
Long Non-Coding RNA CRYBG3 Promotes Lung Cancer Metastasis via Activating the eEF1A1/MDM2/MTBP Axis


 , and
, and 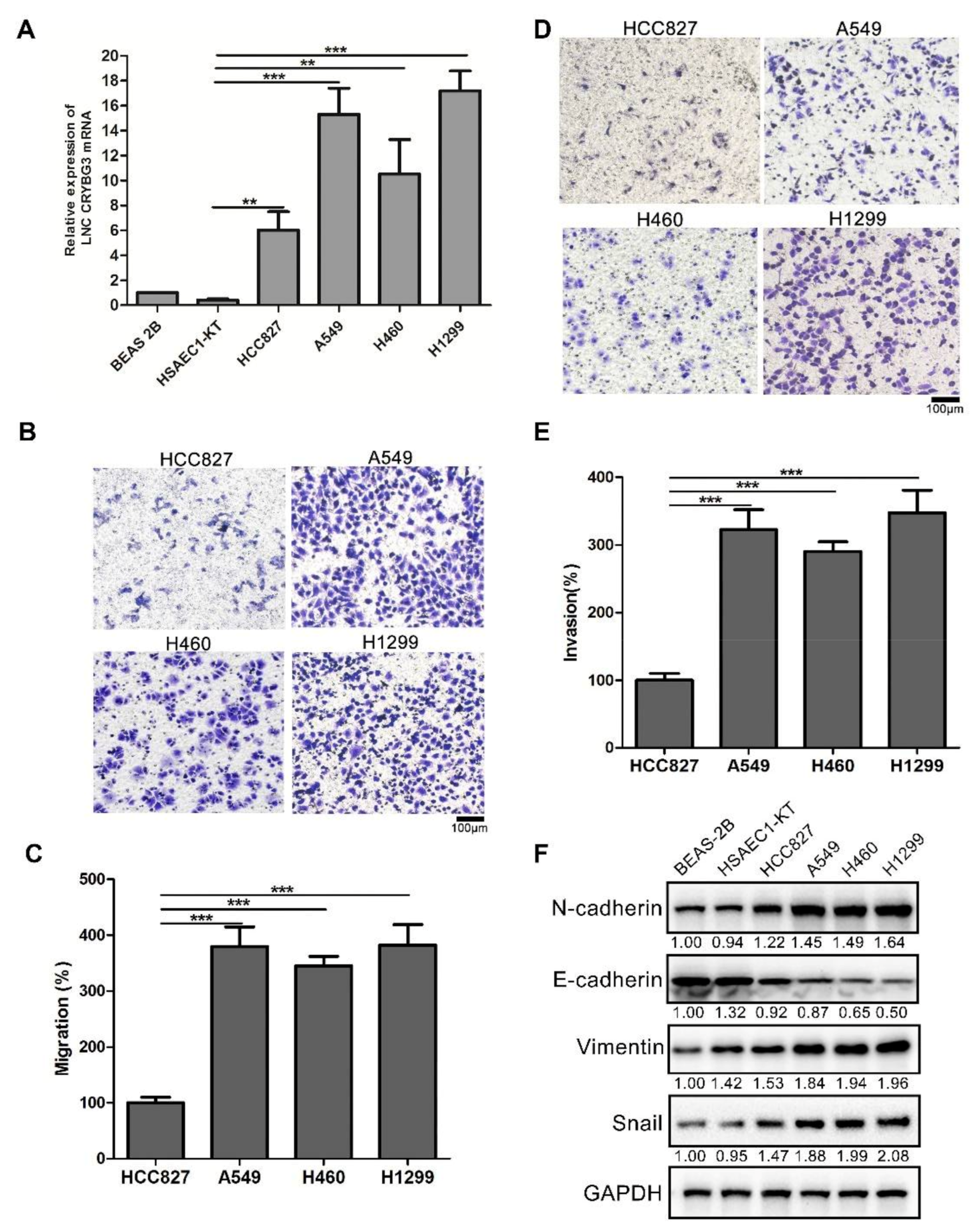{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. LNC CRYBG3 Is Upregulated in NSCLC Cells
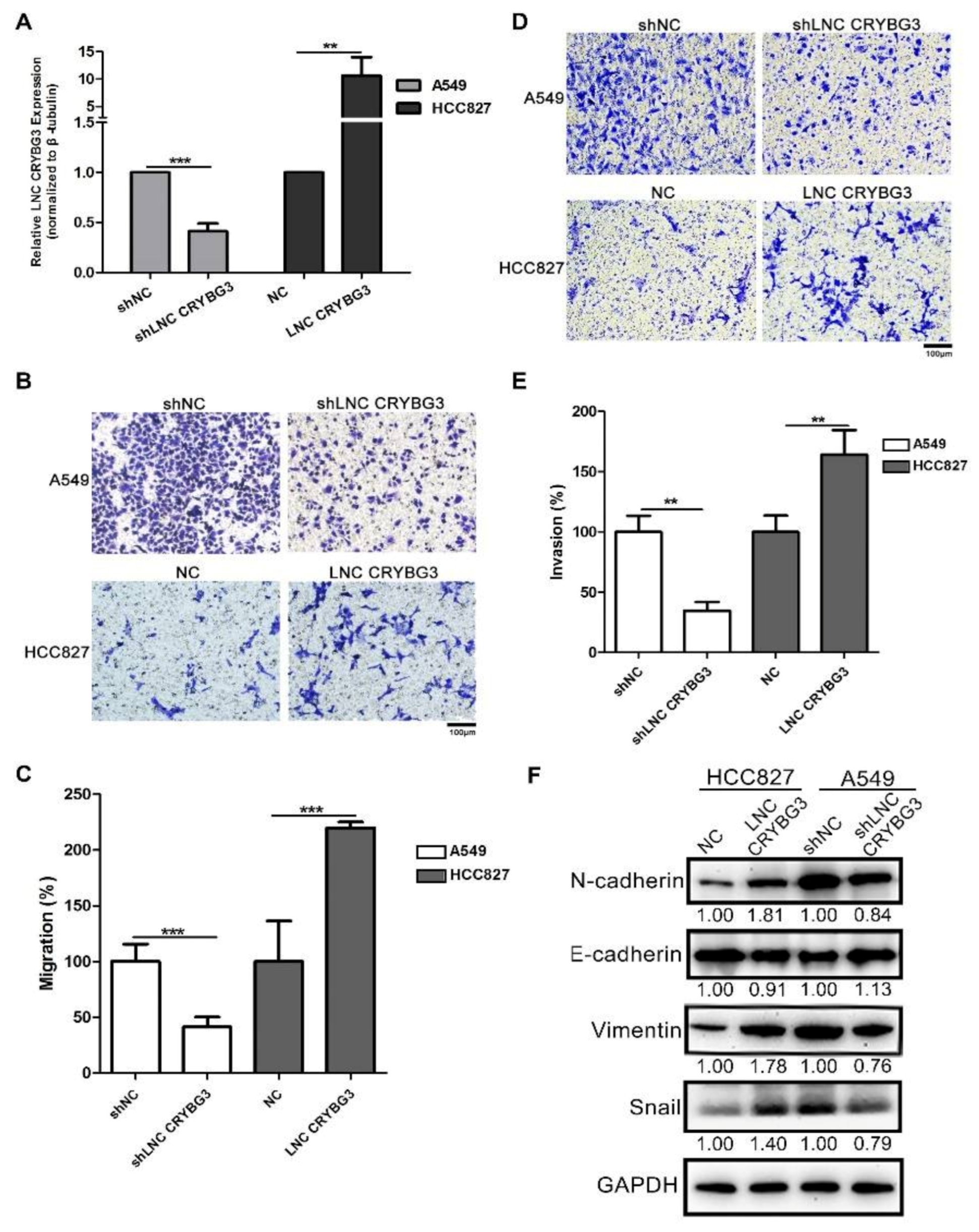
2.2. LNC CRYBG3 Promotes NSCLC Cell Metastasis
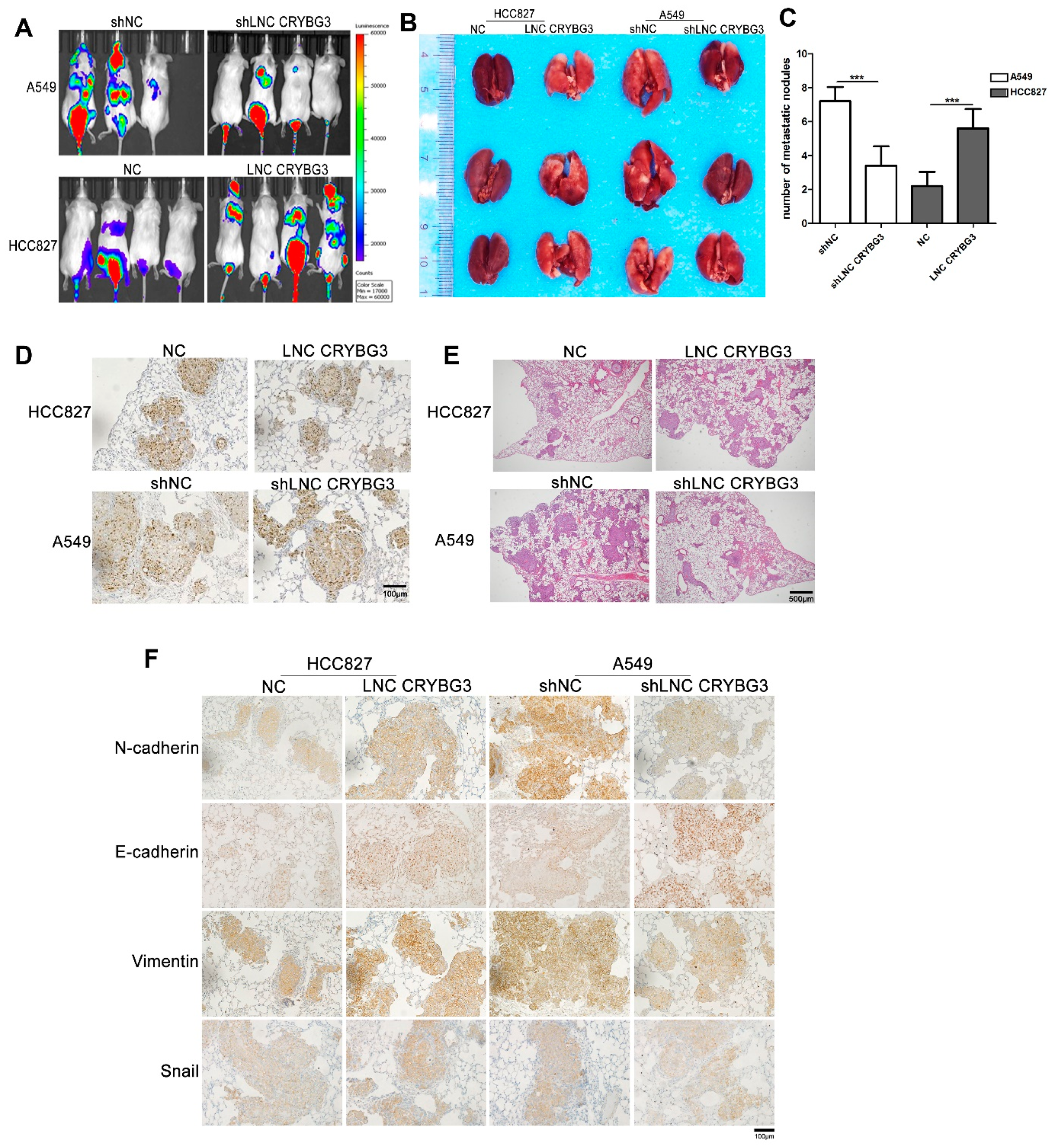
2.3. LNC CRYBG3 Enhances NSCLC Cell Metastatic Ability in NOD/SCID Mice
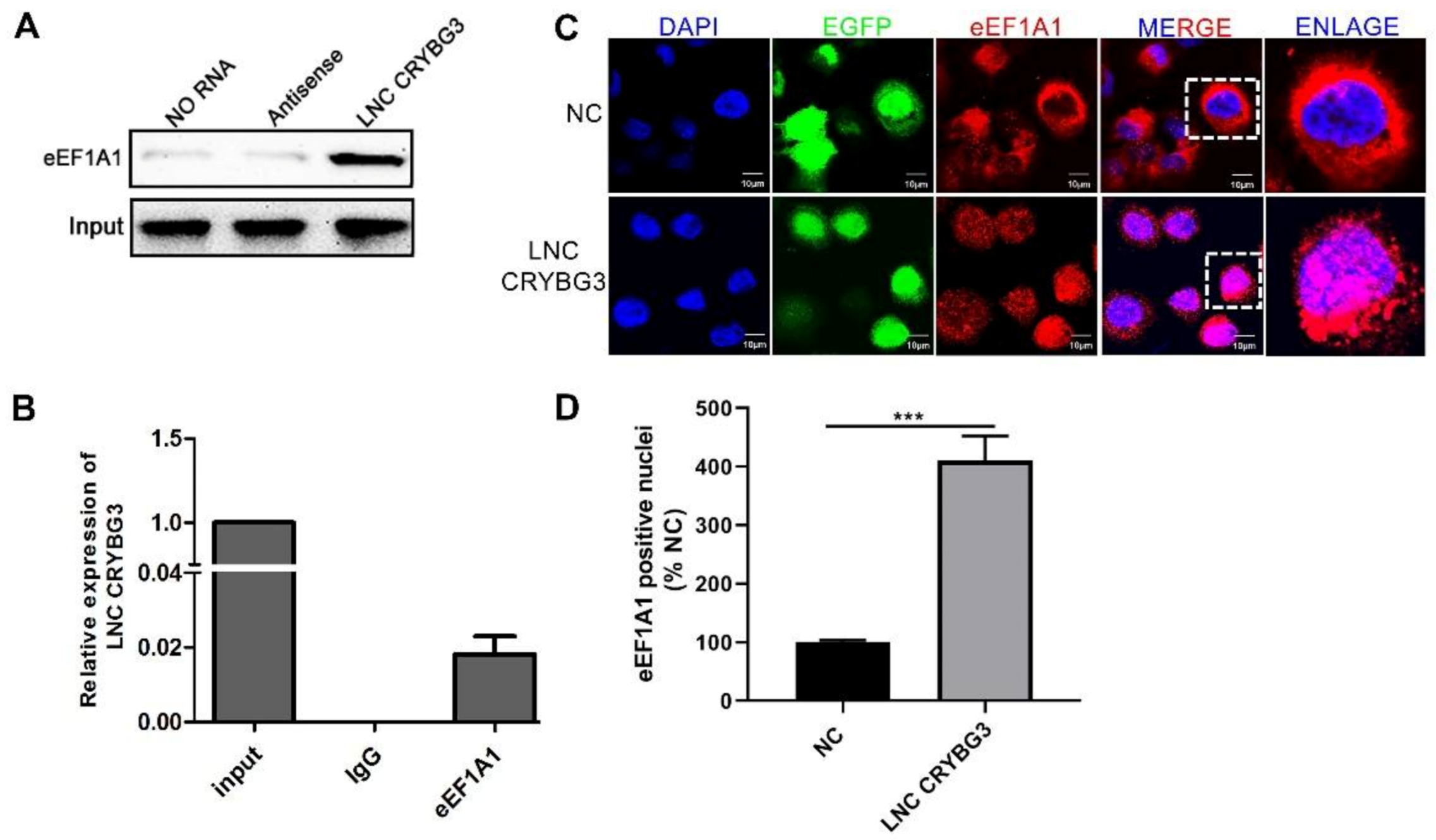
2.4. LNC CRYBG3 Combines with eEF1A1 and Increases Its Nucleus Translocation
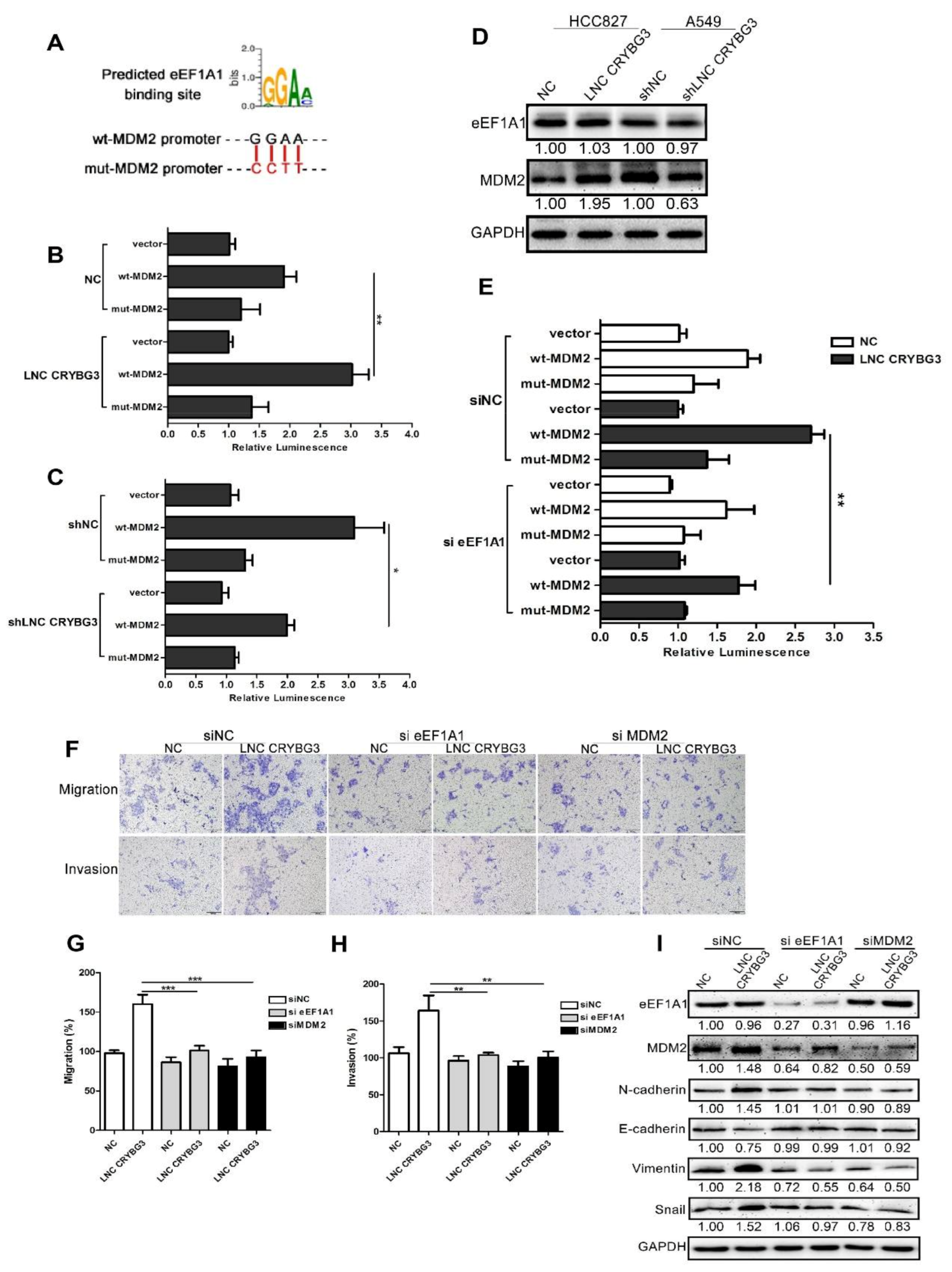
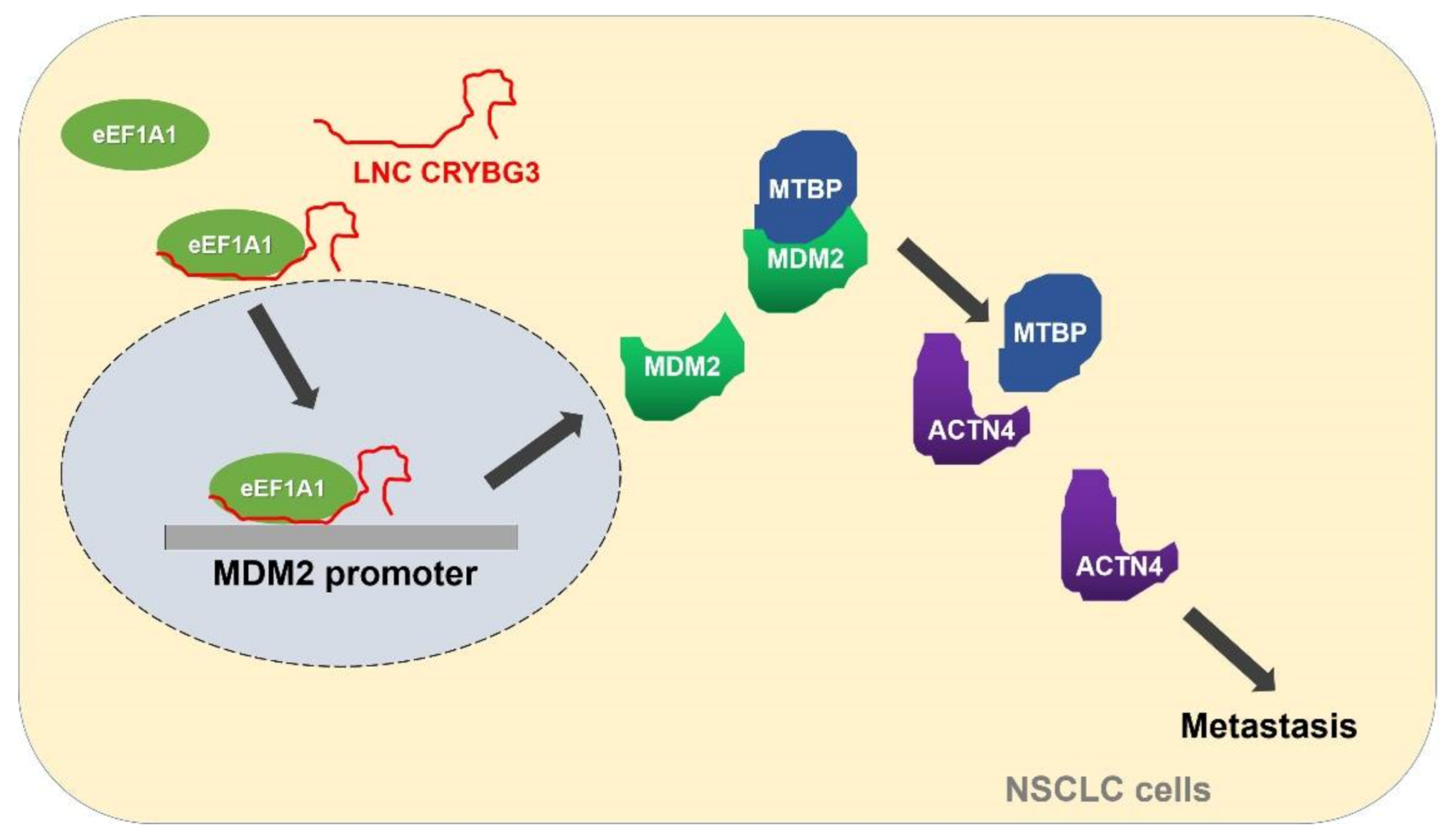
2.5. LNC CRYBG3 Mediates the Binding of eEF1A1 to the Promoter of MDM2 Gene
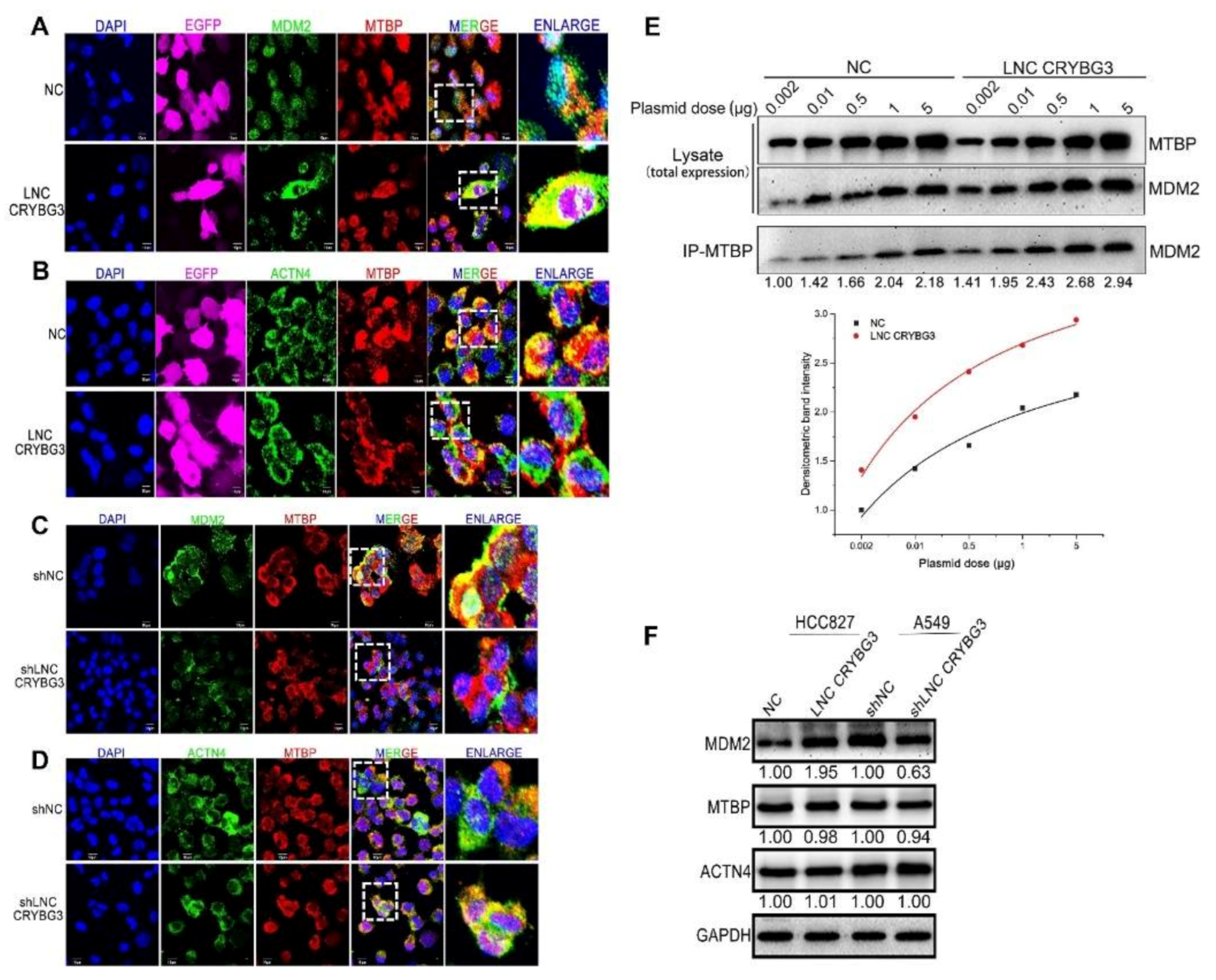
2.6. MDM2 Promotes Metastasis by Blocking the MTBP and ACTN4 Interaction
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. RNA Isolation and Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.3. Gene Silencing Overexpression and Reporter Plasmids
4.4. Transwell Assay
4.5. Animal Experiment
4.6. Western Blot Assay
4.7. siRNA Transfection
4.8. Co-Immunoprecipitation, RNA Pull Down and Immunoprecipitation Analysis, Mass Spectrometry
4.9. Immunofluorescence
