
The Function of Sialidase Revealed by Sialidase Activity Imaging Probe


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Sialidase Activity Imaging Probes
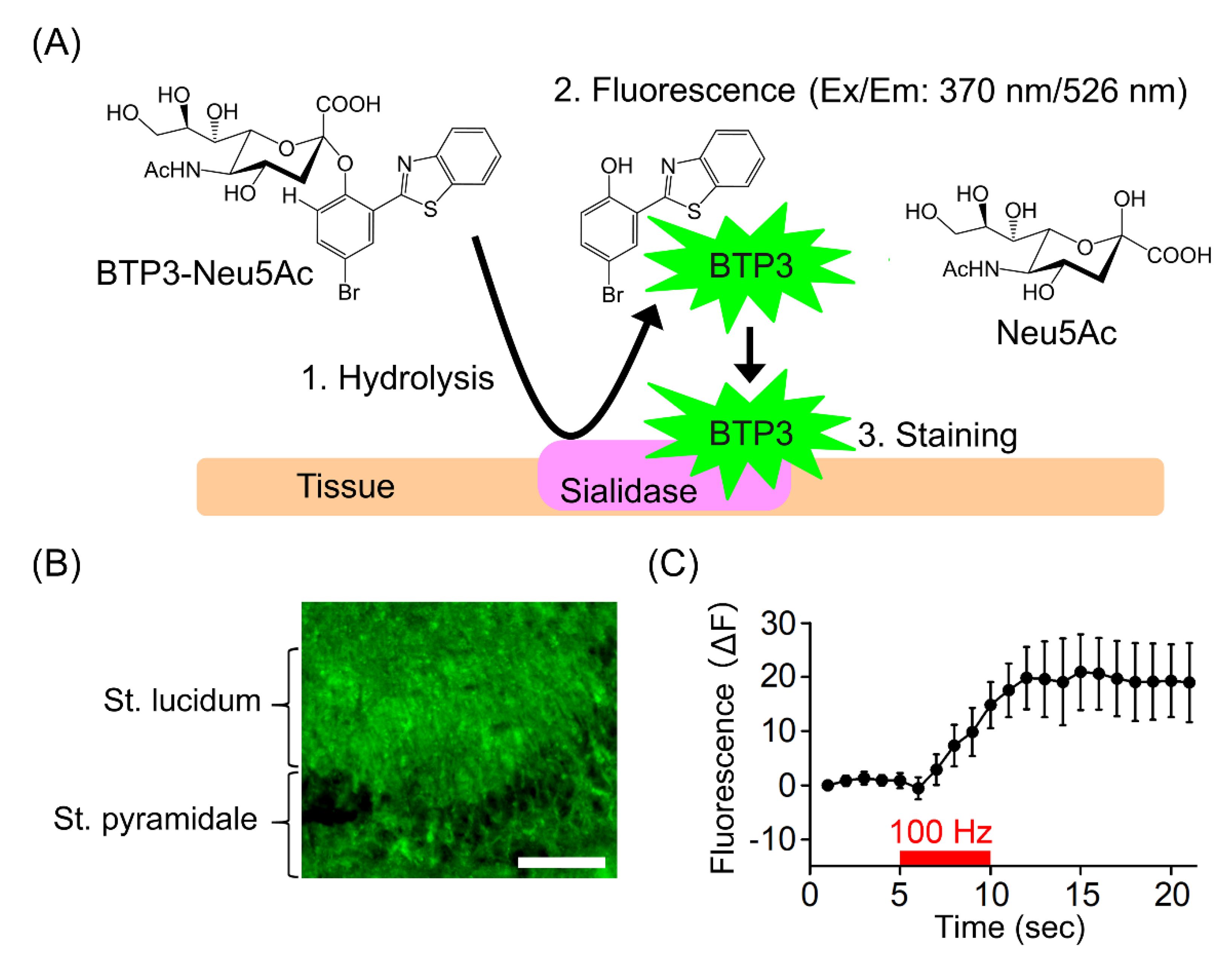
3. Highly Sensitive Imaging Probe for Sialidase Activity
4. Increase in Sialidase Activity in Conjunction with Neural Excitation
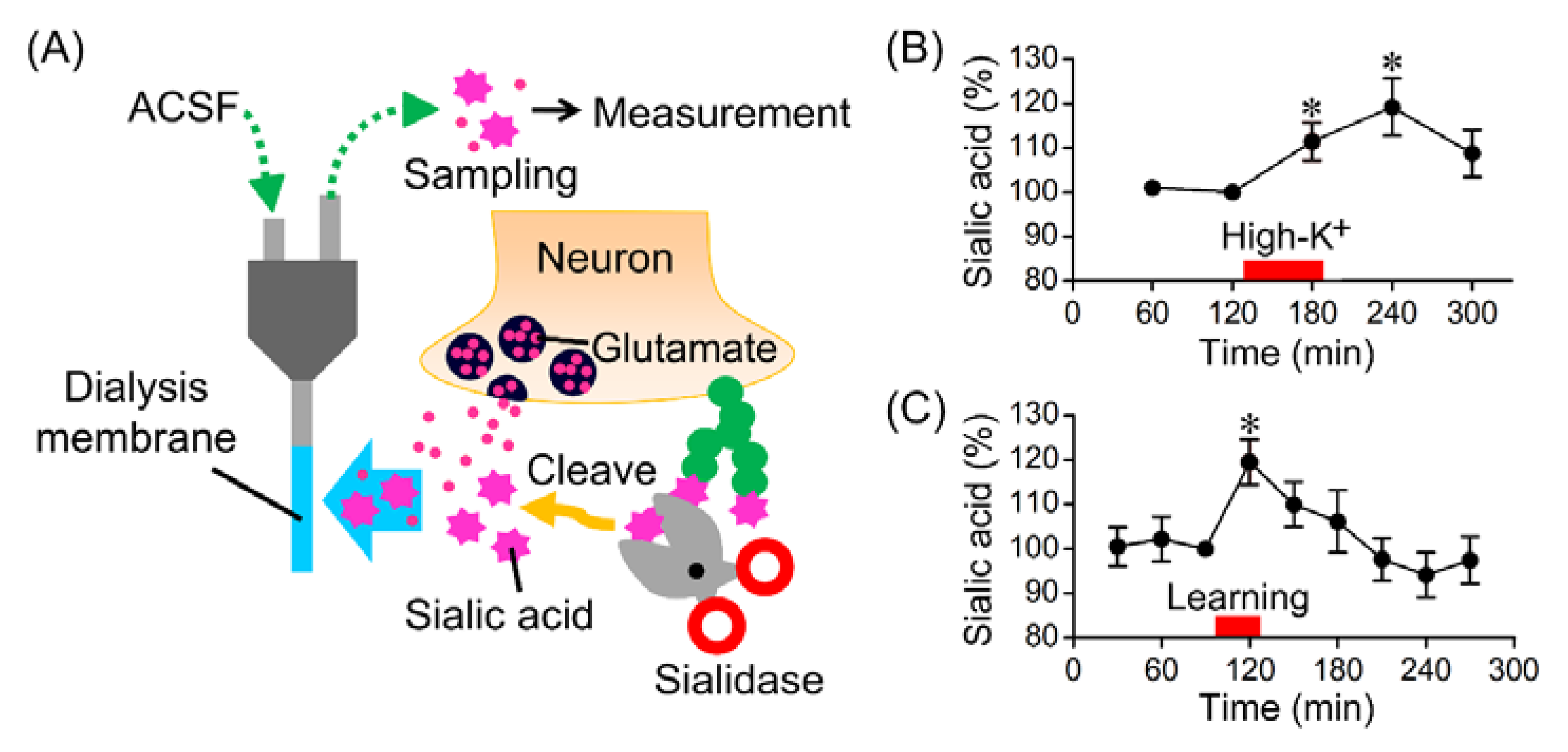
5. In Vivo Sialidase Activity Monitoring
6. Role of Sialidase in Memory Formation
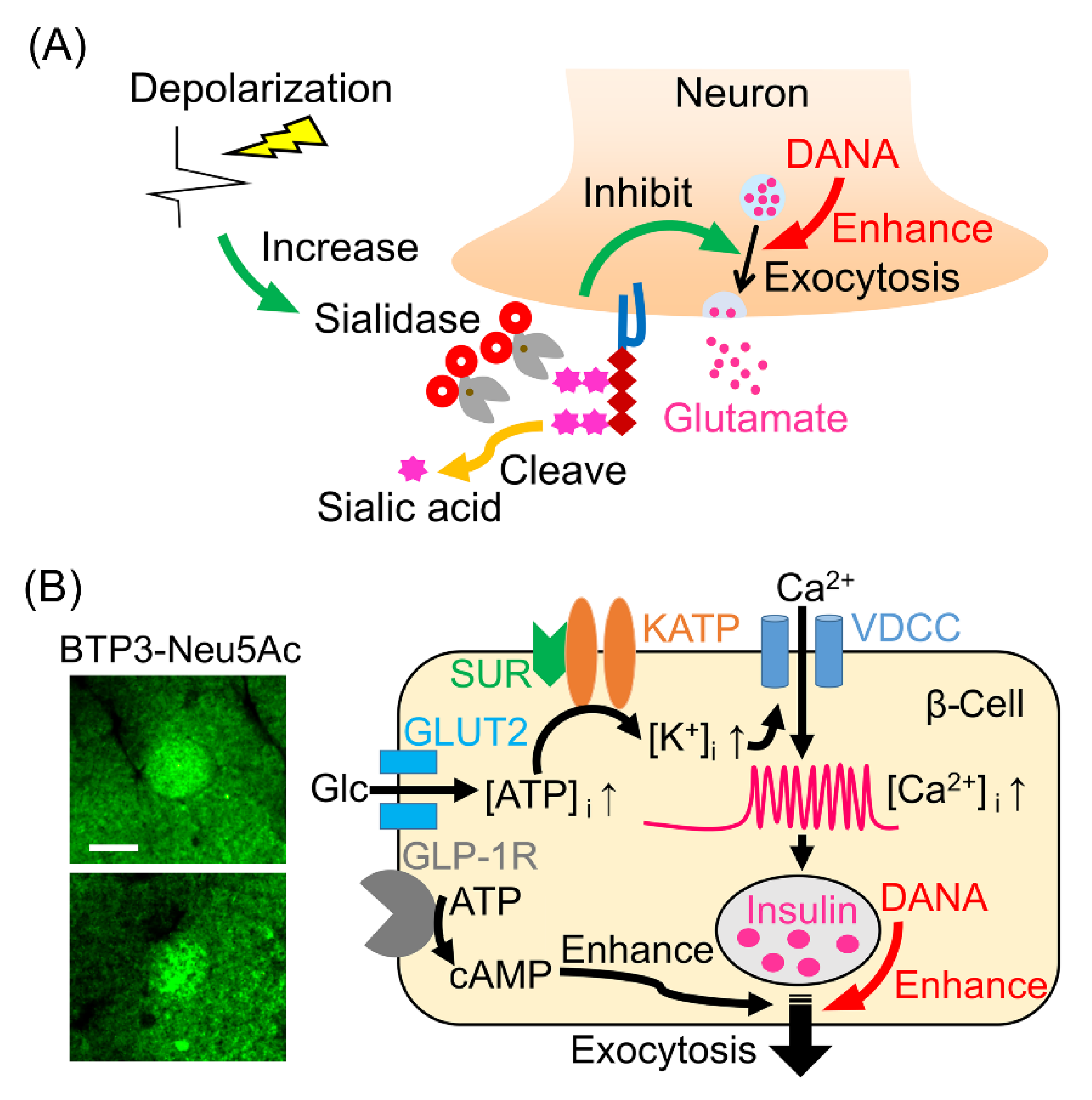
7. Role of Sialidase in the Glutamate Release
8. Role of Sialidase in the Pancreatic Islets
9. Role of Sialidase in the Skin
10. Improvement of Aged Skin by Sialidase
11. Detection of Influenza Viruses
12. Modification of BTP3-Neu5Ac
13. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AMPA | amino-3-hydroxy-5-methyl-4-isoxazolepropionate |
| BDNF | brain-derived neurotrophic factor |
| BTP | 2-(benzo[d]thiazol-2-yl)-4-bromophenol |
| BTP3-Neu5Ac | 2-(benzo[d]thiazol-2-yl)-4-bromophenyl-α-d-N-acetylneuraminic acid |
| BTP9-Neu5Ac | 2-(benzo[d]thiazol-2-yl)-5-(non-1-yn-1-yl)phenyl-α-d-N-acetylneuraminic acid |
| CAGE | choline and geranate |
| DANA | 2:3-dehydro-2-deoxy-N-acetylneuraminic acid |
| DGNA | 2,3-dehydro-2-deoxy-N-glycolylneuraminic acid |
| DPNA | 2,3-dehydro-2-deoxy-N-propanoylneuraminic acid |
| EBP | elastin-binding protein |
| FRV | fast red violet |
| LTP | long-term potentiation |
| NA | viral neuraminidase glycoprotein |
| NAI | influenza virus-specific neuraminidase inhibitor |
| NCAM | neural cell adhesion molecule |
| NMDA | N-methyl-D-aspartate |
| PSA | polysialic acid |
| SAM | senescence-accelerated mouse |
| SAMP | senescence-accelerated mouse prone |
| X | 5-bromo-4-chloro-3-hydroxyindole |
| X-Neu5Ac | 5-bromo-4-chloro-3-hydroxyindole derivative |
| 4MU | 4-methylumbelliferone |
| 4MU-Neu5Ac | 4-methylumbelliferyl-α-d-N-acetylneuraminic acid |
References
- Isaev, D.; Isaeva, E.; Shatskih, T.; Zhao, Q.; Smits, N.C.; Shworak, N.W.; Khazipov, R.; Holmes, G.L. Role of Extracellular Sialic Acid in Regulation of Neuronal and Network Excitability in the Rat Hippocampus. J. Neurosci. 2007, 27, 11587–11594. [Google Scholar] [CrossRef] [PubMed]
- Hammond, M.S.; Sims, C.; Parameshwaran, K.; Suppiramaniam, V.; Schachner, M.; Dityatev, A. Neural cell adhesion molecule-associated polysialic acid inhibits NR2B-containing N-methyl-D-aspartate receptors and prevents glutamate-induced cell death. J. Biol. Chem. 2006, 281, 34859–34869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varbanov, H.; Dityatev, A. Regulation of extrasynaptic signaling by polysialylated NCAM: Impact for synaptic plasticity and cognitive functions. Mol. Cell. Neurosci. 2017, 81, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Murai, R.; Takahashi, Y.; Mori, A.; Hane, M.; Kitajima, K.; Sato, C. Comparative Studies of Polysialic Acids Derived from Five Different Vertebrate Brains. Int. J. Mol. Sci. 2020, 21, 8593. [Google Scholar] [CrossRef]
