
Investigating Primary Cilia during Peripheral Nervous System Formation


Abstract
:1. Introduction
2. Results
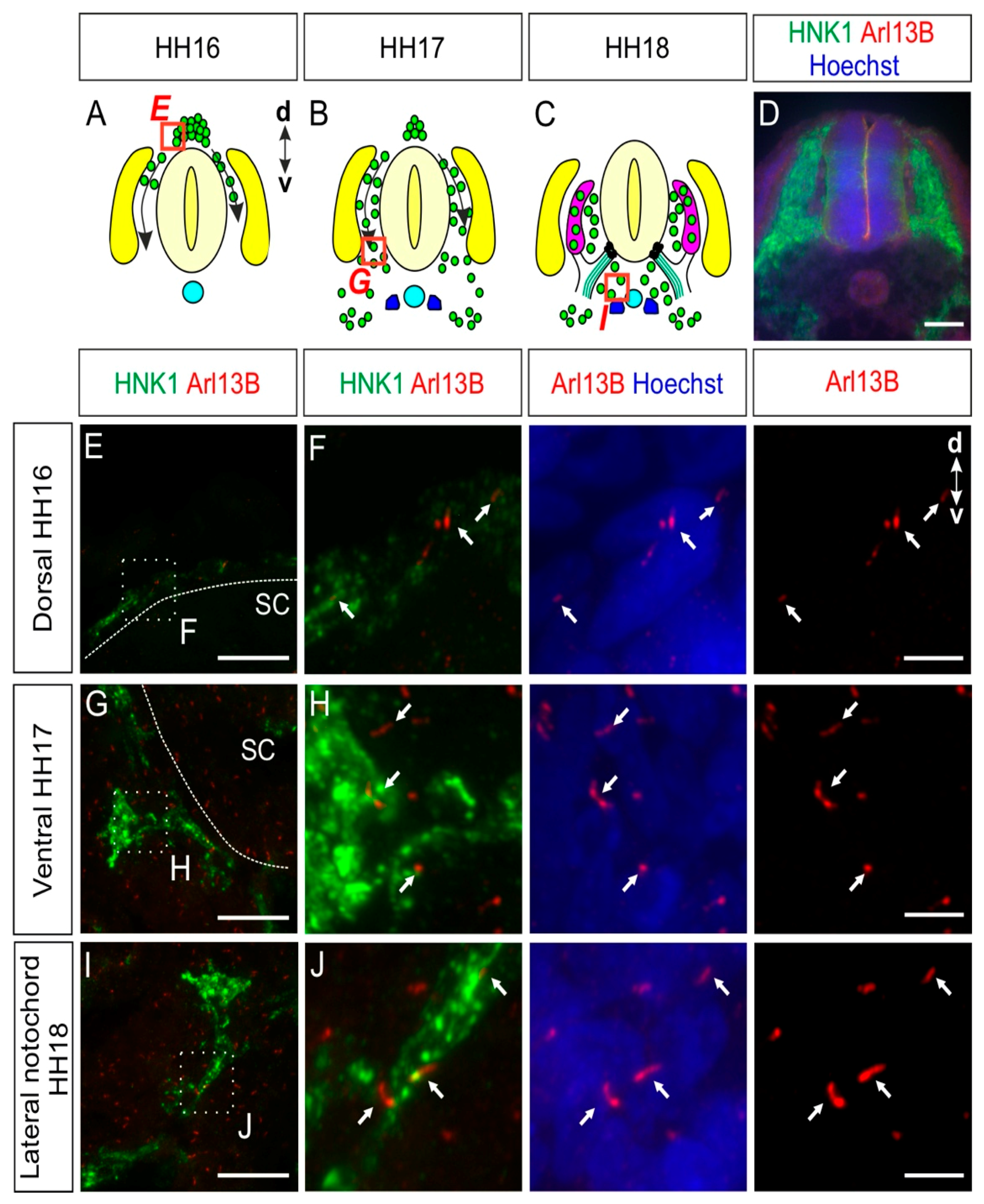
2.1. Trunk Neural Crest Cells Bear a Primary Cilium during Their Early Migration
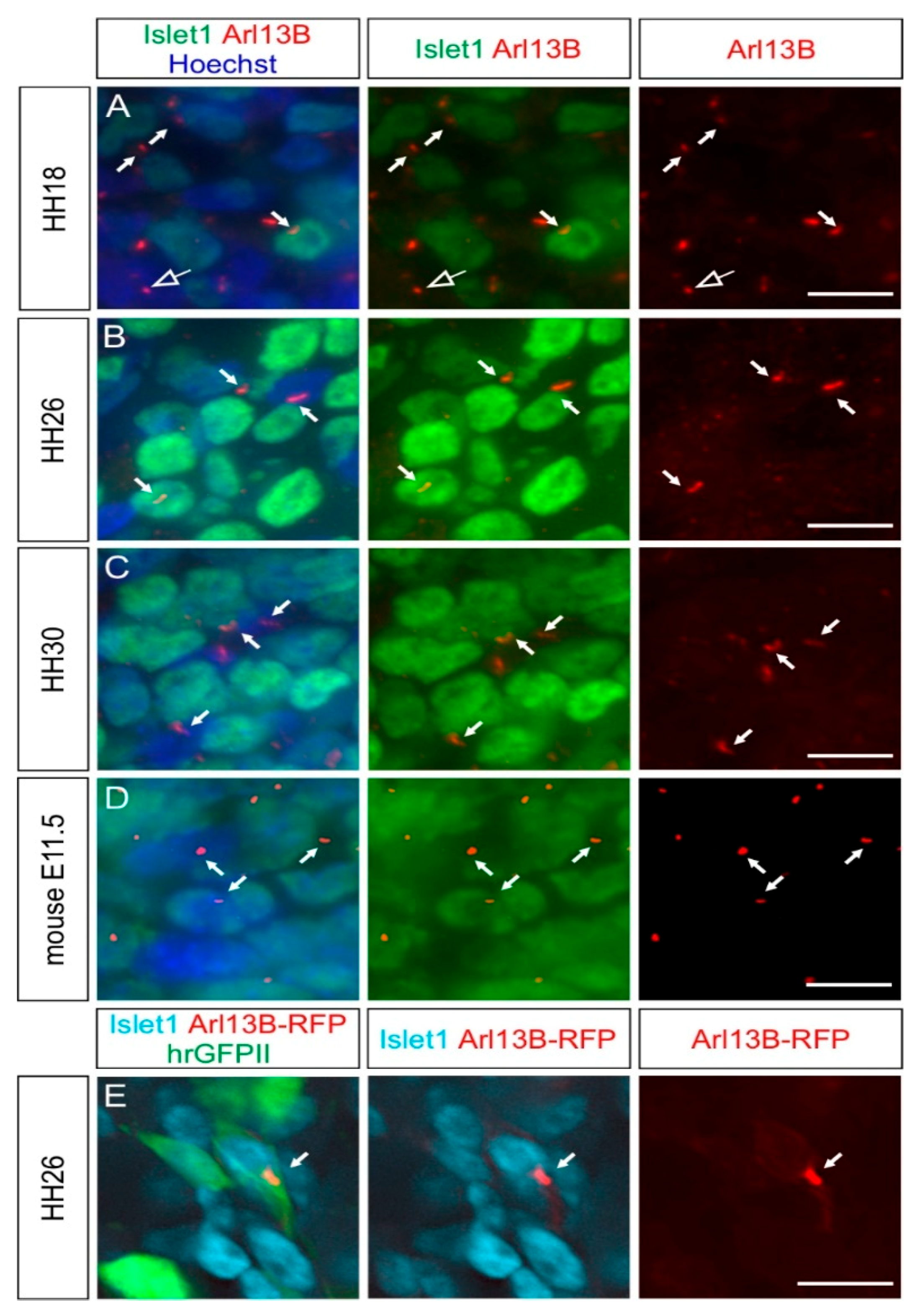
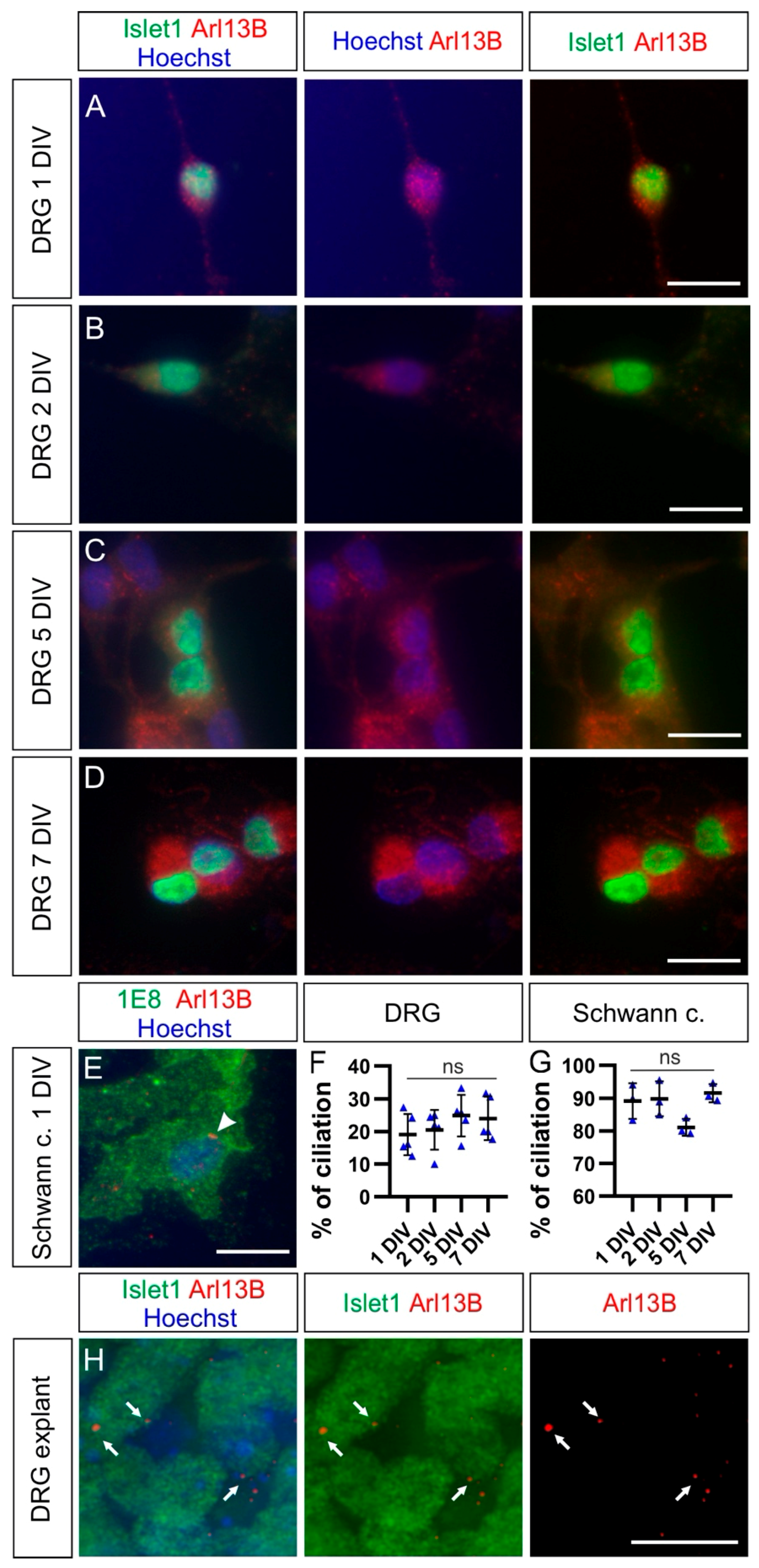
2.2. Dorsal Root Ganglia Neurons Carry a Primary Cilium during Development
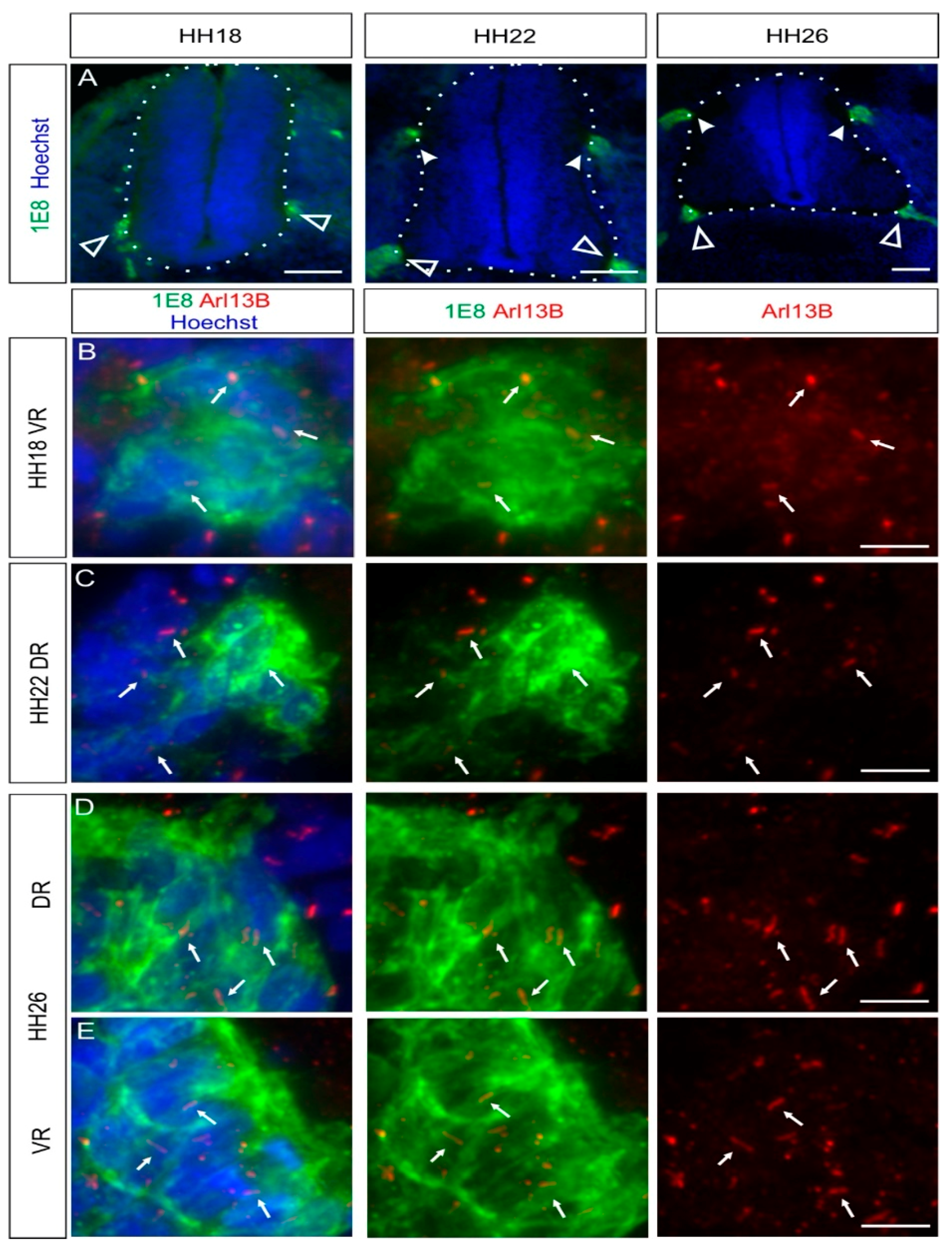
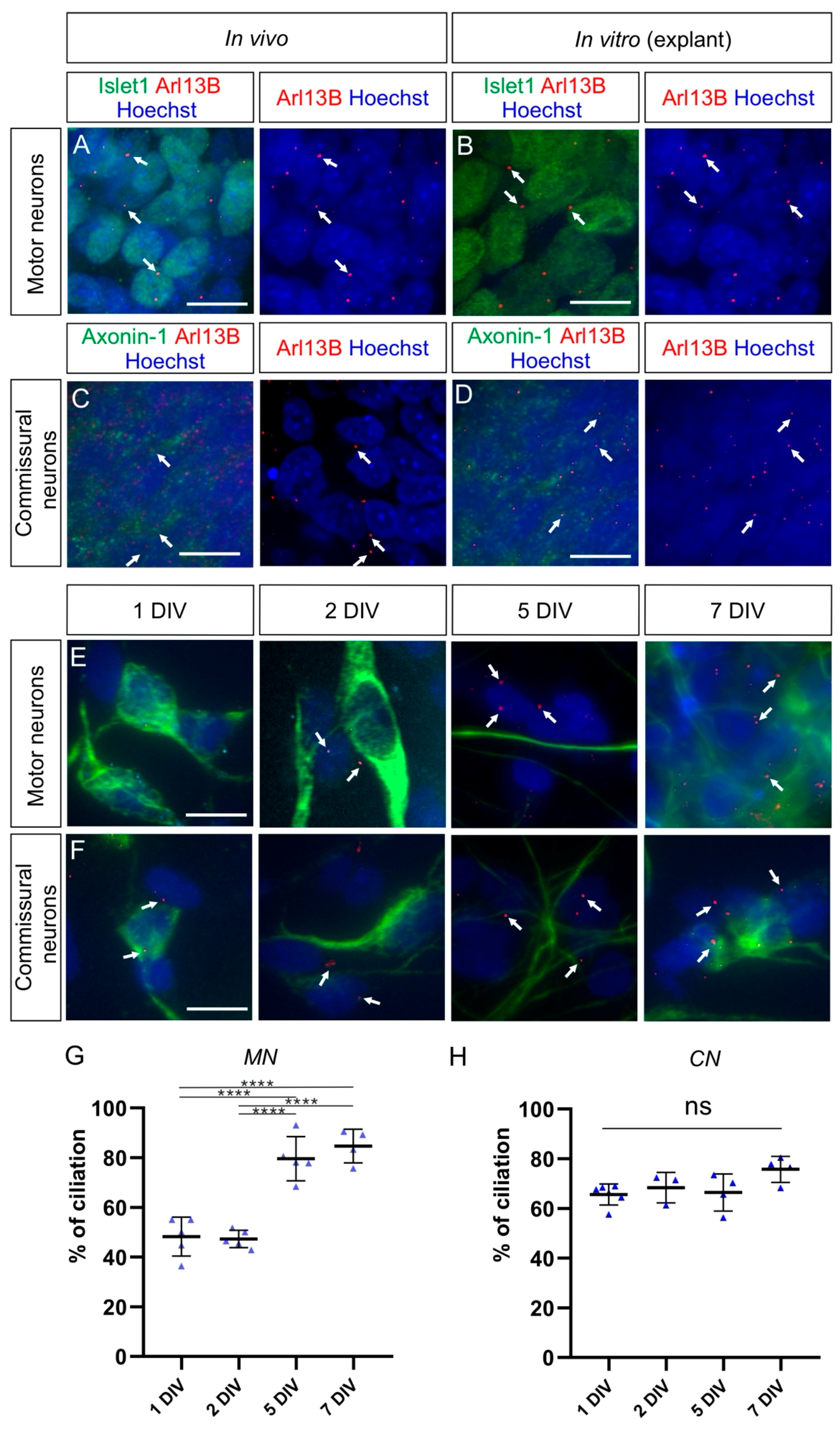
2.3. Boundary Cap Cells Bear a Primary Cilium at Both Ventral and Dorsal Roots
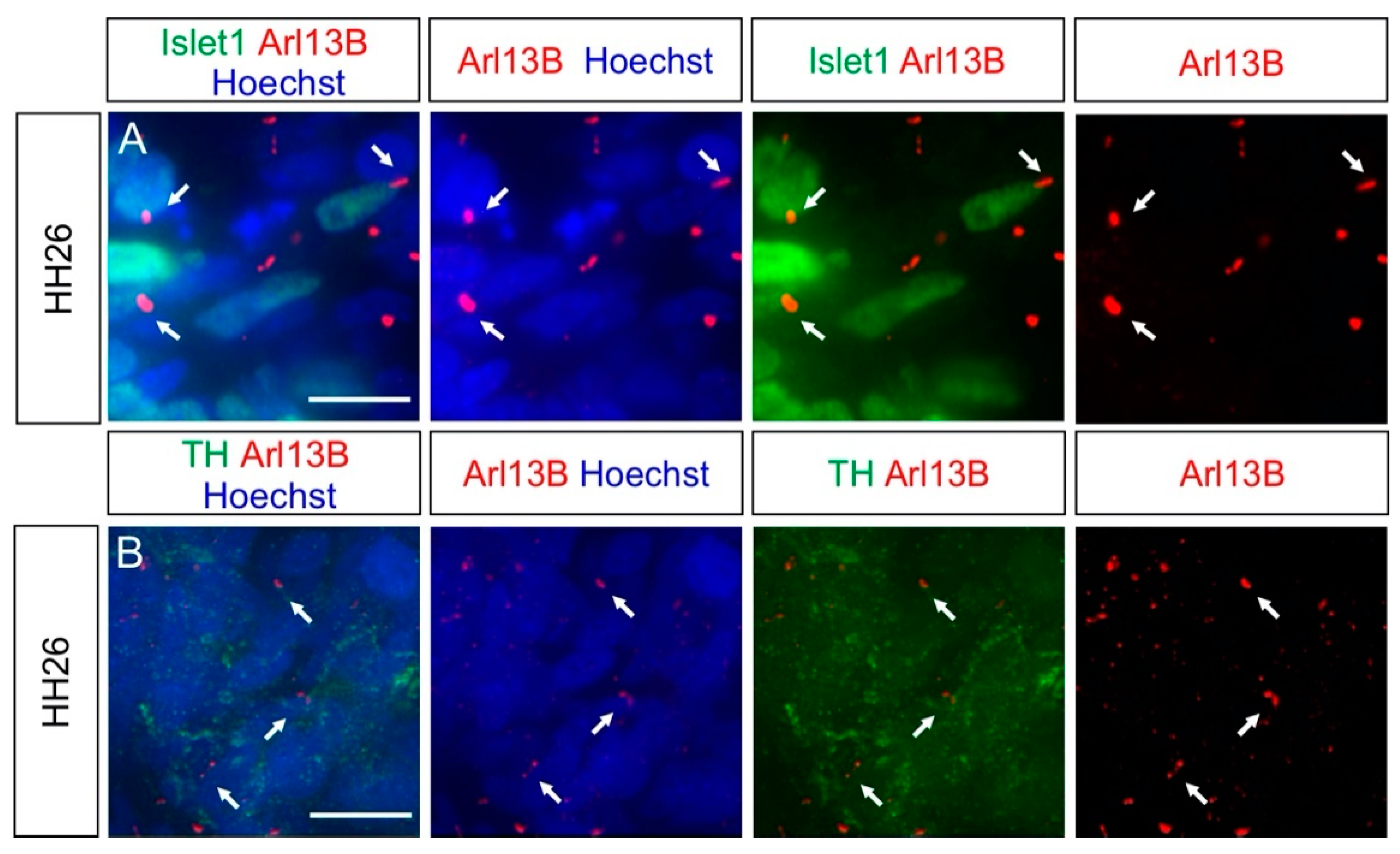
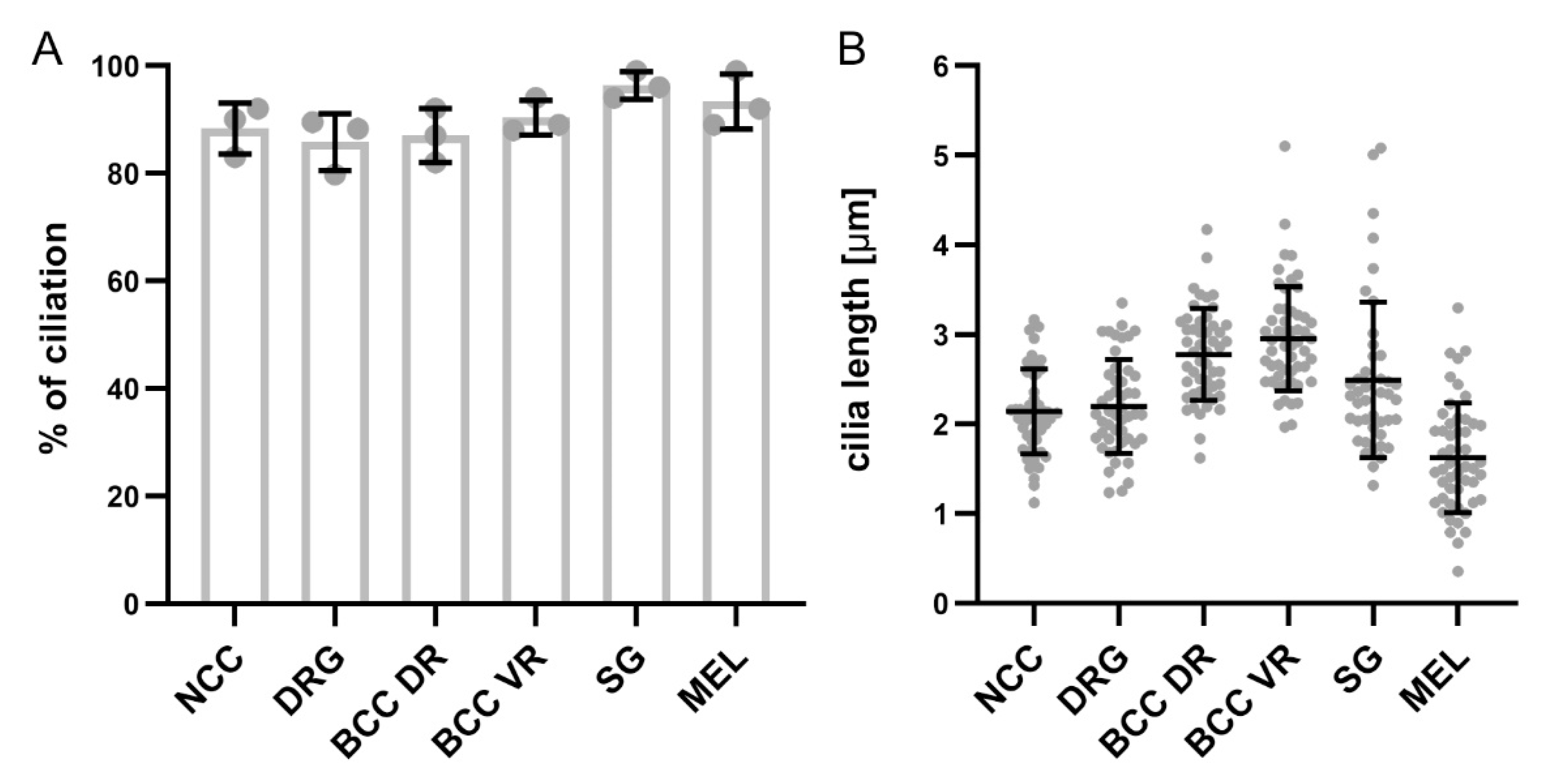
2.4. Carrying a Primary Cilium Appears to Be a Common Feature on NCC Derivatives In Vivo
2.5. Cultured Neurons Reveal Heterogeneity in Their Ciliation
3. Discussion
4. Materials and Methods
4.1. Embryo Dissection and Fixation
4.2. In Ovo Electroporation
4.3. Immunohistochemistry
4.4. Immunohistochemistry for Primary Cilia
4.5. Cultures of Dissociated Neurons
4.6. Explants Cultures
4.7. Immunocytochemistry
4.8. Microscopy
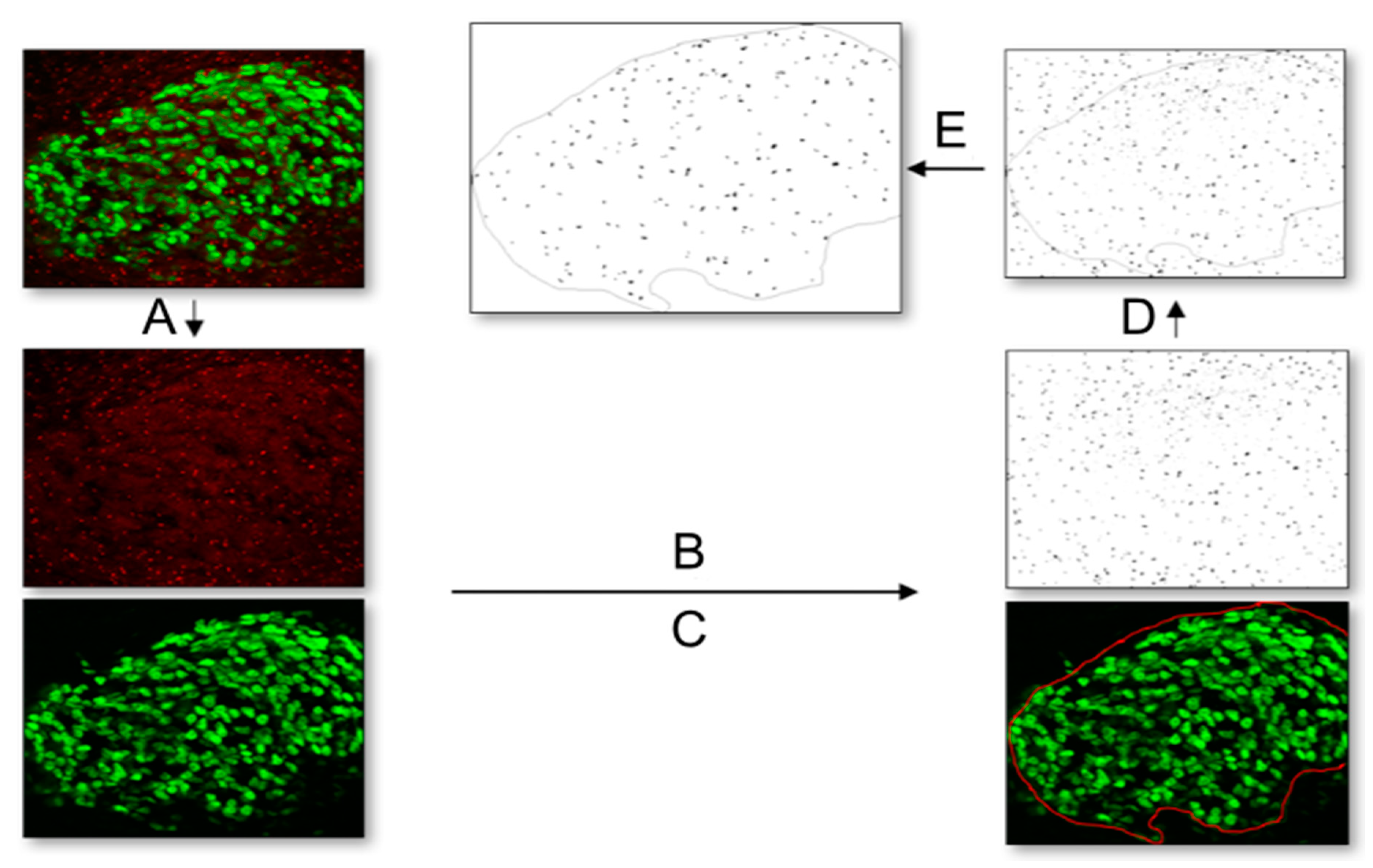
4.9. Quantification of Ciliation Rate and Cilia Length
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CNS | central nervous system |
| PNS | peripheral nervous system |
| DRG | dorsal root ganglia |
| BCC | boundary cap cells |
| NCC | neural crest cells |
| SG | sympathetic ganglia |
| HH | Hamburger and Hamilton stage |
| DIV | day in vitro |
References
- Broekhuis, J.R.; Leong, W.Y.; Jansen, G. Regulation of Cilium Length and Intraflagellar Transport. In International Review of Cell and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 303, pp. 101–138. ISBN 978-0-12-407697-6. [Google Scholar]
- Gerdes, J.M.; Davis, E.E.; Katsanis, N. The Vertebrate Primary Cilium in Development, Homeostasis, and Disease. Cell 2009, 137, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.C.; Katsanis, N. Cilia in Vertebrate Development and Disease. Development 2012, 139, 443–448. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, K.W. Beiträge zur Kenntniss einiger Drüsen und Epithelien: Hierzu Tafel XXVII, XXVIII u. XXIX. Arch. Mikrosk. Anat. 1898, 52, 552–706. [Google Scholar] [CrossRef]
- Wong, S.Y.; Reiter, J.F. The Primary Cilium at the Crossroads of Mammalian Hedgehog Signaling. Curr. Top. Dev. Biol. 2008, 85, 225–260. [Google Scholar] [CrossRef] [PubMed]
- Huangfu, D.; Anderson, K.V. Cilia and Hedgehog Responsiveness in the Mouse. Proc. Natl. Acad. Sci. USA 2005, 102, 11325–11330. [Google Scholar] [CrossRef] [Green Version]