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NSCLC | Non-small cell lung cancer |
| lncRNA | Long non-coding RNA |
| siRNA | Small interfering RNA |
| MTBP | MDM2 binding protein |
| EMT | Epithelial-mesenchymal transition |
| MDM2 | Murine double minute 2 |
| eEF1A1 | Eukaryotic translation elongation factor 1 alpha 1 |
References
- Travis, W. The 2015 WHO classification of lung tumors. Der Pathol. 2014, 35 (Suppl. S2), 188. [Google Scholar] [CrossRef]
- Satoh, H.; Miyazaki, K.; Shiozawa, T.; Yamada, H.; Tamura, T. Pattern of bone metastasis in patients with nonsmall cell lung cancer. Ann. Thorac. Med. 2018, 13, 257. [Google Scholar] [CrossRef] [PubMed]
- Marchese, F.P.; Raimondi, I.; Huarte, M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017, 18, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Cao, D.; Wu, F. Long noncoding RNA UPAT promoted cell proliferation via increasing UHRF1 expression in non-small cell lung cancer. Oncol. Lett. 2018, 16, 1491–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Fang, Q.; Meng, S. Knockdown of long noncoding RNA ENST457720 inhibits proliferation of non-small cell lung cancer cells in vitro and in vivo. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2018, 27, 47–53. [Google Scholar] [CrossRef]
