
Crystal Structure of a Phospholipase D from the Plant-Associated Bacteria Serratia plymuthica Strain AS9 Reveals a Unique Arrangement of Catalytic Pocket


Abstract
:1. Introduction
2. Results
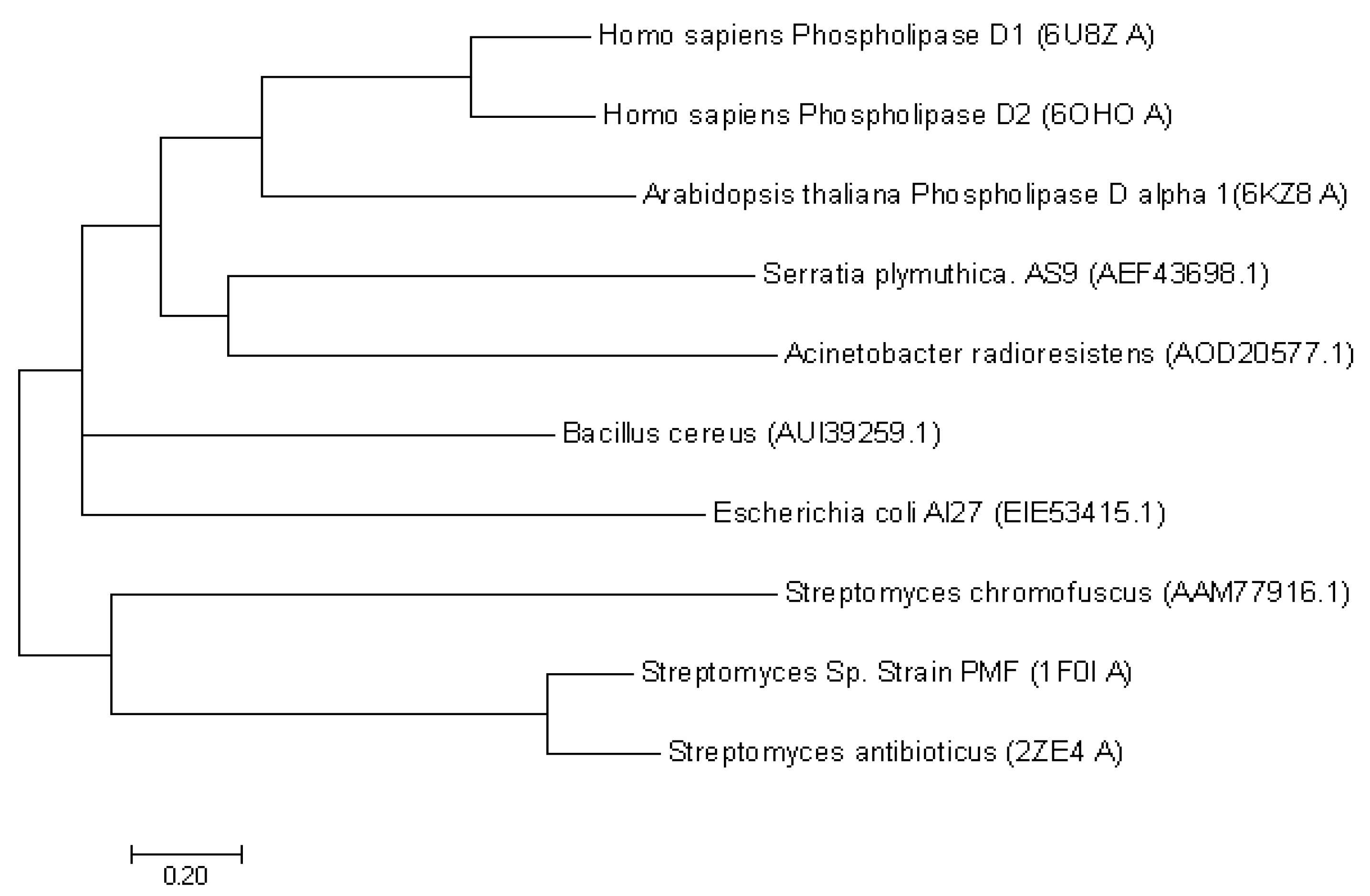
2.1. Bioinformatic Analysis
2.2. Expression and Purification of the Mature SpPLD
2.3. X-ray Diffraction Results
2.4. Overall Structure
2.5. Structural Comparison
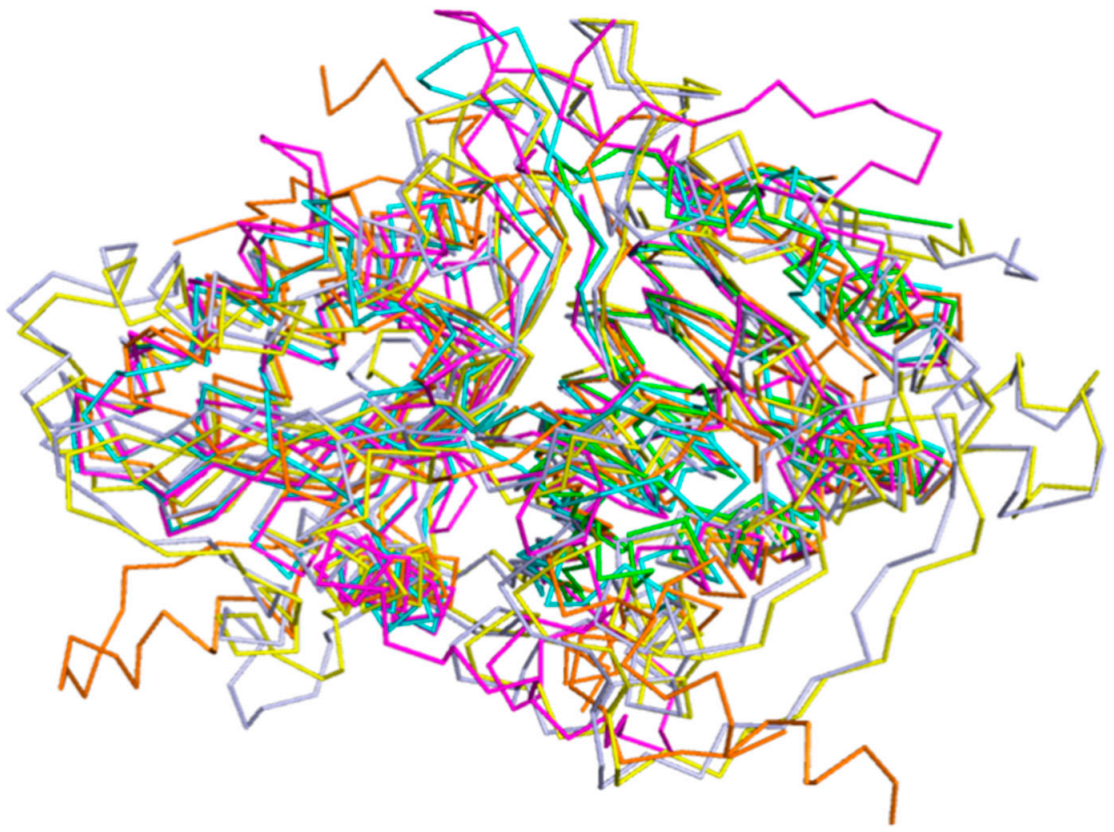
2.5.1. Overall Structure Comparion
2.5.2. Conserved HKD Motifs in the Catalytic Pocket of SpPLD
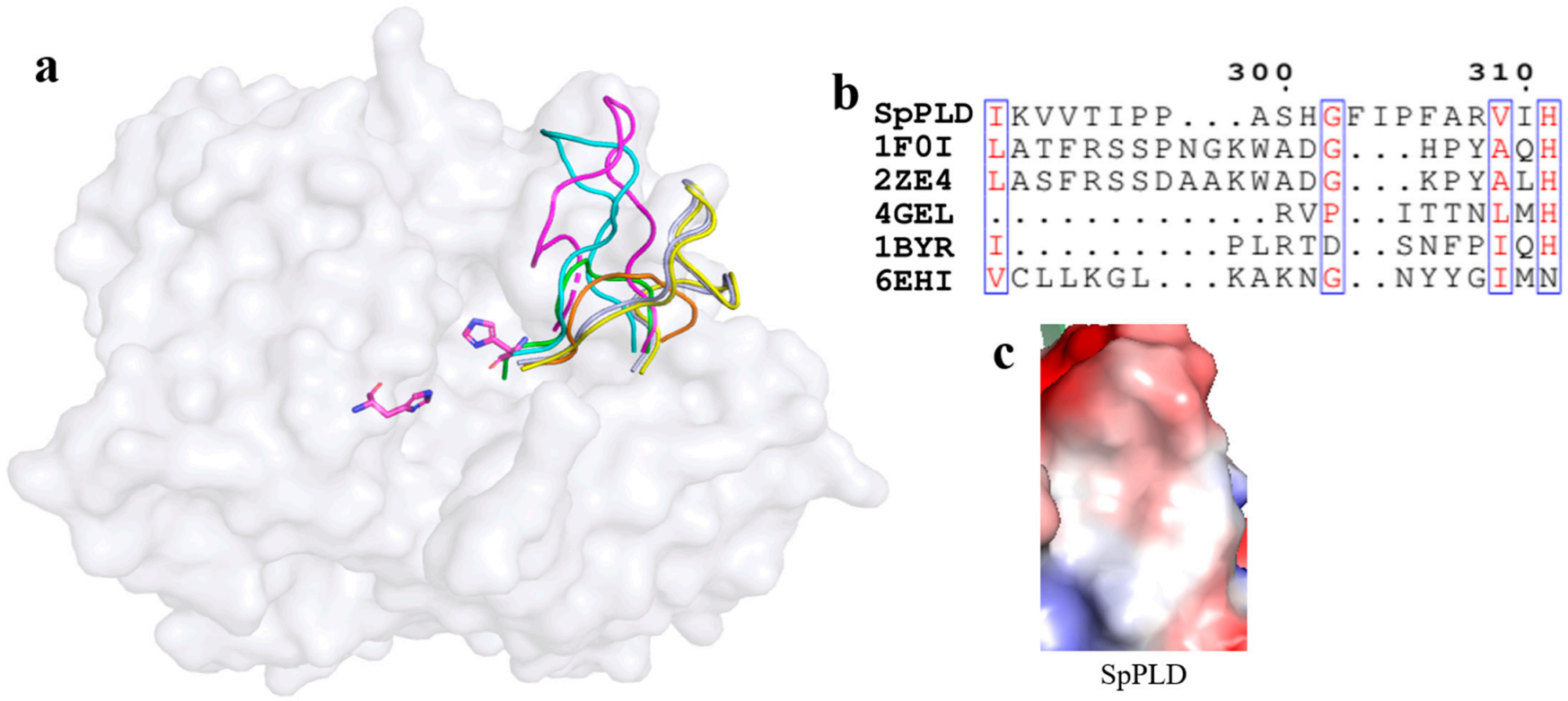
2.5.3. Unique Arrangement in the Catalytic Pocket of SpPLD
2.5.4. An Extent Loop Was Found between α9 and β9 That Distinct from Other PLDs
3. Discussion
3.1. The Unique GS/GT Assembly Unit Found in SpPLD Made it Distinct from Other Members in the Same Family
3.2. Electronic Positive Extented Loop between α9 and β9 Made This Part Unsuitable for Binding the Acyl Chains Anymore
3.3. The Shortened Loop Made SpPLD Unable to Form the Gate to Stable the Substrate
3.4. Loop Entrance Residues Composition Involved in Substrate Recognition Was Also Different from Other PLDs
3.5. A Unique C-Terminal Confromation Was Found in SpPLD
4. Materials and Methods
4.1. Strains, Plasmids and Reagents
4.2. Bioinformatic Analysis of SpPLD
4.3. Protein Expression and Purification
4.4. Crystallization and Structure Determination
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Damnjanovi, J.; Iwasaki, Y. Phospholipase D as a catalyst: Application in phospholipid synthesis, molecular structure and protein engineering. J. Biosci. Bioeng. 2013, 116, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Lv, X.; Li, J.; Shin, H.D.; Du, G.; Liu, L. Combinatorial fine-tuning of phospholipase D expression by Bacillus subtilis WB600 for the production of phosphatidylserine. J. Microbiol. Biotechnol. 2018, 28, 2046–2056. [Google Scholar] [CrossRef]
- Hanahan, D.J.; Chaikoff, I.L. A new phospholipide-splitting enzyme specific for the ester linkage between the nitrogenous base and the phosphoric acid grouping. J. Biol. Chem. 1947, 169, 699–705. [Google Scholar] [CrossRef]