- Otto, E.A.; Schermer, B.; Obara, T.; O’Toole, J.F.; Hiller, K.S.; Mueller, A.M.; Ruf, R.G.; Hoefele, J.; Beekmann, F.; Landau, D.; et al. Mutations in INVS Encoding Inversin Cause Nephronophthisis Type 2, Linking Renal Cystic Disease to the Function of Primary Cilia and Left-Right Axis Determination. Nat. Genet. 2003, 34, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Gloy, J.; Ganner, A.; Bullerkotte, A.; Bashkurov, M.; Krönig, C.; Schermer, B.; Benzing, T.; Cabello, O.A.; Jenny, A.; et al. Inversin, the Gene Product Mutated in Nephronophthisis Type II, Functions as a Molecular Switch between Wnt Signaling Pathways. Nat. Genet. 2005, 37, 537–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, L.; Clement, C.A.; Teilmann, S.C.; Pazour, G.J.; Hoffmann, E.K.; Satir, P.; Christensen, S.T. PDGFRalphaalpha Signaling Is Regulated through the Primary Cilium in Fibroblasts. Curr. Biol. 2005, 15, 1861–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, F.M.; Schou, K.B.; Vilhelm, M.J.; Holm, M.S.; Breslin, L.; Farinelli, P.; Larsen, L.A.; Andersen, J.S.; Pedersen, L.B.; Christensen, S.T. IFT20 Modulates Ciliary PDGFRα Signaling by Regulating the Stability of Cbl E3 Ubiquitin Ligases. J. Cell Biol. 2018, 217, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Boehlke, C.; Kotsis, F.; Patel, V.; Braeg, S.; Voelker, H.; Bredt, S.; Beyer, T.; Janusch, H.; Hamann, C.; Gödel, M.; et al. Primary Cilia Regulate MTORC1 Activity and Cell Size through Lkb1. Nat. Cell Biol. 2010, 12, 1115–1122. [Google Scholar] [CrossRef] [Green Version]
- Satir, P.; Pedersen, L.B.; Christensen, S.T. The Primary Cilium at a Glance. J. Cell Sci. 2010, 123, 499–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchison, H.M.; Valente, E.M. Motile and Non-Motile Cilia in Human Pathology: From Function to Phenotypes: Motile and Non-Motile Ciliopathies. J. Pathol. 2017, 241, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Gleeson, J.G. A Systems-Biology Approach to Understanding the Ciliopathy Disorders. Genome Med. 2011, 3, 59. [Google Scholar] [CrossRef] [Green Version]
- Ware, S.M.; Aygun, M.G.; Hildebrandt, F. Spectrum of Clinical Diseases Caused By Disorders of Primary Cilia. Proc. Am. Thorac. Soc. 2011, 8, 444–450. [Google Scholar] [CrossRef] [Green Version]
- Hildebrandt, F.; Benzing, T.; Katsanis, N. Ciliopathies. N. Engl. J. Med. 2011, 364, 1533–1543. [Google Scholar] [CrossRef] [Green Version]
- Grochowsky, A.; Gunay-Aygun, M. Clinical Characteristics of Individual Organ System Disease in Non-Motile Ciliopathies. Transl. Sci. Rare Dis. 2019, 4, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Guemez-Gamboa, A.; Coufal, N.G.; Gleeson, J.G. Primary Cilia in the Developing and Mature Brain. Neuron 2014, 82, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Suciu, S.K.; Caspary, T. Cilia, Neural Development and Disease. Semin. Cell Dev. Biol. 2020, S1084952119301727. [Google Scholar] [CrossRef]