- Xia, H.; Jing, H.; Li, Y.; Lv, X. Long noncoding RNA HOXD-AS1 promotes non-small cell lung cancer migration and invasion through regulating miR-133b/MMP9 axis. Biomed. Pharmacother. 2018, 106, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Xiao, G.; Chen, Y.; Deng, Y. LncRNA MALAT1 promotes migration and invasion of non-small-cell lung cancer by targeting miR-206 and activating Akt/mTOR signaling. Anti-Cancer Drugs 2018, 29, 725–735. [Google Scholar] [CrossRef]
- Guan, X.; Xu, Y.; Zheng, J. Long noncoding RNA PCAT6 promotes the development of osteosarcoma by increasing MDM2 expression. Oncol. Rep. 2020, 44, 2465–2474. [Google Scholar] [CrossRef]
- Manfredi, J.J. The Mdm2-p53 relationship evolves: Mdm2 swings both ways as an oncogene and a tumor suppressor. Genes Dev. 2010, 24, 1580–1589. [Google Scholar] [CrossRef] [Green Version]
- Bohlman, S.; Manfredi, J.J. p53-Independent effects of Mdm2. Subcell. Biochem. 2014, 85, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.-Y.; Zong, C.S.; Xia, W.; Wei, Y.; Ali-Seyed, M.; Li, Z.; Broglio, K.; Berry, D.A.; Hung, M.-C. MDM2 promotes cell motility and invasiveness by regulating E-cadherin degradation. Mol. Cell. Biol. 2006, 26, 7269–7282. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Gu, L.; He, J.; Zhang, H.; Zhou, M. MDM2 regulates vascular endothelial growth factor mRNA stabilization in hypoxia. Mol. Cell. Biol. 2011, 31, 4928–4937. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, N.; Adhikari, A.S.; Iyer, S.V.; Hekmatdoost, K.; Welch, D.R.; Iwakuma, T. MTBP suppresses cell migration and filopodia formation by inhibiting ACTN4. Oncogene 2012, 32, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Iwakuma, T.; Agarwal, N. MDM2 binding protein, a novel metastasis suppressor. Cancer Metastasis Rev. 2012, 31, 633–640. [Google Scholar] [CrossRef]
- Chen, H.; Pei, H.; Hu, W.; Ma, J.; Zhang, J.; Mao, W.; Nie, J.; Xu, C.; Li, B.; Hei, T.K.; et al. Long non-coding RNA CRYBG3 regulates glycolysis of lung cancer cells by interacting with lactate dehydrogenase A. J. Cancer 2018, 9, 2580–2588. [Google Scholar] [CrossRef] [PubMed]
- Abbas, W.; Kumar, A.; Herbein, G. The eEF1A proteins: At the crossroads of oncogenesis, apoptosis, and viral infections. Front. Oncol. 2015, 5, 75. [Google Scholar] [CrossRef] [Green Version]