- Hanahan, D.J.; Chaikoff, I.L. On the nature of the P-containing lipides of cabbage leaves and their relation to a phospholipide-splitting enzyme contained in these leaves. J. Biol. Chem. 1948, 172, 191–198. [Google Scholar] [CrossRef]
- Hou, H.J.; Gong, J.S.; Dong, Y.X.; Qin, J.; Li, H.; Li, H.; Lu, Z.M.; Zhang, X.M.; Xu, Z.H.; Shi, J.S. Phospholipase D engineering for improving the biocatalytic synthesis of phosphatidylserine. Bioprocess Biosyst. Eng. 2019, 42, 1185–1194. [Google Scholar] [CrossRef] [PubMed]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Stuckey, J.A.; Dixon, J.E. Crystal structure of a phospholipase D family member. Nat. Struct. Biol. 1999, 6, 278–284. [Google Scholar]
- Nishimasu, H.; Ishizu, H.; Saito, K.; Fukuhara, S.; Kamatani, M.K.; Bonnefond, L.; Matsumoto, N.; Nishizawa, T.; Nakanaga, K.; Aoki, J.; et al. Structure and function of Zucchini endoribonuclease in piRNA biogenesis. Nature 2012, 491, 284–287. [Google Scholar] [CrossRef]
- Ipsaro, J.J.; Haase, A.D.; Knott, S.R.; Joshua-Tor, L.; Hannon, G.J. The structural biochemistry of Zucchini implicates it as a nuclease in piRNA biogenesis. Nature 2012, 491, 279–283. [Google Scholar] [CrossRef] [Green Version]
- Grazulis, S.; Manakova, E.; Roessle, M.; Bochtler, M.; Tamulaitiene, G.; Huber, R.; Siksnys, V. Structure of the metal-independent restriction enzyme BfiI reveals fusion of a specific DNA-binding domain with a nonspecific nuclease. Proc. Natl. Acad. Sci. USA 2005, 102, 15797–15802. [Google Scholar] [CrossRef] [Green Version]
- Celma, L.; Corbinais, C.; Vercruyssen, J.; Veaute, X.; de la Sierra-Gallay, I.L.; Guérois, R.; Busso, D.; Mathieu, A.; Marsin, S.; Quevillon-Cheruel, S.; et al. Structural basis for the substrate selectivity of Helicobacter pylori NucT nuclease activity. PLoS ONE 2017, 12, e0189049. [Google Scholar] [CrossRef] [Green Version]