- Métin, C.; Pedraza, M. Cilia: Traffic Directors along the Road of Cortical Development. Neuroscientist 2014, 20, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Asadollahi, R.; Strauss, J.E.; Zenker, M.; Beuing, O.; Edvardson, S.; Elpeleg, O.; Strom, T.M.; Joset, P.; Niedrist, D.; Otte, C.; et al. Clinical and Experimental Evidence Suggest a Link between KIF7 and C5orf42-Related Ciliopathies through Sonic Hedgehog Signaling. Eur. J. Hum. Genet. 2018, 26, 197–209. [Google Scholar] [CrossRef]
- Tan, P.L.; Barr, T.; Inglis, P.N.; Mitsuma, N.; Huang, S.M.; Garcia-Gonzalez, M.A.; Bradley, B.A.; Coforio, S.; Albrecht, P.J.; Watnick, T.; et al. Loss of Bardet Biedl Syndrome Proteins Causes Defects in Peripheral Sensory Innervation and Function. Proc. Natl. Acad. Sci. USA 2007, 104, 17524–17529. [Google Scholar] [CrossRef] [Green Version]
- Etchevers, H.C.; Dupin, E.; Le Douarin, N.M. The Diverse Neural Crest: From Embryology to Human Pathology. Development 2019, 146, dev169821. [Google Scholar] [CrossRef] [Green Version]
- Theveneau, E.; Mayor, R. Neural Crest Delamination and Migration: From Epithelium-to-Mesenchyme Transition to Collective Cell Migration. Dev. Biol. 2012, 366, 34–54. [Google Scholar] [CrossRef] [Green Version]
- Marmigère, F.; Ernfors, P. Specification and Connectivity of Neuronal Subtypes in the Sensory Lineage. Nat. Rev. Neurosci. 2007, 8, 114–127. [Google Scholar] [CrossRef]
- Giovannone, D.; Ortega, B.; Reyes, M.; El-Ghali, N.; Rabadi, M.; Sao, S.; De Bellard, M.E. Chicken Trunk Neural Crest Migration Visualized with HNK1. Acta Histochem. 2015, 117, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Caspary, T.; Larkins, C.E.; Anderson, K.V. The Graded Response to Sonic Hedgehog Depends on Cilia Architecture. Dev. Cell 2007, 12, 767–778. [Google Scholar] [CrossRef] [Green Version]
- Dumoulin, A.; Schmidt, H.; Rathjen, F.G. Sensory Neurons: The Formation of T-Shaped Branches Is Dependent on a CGMP-Dependent Signaling Cascade. Neuroscientist 2021, 27, 47–57. [Google Scholar] [CrossRef]
- Cole, D.G.; Diener, D.R.; Himelblau, A.L.; Beech, P.L.; Fuster, J.C.; Rosenbaum, J.L. Chlamydomonas Kinesin-II–Dependent Intraflagellar Transport (IFT): IFT Particles Contain Proteins Required for Ciliary Assembly in Caenorhabditis Elegans Sensory Neurons. J. Cell Biol. 1998, 141, 993–1008. [Google Scholar] [CrossRef]
- Perrin, F.E.; Rathjen, F.G.; Stoeckli, E.T. Distinct Subpopulations of Sensory Afferents Require F11 or Axonin-1 for Growth to Their Target Layers within the Spinal Cord of the Chick. Neuron 2001, 30, 707–723. [Google Scholar] [CrossRef] [Green Version]