- Vera, M.; Pani, B.; Griffiths, L.A.; Muchardt, C.; Abbott, C.M.; Singer, R.H.; Nudler, E. The translation elongation factor eEF1A1 couples transcription to translation during heat shock response. eLife 2014, 3, e03164. [Google Scholar] [CrossRef]
- Heery, R.; Finn, S.P.; Cuffe, S.; Gray, S.G. Long non-coding RNAs: Key regulators of epithelial-mesenchymal transition, tumour drug resistance and cancer stem cells. Cancers 2017, 9, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, Z.I.; Tam, S.Y.; Law, H.K.W. Autophagy-modulating long non-coding RNAs (LncRNAs) and their molecular events in cancer. Front. Genet. 2019, 9, 750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, P.-F.; Zheng, X.; Fan, X.; Lin, A.-F. Role of cytoplasmic lncRNAs in regulating cancer signaling pathways. J. Zhejiang Univ. Sci. B 2019, 20, 1–8. [Google Scholar] [CrossRef]
- Naderi-Meshkin, H.; Lai, X.; Amirkhah, R.; Vera, J.; Rasko, J.E.J.; Schmitz, U. Exosomal lncRNAs and cancer: Connecting the missing links. Bioinformatics 2019, 35, 352–360. [Google Scholar] [CrossRef]
- Qi, D.; Wang, Q.; Wu, M.; Zhang, X. Comprehensive bioinformatics analysis of lncRNAs in gastric cancer. Oncol. Lett. 2018, 17, 1279–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Zhang, Z.; Xiong, W.; Li, N.; Liu, H.; He, H.; Li, Q.; Liu, Y.; Zhang, L. Estradiol