
Expression Pattern of α-Tubulin, Inversin and Its Target Dishevelled-1 and Morphology of Primary Cilia in Normal Human Kidney Development and Diseases

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Light Microscopy, Electron Microscopy and Immunohistochemistry (α-Tubulin) of Healthy and Pathologically Changed Postnatal Human Kidney Tissue
2.1.1. Healthy Postnatal Kidney
2.1.2. CNF
2.1.3. MCDK
2.1.4. FSGS
2.2. Immunohistochemical Staining to α-Tubulin, Inversin and DVL-1- and Statistical Analysis of Developing and Healthy Postnatal Human Kidneys
2.2.1. α-Tubulin
2.2.2. Double Immunofluorescence Staining to Inversin and DVL-1 and DAPI Nuclear Stain in Developing and Healthy Postnatal Kidneys
Inversin Expression in Developing and Postnatal Kidney Tissue
DVL-1 Expression in Developing and Postnatal Kidney Tissue
2.2.3. Differences in Expression of α-Tubulin, Inversin and DVL-1 between Different Developmental Kidney Stages and Postnatal Kidneys
2.2.4. Relationship between Expressions of α-Tubulin and Inversin to DVL-1 in Developing and Postnatal Kidney Tissue
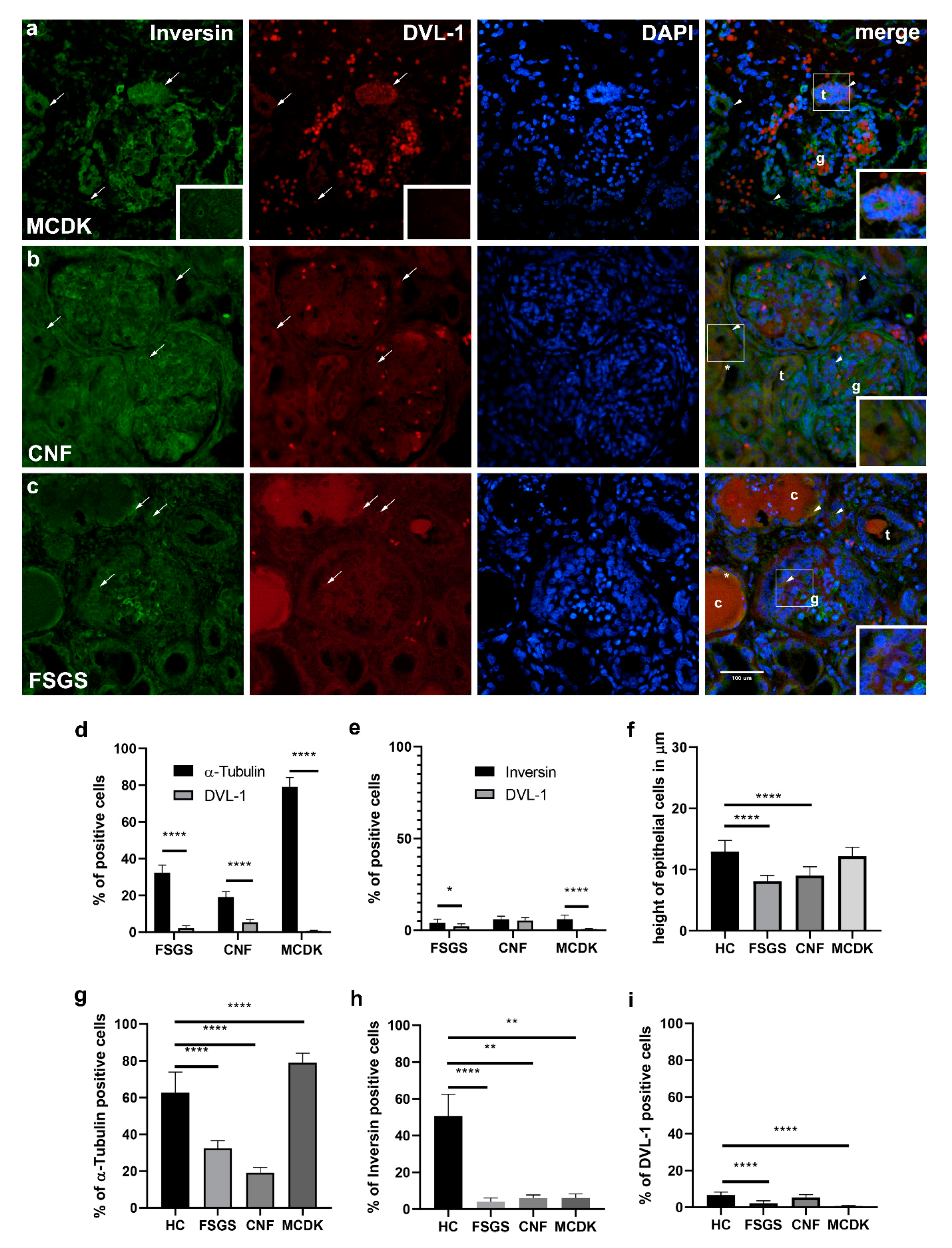
2.2.5. Comparison of Immunohistochemical Staining to α-Tubulin, Inversin and DVL-1- and Statistical Analysis of Pathologically Changed Kidney Tissue (MCDK, CNF, FSGS)
α-Tubulin
Inversin
DVL-1
2.2.6. Relationship between Expressions of α-Tubulin and Inversin to DVL-1 in Pathologically Changed Kidney Tissue (MCDK, CNF, FSGS)
2.2.7. Differences in Epithelial Cell Height of Proximal Convoluted Tubules between Healthy Control and Pathologically Changed Kidney Tissue (MCDK, CNF, FSGS)
2.2.8. Differences in Primary Cilia Length between Healthy Control and Pathologically Changed Kidney Tissue (MCDK, CNF, FSGS)
3. Discussion
4. Materials and Methods
4.1. Human Samples
4.2. Immunohistochemistry
4.3. Electron Microscopy
4.4. Genetic Analysis
4.5. Data Analysis
4.6. Semiquantitative Analysis
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| DVL-1 | Dishevelled-1 |
| GW | Gestational week |
| MET | Mesenchymal to epithelial transition |
| CKD | Chronic kidney disease |
| MCDK | Multicystic dysplastic kidneys |
| FSGS | Focal segmental glomerulosclerosis |
| CNF | Nephrotic syndrome of the Finnish type |
| pct | Proximal convoluted tubules |
| dct | Distal convoluted tubules |
| g | Glomeruli |
References
- US Food and Drug Administration. Guidance for Industry: Nonclinical Safety Evaluation of Pediatric Drug Products; Center for Drug Evaluation and Research: Rockville, MD, USA, 2006. [Google Scholar]