- Fontenas, L.; Kucenas, S. Livin’ On The Edge: Glia Shape Nervous System Transition Zones. Curr. Opin. Neurobiol. 2017, 47, 44–51. [Google Scholar] [CrossRef]
- Radomska, K.J.; Topilko, P. Boundary Cap Cells in Development and Disease. Curr. Opin. Neurobiol. 2017, 47, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Ernsberger, U.; Rohrer, H. Sympathetic Tales: Subdivisons of the Autonomic Nervous System and the Impact of Developmental Studies. Neural Dev. 2018, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Tojima, T.; Hines, J.H.; Henley, J.R.; Kamiguchi, H. Second Messengers and Membrane Trafficking Direct and Organize Growth Cone Steering. Nat. Rev. Neurosci. 2011, 12, 191–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schock, E.N.; Brugmann, S.A. Discovery, Diagnosis, and Etiology of Craniofacial Ciliopathies. Cold Spring Harb. Perspect. Biol. 2017, 9, a028258. [Google Scholar] [CrossRef]
- Elliott, K.H.; Millington, G.; Brugmann, S.A. A Novel Role for Cilia-Dependent Sonic Hedgehog Signaling during Submandibular Gland Development: Novel Role for Cilia-Dependent Shh Signaling During SMG Development. Dev. Dyn. 2018, 247, 818–831. [Google Scholar] [CrossRef]
- Schock, E.N.; Struve, J.N.; Chang, C.-F.; Williams, T.J.; Snedeker, J.; Attia, A.C.; Stottmann, R.W.; Brugmann, S.A. A Tissue-Specific Role for Intraflagellar Transport Genes during Craniofacial Development. PLoS ONE 2017, 12, e0174206. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Feng, J.; Li, J.; Ho, T.-V.; Yuan, Y.; Liu, Y.; Brindopke, F.; Figueiredo, J.C.; Magee, W.; Sanchez-Lara, P.A.; et al. Intraflagellar Transport 88 (IFT88) Is Crucial for Craniofacial Development in Mice and Is a Candidate Gene for Human Cleft Lip and Palate. Hum. Mol. Genet. 2017, ddx002. [Google Scholar] [CrossRef]
- Millington, G.; Elliott, K.H.; Chang, Y.-T.; Chang, C.-F.; Dlugosz, A.; Brugmann, S.A. Cilia-Dependent GLI Processing in Neural Crest Cells Is Required for Tongue Development. Dev. Biol. 2017, 424, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Portal, C.; Rompolas, P.; Lwigale, P.; Iomini, C. Primary Cilia Deficiency in Neural Crest Cells Models Anterior Segment Dysgenesis in Mouse. eLife 2019, 8, e52423. [Google Scholar] [CrossRef] [PubMed]
- Tobin, J.L.; Di Franco, M.; Eichers, E.; May-Simera, H.; Garcia, M.; Yan, J.; Quinlan, R.; Justice, M.J.; Hennekam, R.C.; Briscoe, J.; et al. Inhibition of Neural Crest Migration Underlies Craniofacial Dysmorphology and Hirschsprung’s Disease in Bardet-Biedl Syndrome. Proc. Natl. Acad. Sci. USA 2008, 105, 6714–6719. [Google Scholar] [CrossRef] [Green Version]