- Isomura, R.; Kitajima, K.; Sato, C. Structural and Functional Impairments of Polysialic Acid by a Mutated Polysialyltransferase Found in Schizophrenia. J. Biol. Chem. 2011, 286, 21535–21545. [Google Scholar] [CrossRef] [Green Version]
- Di Cristo, G.; Chattopadhyaya, B.; Kuhlman, S.J.; Fu, Y.; Bélanger, M.-C.; Wu, C.Z.; Rutishauser, U.; Maffei, L.; Huang, Z.J. Activity-dependent PSA expression regulates inhibitory maturation and onset of critical period plasticity. Nat. Neurosci. 2007, 10, 1569–1577. [Google Scholar] [CrossRef]
- Saito, M.; Hagita, H.; Iwabuchi, Y.; Fujii, I.; Ikeda, K.; Ito, M. Fluorescent cytochemical detection of sialidase activity using 5-bromo-4-chloroindol-3-yl-α-D-N-acetylneuraminic acid as the substrate. Histochem. Cell Biol. 2002, 117, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Minami, A.; Shimizu, H.; Meguro, Y.; Shibata, N.; Kanazawa, H.; Ikeda, K.; Suzuki, T. Imaging of sialidase activity in rat brain sections by a highly sensitive fluorescent histochemical method. NeuroImage 2011, 58, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Minami, A. Multidimensional Analysis of Hippocampal Excitatory Neurotransmission and Development of Analytical Tools for Glycans. Yakugaku Zasshi 2015, 135, 1341–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minami, A.; Otsubo, T.; Ieno, D.; Ikeda, K.; Kanazawa, H.; Shimizu, K.; Ohata, K.; Yokochi, T.; Horii, Y.; Fukumoto, H.; et al. Visualization of Sialidase Activity in Mammalian Tissues and Cancer Detection with a Novel Fluorescent Sialidase Substrate. PLoS ONE 2014, 9, e81941. [Google Scholar] [CrossRef] [PubMed]
- Minami, A.; Suzuki, T. Development of analytical tools for sialidase functions and its application in brain. Kagaku Seibutsu 2019, 57, 346–351. [Google Scholar] [CrossRef]
- Minami, A.; Saito, M.; Mamada, S.; Ieno, D.; Hikita, T.; Takahashi, T.; Otsubo, T.; Ikeda, K.; Suzuki, T. Role of Sialidase in Long-Term Potentiation at Mossy Fiber-CA3 Synapses and Hippocampus-Dependent Spatial Memory. PLoS ONE 2016, 11, e0165257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas, C.; Kamiya, M.; Urano, Y. A novel sialidase-activatable fluorescence probe with improved stability for the sensitive detection of sialidase. Bioorg. Med. Chem. Lett. 2020, 30, 126860. [Google Scholar] [CrossRef]
- Kurebayashi, Y.; Takahashi, T.; Miura, T.; Otsubo, T.; Minami, A.; Fujita, Y.; Sakakibara, K.; Tanabe, M.; Iuchi, A.; Ota, R.; et al. Fluorogenic Probes for Accurate in Situ Imaging of Viral and Mammalian Sialidases. ACS Chem. Biol. 2019, 14, 1195–1204. [Google Scholar] [CrossRef]