- Leiros, I.; Secundo, F.; Zambonelli, C.; Servi, S.; Hough, E. The first crystal structure of a phospholipase D. Structure 2000, 8, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Leiros, I.; McSweeney, S.; Hough, E. The reaction mechanism of phospholipase D from Streptomyces sp. Strain PMF. snapshots along the reaction pathway reveal a pentacoordinate reaction intermediate and an unexpected final product. J. Mol. Biol. 2004, 339, 805–820. [Google Scholar] [CrossRef]
- Damnjanović, J.; Nakano, H.; Iwasaki, Y. Deletion of a dynamic surface loop improves stability and changes kinetic behavior of phosphatidylinositol-synthesizing Streptomyces phospholipase D. Biotechnol. Bioeng. 2014, 111, 674–682. [Google Scholar] [CrossRef]
- Inoue, A.; Adachi, M.; Damnjanović, J.; Nakano, H.; Iwasaki, Y. Direct enzymatic synthesis of 1-Phosphatidyl-β-D-glucose by engineered phospholipase D. ChemistrySelect 2016, 1, 4121–4125. [Google Scholar] [CrossRef]
- Damnjanović, J.; Kuroiwa, C.; Tanaka, H.; Ishida, K.; Nakano, H.; Iwasaki, Y. Directing positional specificity in enzymatic synthesis of bioactive 1-phosphatidylinositol by protein engineering of a phospholipase D. Biotechnol. Bioeng. 2016, 113, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Metrick, C.M.; Peterson, E.A.; Santoro, J.C.; Enyedy, I.J.; Murugan, P.; Chen, T.; Michelsen, K.; Cullivan, M.; Spilker, K.A.; Kumar, P.R.; et al. Human PLD structures enable drug design and characterization of isoenzyme selectivity. Nat. Chem. Biol. 2020, 16, 391–399. [Google Scholar] [CrossRef]
- Bowling, F.Z.; Salazar, C.M.; Bell, J.A.; Huq, T.S.; Frohman, M.A.; Airola, M.V. Crystal structure of human PLD1 provides insight into activation by PI (4,5) P2 and RhoA. Nat. Chem. Biol. 2020, 16, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Golovenko, D.; Manakova, E.; Zakrys, L.; Zaremba, M.; Sasnauskas, G.; Gražulis, S.; Siksnys, V. Structural insight into the specificity of the B3 DNA-binding domains provided by the co-crystal structure of the C-terminal fragment of BfiI restriction enzyme. Nucleic Acids Res. 2014, 42, 4113–4122. [Google Scholar] [CrossRef]
- Suzuki, A.; Kakuno, K.; Satio, R.; Iwasaki, Y.; Yamane, T. Crystal structure of phospholipase D from Streptomyces antibioticus. Acta Crystallogr. Sect. A Found. Crystallogr. 2000, 56, S242. [Google Scholar] [CrossRef]