- Saraga-Babic, M.; Vukojevic, K.; Bocina, I.; Drnasin, K.; Saraga, M. Ciliogenesis in normal human kidney development and post-natal life. Pediatr. Nephrol. 2012, 27, 55–63. [Google Scholar] [CrossRef]
- Pei, D.; Shu, X.; Gassama-Diagne, A.; Thiery, J.P. Mesenchymal-epithelial transition in development and reprogramming. Nat. Cell Biol. 2019, 21, 44–53. [Google Scholar] [CrossRef]
- Blake, J.; Rosenblum, N.D. Renal branching morphogenesis: Morphogenetic and signaling mechanisms. Semin. Cell Dev. Biol. 2014, 36, 2–12. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, L.L.; McMahon, A.P. Induction and patterning of the metanephric nephron. Semin. Cell Dev. Biol. 2014, 36, 31–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, M.; Shibata, S.; Shu, Y. Pathogenesis of dysplastic kidney associated with urinary tract obstruction in utero. Nephrol. Dial. Transplant. 2002, 17 (Suppl. S9), 37–38. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhou, C.J.; Liu, Y. Wnt Signaling in Kidney Development and Disease. Prog. Mol. Biol. Transl. Sci. 2018, 153, 181–207. [Google Scholar] [CrossRef] [PubMed]
- Marshall, W.F.; Nonaka, S. Cilia: Tuning in to the cell’s antenna. Curr. Biol. CB 2006, 16, R604–R614. [Google Scholar] [CrossRef] [Green Version]
- Poureetezadi, S.J.; Cheng, C.N.; Chambers, J.M.; Drummond, B.E.; Wingert, R.A. Prostaglandin signaling regulates nephron segment patterning of renal progenitors during zebrafish kidney development. eLife 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Mirvis, M.; Stearns, T.; James Nelson, W. Cilium structure, assembly, and disassembly regulated by the cytoskeleton. Biochem. J. 2018, 475, 2329–2353. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.; Liu, Y.; Zhu, B.; Ding, K.; Yao, T.P.; Chen, F.; Zhan, L.; Xu, P.; Ehrlich, M.; Liang, T.; et al. Loss of alpha-Tubulin Acetylation Is Associated with TGF-beta-induced Epithelial-Mesenchymal Transition. J. Biol. Chem. 2016, 291, 5396–5405. [Google Scholar] [CrossRef] [Green Version]
- Tran, P.V.; Sharma, M.; Li, X.; Calvet, J.P. Developmental signaling: Does it bridge the gap between cilia dysfunction and renal cystogenesis? Birth Defects Res. Part C Embryo Today Rev. 2014, 102, 159–173. [Google Scholar] [CrossRef] [Green Version]
- Warady, B.A.; Chadha, V. Chronic kidney disease in children: The global perspective. Pediatr. Nephrol. 2007, 22, 1999–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Shi, Z.; Jiang, D. "Watch and Wait" Strategy for Multicystic Dysplastic Kidney (MCDK): Status Survey of Perceptions, Attitudes, and Treatment Selection in Chinese Pediatric Urologists and Pediatric Surgeons. Front. Pediatr. 2020, 8, 423. [Google Scholar] [CrossRef]