- Minami, A.; Meguro, Y.; Ishibashi, S.; Ishii, A.; Shiratori, M.; Sai, S.; Horii, Y.; Shimizu, H.; Fukumoto, H.; Shimba, S.; et al. Rapid regulation of sialidase activity in response to neural activity and sialic acid removal during memory processing in rat hippocampus. J. Biol. Chem. 2017, 292, 5645–5654. [Google Scholar] [CrossRef] [Green Version]
- Savotchenko, A.; Romanov, A.; Isaev, D.; Maximyuk, O.; Sydorenko, V.; Holmes, G.L.; Isaeva, E. Neuraminidase Inhibition Primes Short-Term Depression and Suppresses Long-Term Potentiation of Synaptic Transmission in the Rat Hippocampus. Neural Plast. 2015, 2015, 1–10. [Google Scholar] [CrossRef]
- Seki, T.; Arai, Y. Different polysialic acid-neural cell adhesion molecule expression patterns in distinct types of mossy fiber boutons in the adult hippocampus. J. Comp. Neurol. 1999, 410, 115–125. [Google Scholar] [CrossRef]
- Minami, A.; Ishii, A.; Shimba, S.; Kano, T.; Fujioka, E.; Sai, S.; Oshio, N.; Ishibashi, S.; Takahashi, T.; Kurebayashi, Y.; et al. Down-regulation of glutamate release from hippocampal neurons by sialidase. J. Biochem. 2018, 163, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Boll, I.; Jensen, P.; Schwämmle, V.; Larsen, M.R. Depolarization-dependent Induction of Site-specific Changes in Sialylation on N-linked Glycoproteins in Rat Nerve Terminals. Mol. Cell. Proteom. 2020, 19, 1418–1435. [Google Scholar] [CrossRef]
- Dridi, L.; Seyrantepe, V.; Fougerat, A.; Pan, X.; Bonneil, É.; Thibault, P.; Moreau, A.; Mitchell, G.A.; Heveker, N.; Cairo, C.W.; et al. Positive Regulation of Insulin Signaling by Neuraminidase 1. Diabetes 2013, 62, 2338–2346. [Google Scholar] [CrossRef] [Green Version]
- Fougerat, A.; Pan, X.; Smutova, V.; Heveker, N.; Cairo, C.W.; Issad, T.; Larrivee, B.; Medin, J.A.; Pshezhetsky, A.V. Neuraminidase 1 activates insulin receptor and reverses insulin resistance in obese mice. Mol. Metab. 2018, 12, 76–88. [Google Scholar] [CrossRef]
- Lipina, C.; Nardi, F.; Grace, H.; Hundal, H.S. NEU3 sialidase as a marker of insulin sensitivity: Regulation by fatty acids. Cell. Signal. 2015, 27, 1742–1750. [Google Scholar] [CrossRef]
- Sasaki, A.; Hata, K.; Suzuki, S.; Sawada, M.; Wada, T.; Yamaguchi, K.; Obinata, M.; Tateno, H.; Suzuki, H.; Miyagi, T. Over-expression of plasma membrane-associated sialidase attenuates insulin signaling in transgenic mice. J. Biol. Chem. 2003, 278, 27896–27902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alghamdi, F.; Guo, M.; Abdulkhalek, S.; Crawford, N.; Amith, S.R.; Szewczuk, M.R. A novel insulin receptor-signaling platform and its link to insulin resistance and type 2 diabetes. Cell. Signal. 2014, 26, 1355–1368. [Google Scholar] [CrossRef] [Green Version]