- Voigt, F.; Reuter, M.; Kasaruho, A.; Schulz, E.C.; Pillai, R.S.; Barabas, O. Crystal structure of the primary piRNA biogenesis factor Zucchini reveals similarity to the bacterial PLD endonuclease Nuc. RNA 2012, 18, 2128–2134. [Google Scholar] [CrossRef] [Green Version]
- Neupane, S.; Högberg, N.; Alström, S.; Lucas, S.; Han, J.; Lapidus, A.; Cheng, J.F.; Bruce, D.; Goodwin, L.; Pitluck, S.; et al. Complete genome sequence of the rapeseed plant-growth promoting Serratia plymuthica strain AS9. Stand. Genom. Sci. 2012, 6, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, H.; Berg, G. Impact of formulation procedures on the effect of the biocontrol agent Serratia plymuthica HRO-C48 on Verticillium wilt in oilseed rape. BioControl 2008, 53, 905–916. [Google Scholar] [CrossRef]
- Kalbe, C.; Marten, P.; Berg, G. Strains of the genus Serratia as beneficial rhizobacteria of oilseed rape with antifungal properties. Microbiol. Res. 1996, 151, 433–439. [Google Scholar] [CrossRef]
- Quarles, R.H.; Dawson, R.M. The distribution of phospholipase D in developing and mature plants. Biochem. J. 1969, 112, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, X. Phospholipase D and phosphatidic acid in plant immunity. Plant Sci. 2018, 279, 45–50. [Google Scholar] [CrossRef]
- Sung, T.C.; Roper, R.L.; Zhang, Y.; Rudge, S.A.; Temel, R.; Hammond, S.M.; Morris, A.J.; Moss, B.; Engebrecht, J.; Frohman, M.A. Mutagenesis of phospholipase D defines a superfamily including a trans-Golgi viral protein required for poxvirus pathogenicity. EMBO J. 1997, 16, 4519–4530. [Google Scholar] [CrossRef]
- Gottlin, E.B.; Rudolph, A.E.; Zhao, Y.; Matthews, H.R.; Dixon, J.E. Catalytic mechanism of the phospholipase D superfamily proceeds via a covalent phosphohistidine intermediate. Proc. Natl. Acad. Sci. USA 1998, 95, 9202–9207. [Google Scholar] [CrossRef] [Green Version]
- Ogino, C.; Kuroda, S.I.; Tokuyama, S.; Kondo, A.; Shimizu, N.; Tanizawa, K.; Fukuda, H. Phospholipase D from Streptoverticillium cinnamoneum: Protein engineering and application for phospholipid production. J. Mol. Catal. B Enzym. 2003, 23, 107–115. [Google Scholar] [CrossRef]