promotes EMT in endometriosis via MA-LAT1/miR200s sponge function. Reproduction 2019, 157, 179–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Liang, L.; Ouyang, K.; Li, Z.; Yi, X. MALAT1 induces tongue cancer cells’ EMT and inhibits apoptosis through Wnt/beta-catenin signaling pathway. J. Oral Pathol. Med. 2017, 46, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Song, L.; Yao, H.; Zhang, L.; Xu, D.; Gao, F.; Lili, S. MiR-375 is epigenetically downregulated by HPV-16 E6 mediated DNMT1 upregulation and modulates EMT of cervical cancer cells by suppressing lncRNA MALAT1. PLoS ONE 2016, 11, e0163460. [Google Scholar] [CrossRef] [Green Version]
- Shi, B.; Wang, Y.; Yin, F. MALAT1/miR-124/Capn4 axis regulates proliferation, invasion and EMT in nasopharyngeal carcinoma cells. Cancer Biol. Ther. 2017, 18, 792–800. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Madura, K. Increased proteasome activity, ubiquitin-conjugating enzymes, and eEF1A translation factor detected in breast cancer tissue. Cancer Res. 2005, 65, 5599–5606. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Namkung, W.; Yoon, J.S.; Jo, M.J.; Lee, S.H.; Kim, K.H.; Kim, J.Y.; Lee, M.G. The role of translation elongation factor eEF1A in intracellular alkalinization-induced tumor cell growth. Lab. Investig. 2009, 89, 867–874. [Google Scholar] [CrossRef] [Green Version]