- Yoshizumi, S.; Suzuki, S.; Hirai, M.; Hinokio, Y.; Yamada, T.; Yamada, T.; Tsunoda, U.; Aburatani, H.; Yamaguchi, K.; Miyagi, T.; et al. Increased hepatic expression of ganglioside-specific sialidase, NEU3, improves insulin sensitivity and glucose tolerance in mice. Metabolism 2007, 56, 420–429. [Google Scholar] [CrossRef]
- Inokuchi, J.-I.; Inamori, K.-I.; Kabayama, K.; Nagafuku, M.; Uemura, S.; Go, S.; Suzuki, A.; Ohno, I.; Kanoh, H.; Shishido, F. Biology of GM3 Ganglioside. Prog. Mol. Biol. Transl. Sci. 2018, 156, 151–195. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Yamaguchi, K.; Wada, T.; Takeda, A.; Itoyama, Y.; Miyagi, T. Molecular cloning of mouse ganglioside sialidase and its increased expression in Neuro2a cell differentiation. J. Biol. Chem. 2000, 275, 8007–8015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyrantepe, V.; Demir, S.A.; Timur, Z.K.; von Gerichten, J.; Marsching, C.; Erdemli, E.; Oztas, E.; Takahashi, K.; Yamaguchi, K.; Ates, N.; et al. Murine Sialidase Neu3 facilitates GM2 degradation and bypass in mouse model of Tay-Sachs disease. Exp. Neurol. 2018, 299, 26–41. [Google Scholar] [CrossRef]
- Blaise, S.; Romier, B.; Kawecki, C.; Ghirardi, M.; Rabenoelina, F.; Baud, S.; Duca, L.; Maurice, P.; Heinz, A.; Schmelzer, C.E.; et al. Elastin-Derived Peptides Are New Regulators of Insulin Resistance Development in Mice. Diabetes 2013, 62, 3807–3816. [Google Scholar] [CrossRef]
- Minami, A.; Fujita, Y.; Shimba, S.; Shiratori, M.; Kaneko, Y.K.; Sawatani, T.; Otsubo, T.; Ikeda, K.; Kanazawa, H.; Mikami, Y.; et al. The sialidase inhibitor 2,3-dehydro-2-deoxy-N-acetylneuraminic acid is a glucose-dependent potentiator of insulin secretion. Sci. Rep. 2020, 10, 5198. [Google Scholar] [CrossRef]
- Hata, K.; Koseki, K.; Yamaguchi, K.; Moriya, S.; Suzuki, Y.; Yingsakmongkon, S.; Hirai, G.; Sodeoka, M.; von Itzstein, M.; Miyagi, T. Limited Inhibitory Effects of Oseltamivir and Zanamivir on Human Sialidases. Antimicrob. Agents Chemother. 2008, 52, 3484–3491. [Google Scholar] [CrossRef] [Green Version]
- Magesh, S.; Moriya, S.; Suzuki, T.; Miyagi, T.; Ishida, H.; Kiso, M. Design, synthesis, and biological evaluation of human sialidase inhibitors. Part 1: Selective inhibitors of lysosomal sialidase (NEU1). Bioorg. Med. Chem. Lett. 2008, 18, 532–537. [Google Scholar] [CrossRef]
- Zhang, Y.; Albohy, A.; Zou, Y.; Smutova, V.; Pshezhetsky, A.V.; Cairo, C.W. Identification of Selective Inhibitors for Human Neuraminidase Isoenzymes Using C4, C7-Modified 2-Deoxy-2,3-didehydro-N-acetylneuraminic Acid (DANA) Analogues. J. Med. Chem. 2013, 56, 2948–2958. [Google Scholar] [CrossRef]
- Albohy, A.; Zhang, Y.; Smutova, V.; Pshezhetsky, A.V.; Cairo, C.W. Identification of Selective Nanomolar Inhibitors of the Human Neuraminidase, NEU4. ACS Med. Chem. Lett. 2013, 4, 532–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.; Héon-Roberts, R.; Zou, C.; Zheng, R.B.; Pshezhetsky, A.V.; Cairo, C.W. Selective Inhibitors of Human Neuraminidase 1 (NEU1). J. Med. Chem. 2018, 61, 11261–11279. [Google Scholar] [CrossRef]