- Willaredt, M.A.; Gorgas, K.; Gardner, H.A.R.; Tucker, K.L. Multiple Essential Roles for Primary Cilia in Heart Development. Cilia 2012, 1, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higginbotham, H.; Eom, T.-Y.; Mariani, L.E.; Bachleda, A.; Hirt, J.; Gukassyan, V.; Cusack, C.L.; Lai, C.; Caspary, T.; Anton, E.S. Arl13b in Primary Cilia Regulates the Migration and Placement of Interneurons in the Developing Cerebral Cortex. Dev. Cell 2012, 23, 925–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monaco, S.; Baur, K.; Hellwig, A.; Hölzl-Wenig, G.; Mandl, C.; Ciccolini, F. A Flow Cytometry-Based Approach for the Isolation and Characterization of Neural Stem Cell Primary Cilia. Front. Cell. Neurosci. 2019, 12, 519. [Google Scholar] [CrossRef] [Green Version]
- Maj, E.; Künneke, L.; Loresch, E.; Grund, A.; Melchert, J.; Pieler, T.; Aspelmeier, T.; Borchers, A. Controlled Levels of Canonical Wnt Signaling Are Required for Neural Crest Migration. Dev. Biol. 2016, 417, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Diaz, R.; Trainor, P.A. Signals and Switches in Mammalian Neural Crest Cell Differentiation. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [Green Version]
- Clement, C.A.; Ajbro, K.D.; Koefoed, K.; Vestergaard, M.L.; Veland, I.R.; Henriques de Jesus, M.P.R.; Pedersen, L.B.; Benmerah, A.; Andersen, C.Y.; Larsen, L.A.; et al. TGF-β Signaling Is Associated with Endocytosis at the Pocket Region of the Primary Cilium. Cell Rep. 2013, 3, 1806–1814. [Google Scholar] [CrossRef] [Green Version]
- Dumoulin, A.; Ter-Avetisyan, G.; Schmidt, H.; Rathjen, F. Molecular Analysis of Sensory Axon Branching Unraveled a CGMP-Dependent Signaling Cascade. Int. J. Mol. Sci. 2018, 19, 1266. [Google Scholar] [CrossRef] [Green Version]
- Davis, B.M.; Frank, E.; Johnson, F.A.; Scott, S.A. Development of Central Projections of Lumbosacral Sensory Neurons in the Chick. J. Comp. Neurol. 1989, 279, 556–566. [Google Scholar] [CrossRef]
- Mauti, O.; Domanitskaya, E.; Andermatt, I.; Sadhu, R.; Stoeckli, E.T. Semaphorin6A Acts as a Gate Keeper between the Central and the Peripheral Nervous System. Neural Dev. 2007, 2, 28. [Google Scholar] [CrossRef] [Green Version]
- Bron, R.; Vermeren, M.; Kokot, N.; Andrews, W.; Little, G.E.; Mitchell, K.J.; Cohen, J. Boundary Cap Cells Constrain Spinal Motor Neuron Somal Migration at Motor Exit Points by a Semaphorin-Plexin Mechanism. Neural Dev. 2007, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Golding, J.P.; Cohen, J. Border Controls at the Mammalian Spinal Cord: Late-Surviving Neural Crest Boundary Cap Cells at Dorsal Root Entry Sites May Regulate Sensory Afferent Ingrowth and Entry Zone Morphogenesis. Mol. Cell. Neurosci. 1997, 9, 381–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.