- Hauck, P.M.; Wolf, E.R.; Olivos, D.J., III; Batuello, C.N.; McElyea, K.C.; McAtarsney, C.P.; Cournoyer, R.M.; Sandusky, G.E.; Mayo, L.D. Early-stage metastasis requires Mdm2 and Not p53 gain of function. Mol. Cancer Res. 2017, 15, 1598–1607. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhou, Y.; Li, B.; Chen, F.; Shen, W.; Lu, Y.; Zhong, C.; Zhang, C.; Xie, H.; Katanaev, V.L.; et al. WDR74 modulates melanoma tumorigenesis and metastasis through the RPL5–MDM2–p53 pathway. Oncogene 2020, 39, 2741–2755. [Google Scholar] [CrossRef]
- Bai, J.; Wu, K.; Cao, M.H.; Yang, Y.; Pan, Y.; Liu, H.; He, Y.; Itahana, Y.; Huang, L.; Zheng, J.N.; et al. SCF(FBXO22) targets HDM2 for degradation and modulates breast cancer cell invasion and metastasis. Proc. Natl. Acad. Sci. USA 2019, 116, 11754–11763. [Google Scholar] [PubMed] [Green Version]
- Boyd, M.T.; Vlatkovic, N.; Haines, D.S. A novel cellular protein (MTBP) binds to MDM2 and induces a G1 arrest that Is suppressed by MDM2. J. Biol. Chem. 2000, 275, 31883–31890. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, A.; Tang, J.; Guo, Z.; Dai, Y.; Nie, J.; Hu, W.; Liu, N.; Ye, C.; Li, S.; Pei, H.; et al. Long Non-Coding RNA CRYBG3 Promotes Lung Cancer Metastasis via Activating the eEF1A1/MDM2/MTBP Axis. Int. J. Mol. Sci. 2021, 22, 3211. https://doi.org/10.3390/ijms22063211
Wu A, Tang J, Guo Z, Dai Y, Nie J, Hu W, Liu N, Ye C, Li S, Pei H, et al. Long Non-Coding RNA CRYBG3 Promotes Lung Cancer Metastasis via Activating the eEF1A1/MDM2/MTBP Axis. International Journal of Molecular Sciences. 2021; 22(6):3211. https://doi.org/10.3390/ijms22063211
Chicago/Turabian StyleWu, Anqing, Jiaxin Tang, Ziyang Guo, Yingchu Dai, Jing Nie, Wentao Hu, Ningang Liu, Caiyong Ye, Shihong Li, Hailong Pei, and et al. 2021. "Long Non-Coding RNA CRYBG3 Promotes Lung Cancer Metastasis via Activating the eEF1A1/MDM2/MTBP Axis" International Journal of Molecular Sciences 22, no. 6: 3211. https://doi.org/10.3390/ijms22063211
APA StyleWu, A., Tang, J., Guo, Z., Dai, Y., Nie, J., Hu, W., Liu, N., Ye, C., Li, S., Pei, H., & Zhou, G. (2021). Long Non-Coding RNA CRYBG3 Promotes Lung Cancer Metastasis via Activating the eEF1A1/MDM2/MTBP Axis. International Journal of Molecular Sciences, 22(6), 3211. https://doi.org/10.3390/ijms22063211