- Hinek, A.; Pshezhetsky, A.V.; von Itzstein, M.; Starcher, B. Lysosomal Sialidase (Neuraminidase-1) Is Targeted to the Cell Surface in a Multiprotein Complex That Facilitates Elastic Fiber Assembly. J. Biol. Chem. 2006, 281, 3698–3710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennasroune, A.; Romier-Crouzet, B.; Blaise, S.; Laffargue, M.; Efremov, R.G.; Martiny, L.; Maurice, P.; Duca, L. Elastic fibers and elastin receptor complex: Neuraminidase-1 takes the center stage. Matrix Biol. 2019, 84, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Minami, A.; Fujita, Y.; Goto, J.; Iuchi, A.; Fujita, K.; Mikami, Y.; Shiratori, M.; Ishii, A.; Mitragotri, S.; Iwao, Y.; et al. Enhancement of elastin expression by transdermal admin-istration of sialidase isozyme Neu2. Sci. Rep. 2021, 11, 3302. [Google Scholar] [CrossRef]
- Koda, T.; Kijimoto-Ochiai, S.; Uemura, S.; Inokuchi, J.-I. Specific expression of Neu2 type B in mouse thymus and the existence of a membrane-bound form in COS cells. Biochem. Biophys. Res. Commun. 2009, 387, 729–735. [Google Scholar] [CrossRef]
- Miyagi, T.; Yamaguchi, K. Mammalian sialidases: Physiological and pathological roles in cellular functions. Glycobiology 2012, 22, 880–896. [Google Scholar] [CrossRef] [Green Version]
- Elias, P.M. Stratum Corneum Defensive Functions: An Integrated View. J. Investig. Dermatol. 2005, 125, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Menon, G.K.; Cleary, G.W.; Lane, M.E. The structure and function of the stratum corneum. Int. J. Pharm. 2012, 435, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Madison, K.C. Barrier function of the skin: “la raison d’etre” of the epidermis. J. Investig. Dermatol. 2003, 121, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakrewsky, M.; Lovejoy, K.S.; Kern, T.L.; Miller, T.E.; Le, V.; Nagy, A.; Goumas, A.M.; Iyer, R.S.; del Sesto, R.E.; Koppisch, A.T.; et al. Ionic liquids as a class of materials for transdermal delivery and pathogen neutralization. Proc. Natl. Acad. Sci. USA 2014, 111, 13313–13318. [Google Scholar] [CrossRef] [Green Version]
- Tanner, E.E.; Ibsen, K.N.; Mitragotri, S. Transdermal insulin delivery using choline-based ionic liquids (CAGE). J. Control. Release 2018, 286, 137–144. [Google Scholar] [CrossRef]
- Banerjee, A.; Ibsen, K.; Iwao, Y.; Zakrewsky, M.; Mitragotri, S. Transdermal Protein Delivery Using Choline and Geranate (CAGE) Deep Eutectic Solvent. Adv. Health Mater. 2017, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Rittie, L.; Fisher, G.J. Natural and Sun-Induced Aging of Human Skin. Cold Spring Harb. Perspect. Med. 2015, 5, a015370. [Google Scholar] [CrossRef]