-H.; Zanini, M.; Shirvani, H.; Cheng, J.-S.; Yu, H.; Feng, C.-H.; Mercier, A.L.; Hung, S.-Y.; Forget, A.; Wang, C.-H.; et al. Atoh1 Controls Primary Cilia Formation to Allow for SHH-Triggered Granule Neuron Progenitor Proliferation. Dev. Cell 2019, 48, 184–199.e5. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.C.; Lee, J.-W.; Lee, T.H.; Subedi, L.; Wahedi, H.M.; Do, S.-G.; Shin, E.; Moon, E.-Y.; Kim, S.Y. UP256 Inhibits Hyperpigmentation by Tyrosine Expression/Dendrite Formation via Rho-Dependent Signaling and by Primary Cilium Formation in Melanocytes. Int. J. Mol. Sci. 2020, 21, 5341. [Google Scholar] [CrossRef]
- Berbari, N.F.; Bishop, G.A.; Askwith, C.C.; Lewis, J.S.; Mykytyn, K. Hippocampal Neurons Possess Primary Cilia in Culture. J. Neurosci. Res. 2007, 85, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Kalil, K.; Dent, E.W. Branch Management: Mechanisms of Axon Branching in the Developing Vertebrate CNS. Nat. Rev. Neurosci. 2014, 15, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.E.; Suter, D.M. An Integrated Cytoskeletal Model of Neurite Outgrowth. Front. Cell. Neurosci. 2018, 12, 447. [Google Scholar] [CrossRef]
- Armijo-Weingart, L.; Gallo, G. It Takes a Village to Raise a Branch: Cellular Mechanisms of the Initiation of Axon Collateral Branches. Mol. Cell. Neurosci. 2017, 84, 36–47. [Google Scholar] [CrossRef]
- Pinskey, J.M.; Franks, N.E.; McMellen, A.N.; Giger, R.J.; Allen, B.L. Neuropilin-1 Promotes Hedgehog Signaling through a Novel Cytoplasmic Motif. J. Biol. Chem. 2017, 292, 15192–15204. [Google Scholar] [CrossRef] [Green Version]
- Choksi, S.P.; Lauter, G.; Swoboda, P.; Roy, S. Switching on Cilia: Transcriptional Networks Regulating Ciliogenesis. Development 2014, 141, 1427–1441. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.; Morlé, L.; Soulavie, F.; Laurençon, A.; Sagnol, S.; Durand, B. Transcriptional Control of Genes Involved in Ciliogenesis: A First Step in Making Cilia. Biol. Cell 2010, 102, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Oblinger, M.; Lasek, R. A Conditioning Lesion of the Peripheral Axons of Dorsal Root Ganglion Cells Accelerates Regeneration of Only Their Peripheral Axons. J. Neurosci. 1984, 4, 1736–1744. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Williams, P.R.; Jacobi, A.; Wang, C.; Goel, A.; Hirano, A.A.; Brecha, N.C.; Kerschensteiner, D.; He, Z. Elevating Growth Factor Responsiveness and Axon Regeneration by Modulating Presynaptic Inputs. Neuron 2019, 103, 39–51.e5. [Google Scholar] [CrossRef] [PubMed]