- Cardona-Grau, D.; Kogan, B.A. Update on Multicystic Dysplastic Kidney. Curr. Urol. Rep. 2015, 16, 67. [Google Scholar] [CrossRef] [PubMed]
- Kiffel, J.; Rahimzada, Y.; Trachtman, H. Focal segmental glomerulosclerosis and chronic kidney disease in pediatric patients. Adv. Chronic Kidney Dis. 2011, 18, 332–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Agati, V.D.; Kaskel, F.J.; Falk, R.J. Focal segmental glomerulosclerosis. N. Engl. J. Med. 2011, 365, 2398–2411. [Google Scholar] [CrossRef] [Green Version]
- Holmberg, C.; Antikainen, M.; Ronnholm, K.; Ala Houhala, M.; Jalanko, H. Management of congenital nephrotic syndrome of the Finnish type. Pediatr. Nephrol. 1995, 9, 87–93. [Google Scholar] [CrossRef]
- Saraga, M.; Vukojevic, K.; Krzelj, V.; Puretic, Z.; Bocina, I.; Durdov, M.G.; Weber, S.; Dworniczak, B.; Ljubanovic, D.G.; Saraga-Babic, M. Mechanism of cystogenesis in nephrotic kidneys: A histopathological study. BMC Nephrol. 2014, 15, 3. [Google Scholar] [CrossRef] [Green Version]
- Reiser, J.; Sever, S. Podocyte biology and pathogenesis of kidney disease. Annu. Rev. Med. 2013, 64, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Thomas, M.C. Pathogenesis and progression of proteinuria. Contrib. Nephrol. 2011, 170, 48–56. [Google Scholar] [CrossRef]
- Vukojevic, K.; Raguz, F.; Saraga, M.; Filipovic, N.; Bocina, I.; Kero, D.; Glavina Durdov, M.; Martinovic, V.; Saraga-Babic, M. Glomeruli from patients with nephrin mutations show increased number of ciliated and poorly differentiated podocytes. Acta Histochem. 2018, 120, 748–756. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, Y. Wnt/beta-catenin signalling and podocyte dysfunction in proteinuric kidney disease. Nat. Rev. Nephrol. 2015, 11, 535–545. [Google Scholar] [CrossRef]
- Iglesias, D.M.; Hueber, P.A.; Chu, L.; Campbell, R.; Patenaude, A.M.; Dziarmaga, A.J.; Quinlan, J.; Mohamed, O.; Dufort, D.; Goodyer, P.R. Canonical WNT signaling during kidney development. Am. J. Physiol. Ren. Physiol. 2007, 293, F494–F500. [Google Scholar] [CrossRef] [Green Version]
- Park, J.S.; Valerius, M.T.; McMahon, A.P. Wnt/beta-catenin signaling regulates nephron induction during mouse kidney development. Development 2007, 134, 2533–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simons, M.; Gloy, J.; Ganner, A.; Bullerkotte, A.; Bashkurov, M.; Kronig, C.; Schermer, B.; Benzing, T.; Cabello, O.A.; Jenny, A.; et al. Inversin, the gene product mutated in nephronophthisis type II, functions as a molecular switch between Wnt signaling pathways. Nat. Genet. 2005, 37, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Morgan, D.; Eley, L.; Sayer, J.; Strachan, T.; Yates, L.M.; Craighead, A.S.; Goodship, J.A. Expression analyses and interaction with the anaphase promoting complex protein Apc2 suggest a role for inversin in primary cilia and involvement in the cell cycle. Hum. Mol. Genet. 2002, 11, 3345–3350. [Google Scholar] [CrossRef] [Green Version]