- El-Domyati, M.; Attia, S.; Saleh, F.; Brown, D.; Birk, D.E.; Gasparro, F.; Ahmad, H.; Uitto, J. Intrinsic aging vs. photoaging: A comparative histopathological, immunohistochemical, and ultrastructural study of skin. Exp. Dermatol. 2002, 11, 398–405. [Google Scholar] [CrossRef]
- Le Page, A.; Khalil, A.; Vermette, P.; Frost, E.H.; Larbi, A.; Witkowski, J.M.; Fulop, T. The role of elastin-derived peptides in human physiology and diseases. Matrix Biol. 2019, 84, 81–96. [Google Scholar] [CrossRef]
- Zhang, S.; Duan, E. Fighting against skin aging: The way from bench to bedside. Cell Transplant. 2018, 27, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Compton, C.C. Current Concepts in Pediatric Burn Care: The Biology of Cultured Epithelial Autografts: An Eight-Year Study in Pediatric Burn Patients. Eur. J. Pediatr. Surg. 1992, 2, 216–222. [Google Scholar] [CrossRef]
- Raghunath, M.; Bächi, T.; Meuli, M.; Altermatt, S.; Gobet, R.; Bruckner-Tuderman, L.; Steinmann, B. Fibrillin and elastin ex-pression in skin regenerating from cultured keratinocyte autografts: Morphogenesis of microfibrils begins at the dermo-epidermal junction and precedes elastic fiber formation. J. Investig. Dermatol. 1996, 106, 1090–1095. [Google Scholar] [CrossRef] [Green Version]
- Chiba, Y.; Shimada, A.; Kumagai, N.; Yoshikawa, K.; Ishii, S.; Furukawa, A.; Takei, S.; Sakura, M.; Kawamura, N.; Hosokawa, M. The Senescence-accelerated Mouse (SAM): A Higher Oxidative Stress and Age-dependent Degenerative Diseases Model. Neurochem. Res. 2008, 34, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T.; Hosokawa, M.; Higuchi, K. Senescence-accelerated mouse (SAM): A novel murine model of senescence. Exp. Ger-ontol. 1997, 32, 105–109. [Google Scholar] [CrossRef]
- Takeda, T.; Hosokawa, M.; Takeshita, S.; Irino, M.; Higuchi, K.; Matsushita, T.; Tomita, Y.; Yasuhira, K.; Hamamoto, H.; Shimizu, K. A new murine model of accelerated senescence. Mech. Ageing Dev. 1981, 17, 183–194. [Google Scholar] [CrossRef]
- Sakura, M.; Chiba, Y.; Kamiya, E.; Furukawa, A.; Kawamura, N.; Niwa, M.; Takeuchi, M.; Hosokawa, M. Spontaneous occurrence of photoageing-like phenotypes in the dorsal skin of old SAMP1 mice, an oxidative stress model. Exp. Dermatol. 2013, 22, 62–64. [Google Scholar] [CrossRef]
- Changarathil, G.; Ramirez, K.; Isoda, H.; Sada, A.; Yanagisawa, H. Wild-type and SAMP8 mice show age-dependent changes in distinct stem cell compartments of the interfollicular epidermis. PLoS ONE 2019, 14, e0215908. [Google Scholar] [CrossRef]
- Todo, H. Transdermal Permeation of Drugs in Various Animal Species. Pharmaceutics 2017, 9, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fornieri, C.; Quaglino, D.; Lungarella, G.; Cavarra, E.; Tiozzo, R.; Giro, M.G.; Canciani, M.; Davidson, J.M.; Ronchetti, I.P. Elastin Production and Degradation in Cutis Laxa Acquisita. J. Investig. Dermatol. 1994, 103, 583–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truter, S.; Rosenbaum-Fiedler, J.; Sapadin, A.; Lebwohl, M. Calcification of elastic fibers in pseudoxanthoma elasticum. Mt. Sinai J. Med. J. Transl. Pers. Med. 1996, 63, 210–215. [Google Scholar]