- Andermatt, I.; Wilson, N.; Stoeckli, E.T. In Ovo Electroporation of miRNA-Based-Plasmids to Investigate Gene Function in the Developing Neural Tube. In Gene Function Analysis; Ochs, M.F., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2014; Volume 1101, pp. 353–368. ISBN 978-1-62703-720-4. [Google Scholar]
- Wilson, N.H.; Stoeckli, E.T. Cell Type Specific, Traceable Gene Silencing for Functional Gene Analysis during Vertebrate Neural Development. Nucleic Acids Res. 2011, 39, e133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.M.; Senanayake, U.; Artibani, M.; Taylor, G.; Wells, D.; Ahmed, A.A.; Sauka-Spengler, T. Genome and Epigenome Engineering CRISPR Toolkit for in Vivo Modulation of Cis -Regulatory Interactions and Gene Expression in the Chicken Embryo. Development 2018, 145, dev160333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, S.; Piacentino, M.L.; Vieceli, F.M.; Bronner, M.E. Optimization of CRISPR/Cas9 Genome Editing for Loss-of-Function in the Early Chick Embryo. Dev. Biol. 2017, 432, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, V.; Hamilton, H.L. A Series of Normal Stages in the Development of the Chick Embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef]
- Andermatt, I.; Wilson, N.H.; Bergmann, T.; Mauti, O.; Gesemann, M.; Sockanathan, S.; Stoeckli, E.T. Semaphorin 6B Acts as a Receptor in Post-Crossing Commissural Axon Guidance. Development 2014, 141, 3709–3720. [Google Scholar] [CrossRef] [Green Version]
- Wilson, N.H.; Stoeckli, E.T. In Ovo Electroporation of MiRNA-Based Plasmids in the Developing Neural Tube and Assessment of Phenotypes by DiI Injection in Open-Book Preparations. JoVE 2012, 4384. [Google Scholar] [CrossRef] [Green Version]
- Das, R.M.; Storey, K.G. Apical Abscission Alters Cell Polarity and Dismantles the Primary Cilium During Neurogenesis. Science 2014, 343, 200–204. [Google Scholar] [CrossRef] [Green Version]
- Dumoulin, A.; Dagane, A.; Dittmar, G.; Rathjen, F.G. S-Palmitoylation Is Required for the Control of Growth Cone Morphology of DRG Neurons by CNP-Induced CGMP Signaling. Front. Mol. Neurosci. 2018, 11, 345. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DRG | Schwann Cells | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 DIV | 2 DIV | 5 DIV | 7 DIV | 1 DIV | 2 DIV | 5 DIV | 7 DIV | |
| average: | 19% | 20% | 25% | 24% | 89% | 90% | 81% | 92% |
| standard deviation: | 6% | 6% | 6% | 7% | 5% | 5% | 3% | 3% |
| N(replicates): | 5 | 5 | 5 | 5 | 3 | 3 | 3 | 3 |
| Commissural Neurons | Motor Neurons | |||||||
| 1 DIV | 2 DIV | 5 DIV | 7 DIV | 1 DIV | 2 DIV | 5 DIV | 7 DIV | |
| average: | 66% | 68% | 66% | 76% | 48% | 47% | 80% | 85% |
| standard deviation: | 4% | 6% | 7% | 5% | 8% | 4% | 9% | 7% |
| N(replicates): | 4 | 4 | 4 | 4 | 6 | 6 | 4 | 4 |
| Antigen | Species | Source | Cat# | Dilution |
|---|---|---|---|---|
