z.; Atanasov, V.; Pantcheva, I.; Tcholakova, S.; Denkov, N.; Stoyanov, S. Lowering of cholesterol bioaccessibility and serum concentrations by saponins: In vitro and in vivo studies. Food Funct. 2015, 6, 501–512. [Google Scholar] [CrossRef]
- Netala, V.R.; Ghosh, S.B.; Bobbu, P.; Anitha, D.; Tartte, V. Triterpenoid saponins: A review on biosynthesis, applications and mechanism of their action. Int. J. Pharm. Pharm. Sci. 2015, 7, 24–28. [Google Scholar]
- Lorent, J.H.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.P. The amphiphilic nature of saponins and their effects on artificial and biological membranes and potential consequences for red blood and cancer cells. Org. Biomol. Chem. 2014, 12, 8803–8822. [Google Scholar] [CrossRef] [PubMed]
- Manrique-Moreno, M.; Londoño-Londoño, J.; Jemioła-Rzemińska, M.; Strzałka, K.; Villena, F.; Avello, M.; Suwalsky, M. Structural effects of the solanum steroids solasodine, diosgenin and solanine on human erythrocytes and molecular models of eukaryotic membranes. Biochim. Biophys. Acta 2014, 1838, 266–277. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, M.; Nojima, S.; Akiyama, T.; Sankawa, U.; Inoue, K. Interaction of digitonin and its analogs with membrane cholesterol. J. Biochem. 1984, 96, 1231–1239. [Google Scholar] [CrossRef]
- Lorent, J.; Lins, L.; Domenech, O.; Quetin-Leclercq, J.; Brasseur, R.; Mingeot-Leclercq, M.-P. Domain formation and permeabilization induced by the saponin α-hederin and its aglycone hederagenin in a cholesterol-containing bilayer. Langmuir 2014, 30, 4556–4569. [Google Scholar] [CrossRef]
- Sreij, R.; Dargel, C.; Moleiro, L.H.; Monroy, F.; Hellweg, T. Aescin incorporation and nanodomain formation in DMPC model membranes. Langmuir 2017, 33, 12351–12361. [Google Scholar] [CrossRef] [PubMed]
- Naruse, M.; Suetomo, H.; Matsubara, T.; Sato, T.; Yanagawa, H.; Hoshi, M.; Matsumoto, M. Acrosome reaction-related steroidal saponin, Co-ARIS, from the starfish induces structural changes in microdomains. Dev. Biol. 2010, 347, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Keukens, E.A.; de Vrije, T.; van den Boom, C.; de Waard, P.; Plasman, H.H.; Thiel, F.; Chupin, V.; Jongen, W.M.; de Kruijff, B. Molecular basis of glycoalkaloid induced membrane disruption Biochim. Biophys. Acta 1995, 1240, 216–280. [Google Scholar] [CrossRef] [Green Version]
- Li, X.X.; Davis, B.; Haridas, V.; Gutterman, J.U.; Colombini, M. Proapoptotic triterpene electrophiles (avicins) form channels in membranes: Cholesterol dependence. Biophys. J. 2005, 88, 2577–2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, F.; Wang, R. Hemolytic mechanism of dioscin proposed by molecular dynamics simulations J. Mol. Model. 2010, 16, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Lorent, J.; Le Duff, C.S.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.P. Induction of highly curved structures in relation to membrane permeabilization and budding by the triterpenoid saponins, α- and δ-Hederin. J. Biol. Chem. 2013, 288, 14000–14017. [Google Scholar] [CrossRef] [Green Version]
- Gogelein, H.; Huby, A. Interaction of saponin and digitonin with black lipid membranes and lipid monolayers. Biochim. Biophys. Acta 1984, 773, 32–38. [Google Scholar] [CrossRef]
- Bangham, A.D.; Horne, R.W.; Glauert, A.M.; Dingle, J.T.; Lucy, J.A. Action of saponin on biological cell membranes. Nature 1962, 196, 952–955. [Google Scholar] [CrossRef] [PubMed]
- Efimova, S.S.; Ostroumova, O.S. Effect of dipole modifiers on the magnitude of the dipole potential of sterol-containing bilayers. Langmuir 2012, 28, 9908–9914. [Google Scholar] [CrossRef]
- Ostroumova, O.S.; Efimova, S.S.; Schagina, L.V. Phloretin induced reduction in dipole potential of sterol containing bilayers. J. Membr. Biol. 2013, 246, 985–991. [Google Scholar] [CrossRef]