- Lasio, M.L.D.; Kozel, B.A. Elastin-driven genetic diseases. Matrix Biol. 2018, 71–72, 144–160. [Google Scholar] [CrossRef]
- Ezure, T.; Amano, S. Increment of subcutaneous adipose tissue is associated with decrease of elastic fibres in the dermal layer. Exp. Dermatol. 2015, 24, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Kobasa, D.; Wells, K.; Kawaoka, Y. Amino acids responsible for the absolute sialidase activity of the influenza A virus neu-raminidase: Relationship to growth in the duck intestine. J. Virol. 2001, 75, 11773–11780. [Google Scholar] [CrossRef] [Green Version]
- Nayak, D.P.; Reichl, U. Neuraminidase activity assays for monitoring MDCK cell culture derived influenza virus. J. Virol. Methods 2004, 122, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Potier, M.; Mameli, L.; Bélisle, M.; Dallaire, L.; Melançon, S. Fluorometric assay of neuraminidase with a sodium (4-methylumbelliferyl-α-d-N-acetylneuraminate) substrate. Anal. Biochem. 1979, 94, 287–296. [Google Scholar] [CrossRef]
- Marathe, B.M.; Lévêque, V.; Klumpp, K.; Webster, R.G.; Govorkova, E.A. Determination of Neuraminidase Kinetic Constants Using Whole Influenza Virus Preparations and Correction for Spectroscopic Interference by a Fluorogenic Substrate. PLoS ONE 2013, 8, e71401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buxton, R.C.; Edwards, B.; Juo, R.R.; Voyta, J.C.; Tisdale, M.; Bethell, R.C. Development of a Sensitive Chemiluminescent Neuraminidase Assay for the Determination of Influenza Virus Susceptibility to Zanamivir. Anal. Biochem. 2000, 280, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Kurebayashi, Y.; Takahashi, T.; Otsubo, T.; Ikeda, K.; Takahashi, S.; Takano, M.; Agarikuchi, T.; Sato, T.; Matsuda, Y.; Minami, A.; et al. Imaging of influenza virus sialidase activity in living cells. Sci. Rep. 2014, 4, 4877. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Takano, M.; Kurebayashi, Y.; Agarikuchi, T.; Suzuki, C.; Fukushima, K.; Takahashi, S.; Otsubo, T.; Ikeda, K.; Minami, A.; et al. Rapid Fluorescent Detection Assay for Human Parainfluenza Viruses. Biol. Pharm. Bull. 2015, 38, 1214–1219. [Google Scholar] [CrossRef]
- Takahashi, T.; Takano, M.; Agarikuchi, T.; Kurebayashi, Y.; Minami, A.; Otsubo, T.; Ikeda, K.; Suzuki, T. A novel method for detection of Newcastle disease virus with a fluorescent sialidase substrate. J. Virol. Methods 2014, 209, 136–142. [Google Scholar] [CrossRef]