- Damnjanović, J.; Takahashi, R.; Suzuki, A.; Nakano, H.; Iwasaki, Y. Improving thermostability of phosphatidylinositol-synthesizing Streptomyces phospholipase D. Protein Eng. Des. Sel. 2012, 25, 415–424. [Google Scholar] [CrossRef]
- Uesugi, Y.; Arima, J.; Iwabuchi, M.; Hatanaka, T. Sensor of phospholipids in Streptomyces phospholipase D. FEBS J. 2007, 274, 2672–2681. [Google Scholar] [CrossRef]
- Uesugi, Y.; Hatanaka, T. Phospholipase D mechanism using Streptomyces PLD. Biochim. Biophys. Acta. 2009, 1791, 962–969. [Google Scholar] [CrossRef] [PubMed]
- Armenteros, J.J.A.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; Heijne, G.V.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-Coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holm, L.; Rosenström, P. Dali server: Conservation mapping in 3D. Nucleic Acids Res. 2010, 38, W545–W549. [Google Scholar] [CrossRef]
- Hendrickson, W.A.; Horton, J.R.; LeMaster, D.M. Selenomethionyl proteins produced for analysis by multiwavelength anomalous diffraction (MAD): A vehicle for direct determination of three-dimensional structure. EMBO J. 1990, 9, 1665–1672. [Google Scholar] [CrossRef]
- Kabsch, W. XDS. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Hough, M.A.; Wilson, K.S. From crystal to structure with CCP4. Acta Crystallogr. Sect. D Struct. Biol. 2018, 74, 67. [Google Scholar] [CrossRef] [Green Version]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkóczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.W.; Jain, S.; McCoy, A.J.; et al. Macromolecular structure determination using X-rays, neutrons and electrons: Recent developments in Phenix. Acta Crystallogr. Sect. D Struct. Biol. 2019, 75, 861–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskowski, R.A.; Moss, D.S.; Thornton, J.M. Main-chain Bond Lengths and Bond Angles in Protein Structures. J. Mol. Biol. 1993, 231, 1049–1067. [Google Scholar] [CrossRef] [PubMed]