- Efimova, S.S.; Zakharova, A.A.; Medvedev, R.Y.; Ostroumova, O.S. Ion channels induced by antimicrobial agents in model lipid membranes are modulated by plant polyphenols through surrounding lipid media. J. Membr. Biol. 2018, 251, 551–562. [Google Scholar] [CrossRef]
- Efimova, S.S.; Zakharova, A.A.; Ostroumova, O.S. Alkaloids modulate the functioning of ion channels produced by antimicrobial agents via an influence on the lipid host. Front. Cell Dev. Biol. 2020, 8, 537. [Google Scholar] [CrossRef] [PubMed]
- Gawrisch, K.; Ruston, D.; Zimmerberg, J.; Parsegian, V.A.; Rand, R.P.; Fuller, N. Membrane dipole potentials, hydration forces, and the ordering of water at membrane surfaces. Biophys. J. 1992, 61, 1213–1223. [Google Scholar] [CrossRef] [Green Version]
- Ermakov, Y.A.; Nesterenko, A.M. Boundary potential of lipid bilayers: Methods and interpretations. J. Phys. Conf. Ser. 2017, 780, 012002. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.-Y.; Olijve, L.L.C.; Kausik, R.; Han, S. Cholesterol enhances surface water diffusion of phospholipid bilayers. J. Chem. Phys. 2014, 141, 22D513. [Google Scholar] [CrossRef] [Green Version]
- Sommer, A.; Paltauf, F.; Hermetter, A. Dipolar solvent relaxation on a nanosecond time scale in ether phospholipid membranes as determined by multifrequency phase and modulation fluorometry. Biochemistry 1990, 29, 11134–11140. [Google Scholar] [CrossRef]
- Kruczek, J.; Saunders, M.; Khosla, M.; Tu, Y.; Pandit, S.A. Molecular dynamics simulations of ether- and ester-linked phospholipids. Biochim. Biophys. Acta Biomembr. 2017, 1859, 2297–2307. [Google Scholar] [CrossRef]
- Rokitskaya, T.I.; Antonenko, Y.N.; Kotova, E.A. Effect of the dipole potential of a bilayer lipid membrane on gramicidin channel dissociation kinetics. Biophys. J. 1997, 73, 850–854. [Google Scholar] [CrossRef] [Green Version]
- Koynova, R.; Caffrey, M. Phases and phase transitions of the phosphatidylcholines. Biochim. Biophys. Acta 1998, 1376, 91–145. [Google Scholar] [CrossRef]
- Wang, P.Y.; Lu, J.Z.; Chen, J.W.; Hwang, F. Interaction of the interdigitated DPPG or DPPG/DMPC bilayer with human erythrocyte band 3: Differential scanning calorimetry and fluorescence studies. Chem. Phys. Lipids 1994, 69, 241–249. [Google Scholar] [CrossRef]
- Chen, X.; Hua, W.; Huang, Z.; Allen, H.C. Interfacial water structure associated with phospholipid membranes studied by phase-sensitive vibrational sum frequency generation spectroscopy. J. Am. Chem. Soc. 2010, 132, 11336–11342. [Google Scholar] [CrossRef] [PubMed]
- Offringa, J.C.; Plekkenpol, R.; Crommelin, D.J. A differential scanning calorimetry study of the thermal behavior of water-dipalmitoylphosphatidylcholine mixtures at subzero temperatures: Effects of water content, surface charge, and cholesterol. J. Pharm. Sci. 1987, 76, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Jang, H.; Harlen, K.; Puri, A.; Nussinov, R.; Schneider, J.P.; Blumenthal, R. Mechanism of membrane permeation induced by synthetic β-hairpin peptides. Biophys. J. 2013, 105, 2093–2103. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, J.N.; Klausner, R.D.; Innerarity, T.; Ralston, E.; Blumenthal, R. Phase transition release, a new approach to the interaction of proteins with lipid vesicles. Application to lipoproteins. Biochim. Biophys. Acta 1981, 647, 270–284. [Google Scholar] [CrossRef]
- Chulkov, E.G.; Schagina, L.V.; Ostroumova, O.S. Membrane dipole modifiers modulate single-length nystatin channels via reducing elastic stress in the vicinity of the lipid mouth of a pore. Biochim. Biophys. Acta Biomembranes. 2015, 1848, 192–199. [Google Scholar] [CrossRef] [Green Version]
- Malev, V.V.; Schagina, L.V.; Gurnev, P.A.; Takemoto, J.Y.; Nestorovich, E.M.; Bezrukov, S.M. Syringomycin E channel: A lipidic pore stabilized by lipopeptide? Biophys. J. 2002, 82, 1985–1994. [Google Scholar] [CrossRef] [Green Version]