- Takano, M.; Takahashi, T.; Agarikuchi, T.; Kurebayashi, Y.; Minami, A.; Otsubo, T.; Ikeda, K.; Kanazawa, H.; Suzuki, T. His-tochemical fluorescent staining of Sendai virus-infected cells with a novel sialidase substrate. Virology 2014, 465, 206–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, T.; Agarikuchi, T.; Kurebayashi, Y.; Shibahara, N.; Suzuki, C.; Kishikawa, A.; Fukushima, K.; Takano, M.; Suzuki, F.; Wada, H.; et al. Easy and rapid detection of mumps virus by live fluorescent visualization of virus-Infected cells. PLoS ONE 2015, 10, e0144038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurebayashi, Y.; Takahashi, T.; Tamoto, C.; Sahara, K.; Otsubo, T.; Yokozawa, T.; Shibahara, N.; Wada, H.; Minami, A.; Ikeda, K.; et al. High-Efficiency Capture of Drug Resistant-Influenza Virus by Live Imaging of Sialidase Activity. PLoS ONE 2016, 11, e0156400. [Google Scholar] [CrossRef] [PubMed]
- Kato, D.; Kurebayashi, Y.; Takahashi, T.; Otsubo, T.; Otake, H.; Yamazaki, M.; Tamoto, C.; Minami, A.; Ikeda, K.; Suzuki, T. An easy, rapid, and sensitive method for detection of drug-resistant influenza virus by using a sialidase fluorescent imaging probe, BTP3-Neu5Ac. PLoS ONE 2018, 13, e0200761. [Google Scholar] [CrossRef] [PubMed]
- McAuley, J.L.; Gilbertson, B.P.; Trifkovic, S.; Brown, L.E.; McKimm-Breschkin, J.L. Influenza Virus Neuraminidase Structure and Functions. Front. Microbiol. 2019, 10, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsubo, T.; Minami, A.; Fujii, H.; Taguchi, R.; Takahashi, T.; Suzuki, T.; Teraoka, F.; Ikeda, K. 2-(Benzothiazol-2-yl)-phenyl-beta-d-galactopyranoside derivatives as fluorescent pigment dyeing substrates and their application for the assay of beta-d-galactosidase activities. Bioorg. Med. Chem. Lett. 2013, 23, 2245–2249. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minami, A.; Kurebayashi, Y.; Takahashi, T.; Otsubo, T.; Ikeda, K.; Suzuki, T. The Function of Sialidase Revealed by Sialidase Activity Imaging Probe. Int. J. Mol. Sci. 2021, 22, 3187. https://doi.org/10.3390/ijms22063187
Minami A, Kurebayashi Y, Takahashi T, Otsubo T, Ikeda K, Suzuki T. The Function of Sialidase Revealed by Sialidase Activity Imaging Probe. International Journal of Molecular Sciences. 2021; 22(6):3187. https://doi.org/10.3390/ijms22063187
Chicago/Turabian StyleMinami, Akira, Yuuki Kurebayashi, Tadanobu Takahashi, Tadamune Otsubo, Kiyoshi Ikeda, and Takashi Suzuki. 2021. "The Function of Sialidase Revealed by Sialidase Activity Imaging Probe" International Journal of Molecular Sciences 22, no. 6: 3187. https://doi.org/10.3390/ijms22063187
APA StyleMinami, A., Kurebayashi, Y., Takahashi, T., Otsubo, T., Ikeda, K., & Suzuki, T. (2021). The Function of Sialidase Revealed by Sialidase Activity Imaging Probe. International Journal of Molecular Sciences, 22(6), 3187. https://doi.org/10.3390/ijms22063187