| Arl13B | Rabbit (polyclonal) | Proteintech Group (Manchester, UK) | 13967-1-AP, RRID:AB_2121979 | 1:1000 |
| Ift88 | Rabbit (polyclonal) | Proteintech Group | 17711-1-AP, RRID:AB_2060867 | 1:1000 |
| Islet1 (clone 40.2D6) | Mouse (monoclonal) | DSHB (Iowa City, IA, USA) | 40.2D6, RRID:AB_528315 | 1:30 (supernatant) |
| MelEM | Mouse (monoclonal) | DSHB | MelEM, RRID:AB_531849 | 1:2 (supernatant) |
| P0 (clone 1E8) | Mouse (monoclonal) | DSHB | 1E8, RRID:AB_2078498 | 1:2 (supernatant) |
| HNK1 (clone 1C10) | Mouse (monoclonal) | DSHB | 1C10, RRID:AB_10570406 | 1:2 (supernatant) |
| Axonin-1/Contactin-2 | Goat (polyclonal) | Sonderegger Lab | NA | 1:1000 |
| Tyrosine Hydroxylase | Mouse (monoclonal) | DSHB | aTH, RRID:AB_528490 | 1:5 (supernatant) |
| Neurofilament-M (clone RMO270) | Mouse (monoclonal) | Invitrogen (Thermo Fisher Scientific, Waltham, MA, USA) | RMO270, RRID:AB_2315286 | 1:250 |
| Secondary Antibodies | Source | Cat# | Dilution |
|---|---|---|---|
| Donkey-anti-mouse IgG-Alexa-488 | Invitrogen (Thermor Fisher Scientific, Waltham, MA, USA) | A21202 RRID:AB_141607 | 1:1000 |
| Donkey-anti-rabbit-Cy3 | Jackson ImmunoResearch (West Grove, PA, USA) | 715-165-152 RRID: AB_2307443 | 1:1000 |
| Donkey-anti-goat-Cy5 | Jackson ImmunoResearch | 705-175-147 RRID: AB_2340415 | 1:1000 |
| Donkey-anti-Goat IgG-Alexa-488 | Invitrogen | A11055 RRID:AB_2534102 | 1:1000 |
| Cell Populations | ||||
|---|---|---|---|---|
| DRG | MEM with Glutamax (Invitrogen, cat# 41090-028) | Albumax (4 mg/mL, Invitrogen, cat# 11020-021) | N3 | NGF (20 ng/mL; Invitrogen, cat# 13290-010) |
| Commissural neurons/motor neurons | MEM with Glutamax (Invitrogen, cat# 41090-028) | Albumax (4 mg/mL, Invitrogen, cat# 11020-021) | N3 | Pyruvate (1 mM; Sigma, cat# P5280) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yusifov, E.; Dumoulin, A.; Stoeckli, E.T. Investigating Primary Cilia during Peripheral Nervous System Formation. Int. J. Mol. Sci. 2021, 22, 3176. https://doi.org/10.3390/ijms22063176
Yusifov E, Dumoulin A, Stoeckli ET. Investigating Primary Cilia during Peripheral Nervous System Formation. International Journal of Molecular Sciences. 2021; 22(6):3176. https://doi.org/10.3390/ijms22063176
Chicago/Turabian StyleYusifov, Elkhan, Alexandre Dumoulin, and Esther T. Stoeckli. 2021. "Investigating Primary Cilia during Peripheral Nervous System Formation" International Journal of Molecular Sciences 22, no. 6: 3176. https://doi.org/10.3390/ijms22063176
APA StyleYusifov, E., Dumoulin, A., & Stoeckli, E. T. (2021). Investigating Primary Cilia during Peripheral Nervous System Formation. International Journal of Molecular Sciences, 22(6), 3176. https://doi.org/10.3390/ijms22063176