- Bramucci, E.; Paiardini, A.; Bossa, F.; Pascarella, S. PyMod: Sequence similarity searches, multiple sequence-structure alignments, and homology modeling within PyMOL. BMC Bioinf. 2012, 13, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Taxonomy | Name | Organism | PDB Codes | Deposition Date | Reference |
|---|---|---|---|---|---|---|
| 1 | Microorganism | Nuc | Salmonella typhimurium | 1BYS (complexed with Tungstate) 1BYR | 1998 | [7] |
| 2 | Microorganism | PLDPMF | Streptomyces sp. PMF | 1F0I (apo) 1V0R (Tungstate-inhibited form) 1V0S (Uninhibited form) 1V0T (complex with product glycerophosphate) 1V0U (complex with product glycerophosphate) 1V0V (complex with substrate dibutyrylphosphatidylcholine) 1V0W (complex with substrate dibutyrylphosphatidylcholine) 1V0Y (complex with substrate dibutyrylphosphatidylcholine) | 2000 2004 | [12,13] |
| 3 | Microorganism | BfiI | Bacillus firmus | 2C1L 3ZI5 (C-Terminal Fragment of BfiI domain in complex with cognate DNA) | 2005 2013 | [10,19] |
| 4 | Microorganism | SaPLD | Streptomyces antibioticus | 2ZE4 2ZE9 (H168A mutant complex with phosphatidylcholine) | 2007 | [20] |
| 5 | Mammal | mZuc / PLD6 / MitoPLD | Mus musculus | 4GGJ 4GGK (complex with tungstate) | 2012 | [9] |
| 6 | Arthropoda | Zuc | Drosophila melanogaster | 4GEL 4GEN (monomer) 4GEM (K171A mutant) 4H4A (C-terminal domain) | 2012 | [21] |
| 7 | Microorganism | NucT | Helicobacter pylori | 6EHI | 2017 | [11] |
| 8 | Mammal | hPLD1 | Homo sapiens | 6U8Z 6OHR (complex with compound 5) | 2019 | [18] |
| 9 | Mammal | hPLD2 | Homo sapiens | 6OHM (complex with tungstate) 6OHO6OHP (complex with halopemide) 6OHQ (complex with compound 4) 6OHS (complex with compound ML299) | 2019 | [17] |
| 10 | Microorganism | SpPLD | Serratia plymuthica strain AS9 | 7E0M | 2021 | Present research |
| SpPLD | |
|---|---|
| Wavelength | |
| Resolution range | 23.88–1.79 (1.854–1.79) |
| Space group | P212121 |
| Unit cell | a = 78.78Å, b = 97.57 Å, c = 117.03 Å, α = 90 Å, β = 90 Å, γ = 90 Å |
| Total reflections | 928,590 (56,570) |
| Unique reflections | 79,256 (6680) |
| Multiplicity | 11.7 (8.4) |
| Completeness (%) | 96.98 (79.33) |
| Mean I/sigma(I) | 12.77 (1.47) |