- Ostroumova, O.S.; Gurnev, P.A.; Schagina, L.V.; Bezrukov, S.M. Asymmetry of syringomycin E channel studied by polymer partitioning. FEBS Lett. 2007, 581, 804–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidwai, A.P.; Takemoto, J.Y. Bacterial phytotoxin, syringomycin, induces a protein kinase-mediated phosphorylation of red beet plasma membrane polypeptides. Proc. Natl. Acad. Sci. USA 1987, 84, 6755–6759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montal, M.; Muller, P. Formation of bimolecular membranes from lipid monolayers and study of their electrical properties. Proc. Nat. Acad. Sci. USA 1972, 65, 3561–3566. [Google Scholar] [CrossRef] [Green Version]
- Andersen, O.S.; Finkelstein, A.; Katz, I.; Cass, A. Effect of phloretin on the permeability of thin lipid membranes. J. Gen. Physiol. 1976, 67, 749–771. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Charge * | LogDo/w * | C, μM & | −Δφb(max), mV | |||
|---|---|---|---|---|---|---|---|
| DPhPC | DPhPC/CHOL | DPhPC | DPhPC/CHOL | HOPC | |||
| digitonin | 0.00 | −4.96 | 150 ± 25 | 75 ± 25 | 36 ± 4 | 27 ± 5 | 4 ± 2 |
| tribulosin | na | na | 200 ± 25 | 125 ± 25 | 47 ± 6 | 22 ± 9 | 6 ± 2 |
| dioscin | 0.00 | 1.71 | 150 ± 25 | 100 ± 25 | 39 ± 8 | 26 ± 9 | 5 ± 1 |
| diosgenin | 0.00 | 4.93 | 150 ± 25 | 125 ± 25 | 6 ± 2 | 5 ± 3 | 3 ± 2 |
| solasodine | 0.99 | 2.50 | 150 ± 25 | 100 ± 25 | 5 ± 2 | 7 ± 2 | 2 ± 1 |
| escin | −1.00 | −4.29 | 150 ± 25 | 75 ± 25 | 20 ± 5 | 16 ± 4 | 5 ± 2 |
| uvaol | 0.00 | 6.11 | 200 ± 25 | 150 ± 25 | 1 ± 1 | 1 ± 1 | 5 ± 3 |
| lupeol | 0.00 | 7.45 | 200 ± 25 | 150 ± 25 | 1 ± 1 | 1 ± 1 | 3 ± 1 |
| betulin | 0.00 | 6.17 | 150 ± 25 | 100 ± 25 | 1 ± 1 | 3 ± 1 | 3 ± 2 |
| Compound | GSC, pS | τ, s | |
|---|---|---|---|
| – | 21.5 ± 0.5 | 3.0 ± 1.0 | |
| tribulosin | 23.6 ± 0.8 | 4.7 ± 0.3 | |
| lupeol | 20.9 ± 0.6 | 3.1 ± 0.6 |
| Compound | −∆Tm, °C | ∆T1/2, °C | ||||
|---|---|---|---|---|---|---|
| DPPC | DPPC/CHOL | DPPG | DPPC | DPPC/CHOL | DPPG | |
| digitonin | 0.7 ± 0.1 | 0.4 ± 0.1 | 0 | 0.4 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| tribulosin | 0.9 ± 0.2 | 0.5 ± 0.1 | 0 | 0.3 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| dioscin | 0.8 ± 0.2 | 0.5 ± 0.1 | 0.1 ± 0.1 | 0.7 ± 0.2 | 0.5 ± 0.2 | 0.2 ± 0.1 |
| diosgenin | 0.4 ± 0.1 | 0.2 ± 0.1 | 0 | 0.5 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| solasodine | 0.5 ± 0.1 | 0.3 ± 0.1 | 0 | 0.4 ± 0.1 | 0.2 ± 0.1 | 0.1 ± 0.1 |
| escin | 0 | 0 | 0 | 0 | 0 | 0 |
| uvaol | 0.2 ± 0.1 | 0.8 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.3 ± 0.1 | 0.1 ± 0.1 |
| lupeol | 0.3 ± 0.1 | 0.4 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.2 ± 0.1 |
| betulin | 0.2 ± 0.1 | 0.5 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| Compound | SrE | NyS |
|---|---|---|
| dioscin | 19.1 ± 6.8 | 1.0 ± 0.1 |
| uvaol | 1.1 ± 0.1 | 0.8 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Efimova, S.S.; Ostroumova, O.S. Is the Membrane Lipid Matrix a Key Target for Action of Pharmacologically Active Plant Saponins? Int. J. Mol. Sci. 2021, 22, 3167. https://doi.org/10.3390/ijms22063167
Efimova SS, Ostroumova OS. Is the Membrane Lipid Matrix a Key Target for Action of Pharmacologically Active Plant Saponins? International Journal of Molecular Sciences. 2021; 22(6):3167. https://doi.org/10.3390/ijms22063167
Chicago/Turabian StyleEfimova, Svetlana S., and Olga S. Ostroumova. 2021. "Is the Membrane Lipid Matrix a Key Target for Action of Pharmacologically Active Plant Saponins?" International Journal of Molecular Sciences 22, no. 6: 3167. https://doi.org/10.3390/ijms22063167
APA StyleEfimova, S. S., & Ostroumova, O. S. (2021). Is the Membrane Lipid Matrix a Key Target for Action of Pharmacologically Active Plant Saponins? International Journal of Molecular Sciences, 22(6), 3167. https://doi.org/10.3390/ijms22063167