| Wilson B-factor | 25.38 |
| R-merge | 0.1337 (1.706) |
| R-meas | 0.1397 (1.822) |
| R-pim | 0.0396 (0.6073) |
| CC1/2 | 0.997 (0.503) |
| CC* | 0.999 (0.818) |
| Reflections used in refinement | 82,957 (6676) |
| Reflections used for R-free | 4150 (335) |
| R-work | 0.1695 (0.2706) |
| R-free | 0.1848 (0.3129) |
| CC(work) | 0.969 (0.788) |
| CC(free) | 0.967 (0.713) |
| Number of non-hydrogen atoms | 6902 |
| macromolecules | 6104 |
| ligands | 5 |
| solvent | 793 |
| Protein residues | 772 |
| RMS(bonds) | 0.011 |
| RMS(angles) | 1.47 |
| Ramachandran favored (%) | 97.14 |
| Ramachandran allowed (%) | 2.86 |
| Ramachandran outliers (%) | 0.00 |
| Rotamer outliers (%) | 0.93 |
| Clashscore | 4.92 |
| Average B-factor | 30.06 |
| macromolecules | 28.77 |
| ligands | 38.60 |
| solvent | 39.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Liu, S.; Mao, X.; Cui, R.; Yang, B.; Wang, Y. Crystal Structure of a Phospholipase D from the Plant-Associated Bacteria Serratia plymuthica Strain AS9 Reveals a Unique Arrangement of Catalytic Pocket. Int. J. Mol. Sci. 2021, 22, 3219. https://doi.org/10.3390/ijms22063219
Wang F, Liu S, Mao X, Cui R, Yang B, Wang Y. Crystal Structure of a Phospholipase D from the Plant-Associated Bacteria Serratia plymuthica Strain AS9 Reveals a Unique Arrangement of Catalytic Pocket. International Journal of Molecular Sciences. 2021; 22(6):3219. https://doi.org/10.3390/ijms22063219
Chicago/Turabian StyleWang, Fanghua, Siyu Liu, Xuejing Mao, Ruiguo Cui, Bo Yang, and Yonghua Wang. 2021. "Crystal Structure of a Phospholipase D from the Plant-Associated Bacteria Serratia plymuthica Strain AS9 Reveals a Unique Arrangement of Catalytic Pocket" International Journal of Molecular Sciences 22, no. 6: 3219. https://doi.org/10.3390/ijms22063219
APA StyleWang, F., Liu, S., Mao, X., Cui, R., Yang, B., & Wang, Y. (2021). Crystal Structure of a Phospholipase D from the Plant-Associated Bacteria Serratia plymuthica Strain AS9 Reveals a Unique Arrangement of Catalytic Pocket. International Journal of Molecular Sciences, 22(6), 3219. https://doi.org/10.3390/ijms22063219