- Nurnberger, J.; Kribben, A.; Opazo Saez, A.; Heusch, G.; Philipp, T.; Phillips, C.L. The Invs gene encodes a microtubule-associated protein. J. Am. Soc. Nephrol. 2004, 15, 1700–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mochizuki, T.; Saijoh, Y.; Tsuchiya, K.; Shirayoshi, Y.; Takai, S.; Taya, C.; Yonekawa, H.; Yamada, K.; Nihei, H.; Nakatsuji, N.; et al. Cloning of inv, a gene that controls left/right asymmetry and kidney development. Nature 1998, 395, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Otto, E.A.; Schermer, B.; Obara, T.; O’Toole, J.F.; Hiller, K.S.; Mueller, A.M.; Ruf, R.G.; Hoefele, J.; Beekmann, F.; Landau, D.; et al. Mutations in INVS encoding inversin cause nephronophthisis type 2, linking renal cystic disease to the function of primary cilia and left-right axis determination. Nat. Genet. 2003, 34, 413–420. [Google Scholar] [CrossRef]
- Lienkamp, S.; Ganner, A.; Boehlke, C.; Schmidt, T.; Arnold, S.J.; Schafer, T.; Romaker, D.; Schuler, J.; Hoff, S.; Powelske, C.; et al. Inversin relays Frizzled-8 signals to promote proximal pronephros development. Proc. Natl. Acad. Sci. USA 2010, 107, 20388–20393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.; Mazzeu, J.F.; Hoischen, A.; Jhangiani, S.N.; Gambin, T.; Alcino, M.C.; Penney, S.; Saraiva, J.M.; Hove, H.; Skovby, F.; et al. DVL1 frameshift mutations clustering in the penultimate exon cause autosomal-dominant Robinow syndrome. Am. J. Hum. Genet. 2015, 96, 612–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurnberger, J.; Bacallao, R.L.; Phillips, C.L. Inversin forms a complex with catenins and N-cadherin in polarized epithelial cells. Mol. Biol. Cell 2002, 13, 3096–3106. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, D.; Saijoh, Y.; Nonaka, S.; Sasaki, G.; Ikawa, Y.; Yokoyama, T.; Hamada, H. The left-right determinant Inversin is a component of node monocilia and other 9+0 cilia. Development 2003, 130, 1725–1734. [Google Scholar] [CrossRef] [Green Version]
- Phillips, C.L.; Miller, K.J.; Filson, A.J.; Nurnberger, J.; Clendenon, J.L.; Cook, G.W.; Dunn, K.W.; Overbeek, P.A.; Gattone, V.H., 2nd; Bacallao, R.L. Renal cysts of inv/inv mice resemble early infantile nephronophthisis. J. Am. Soc. Nephrol. 2004, 15, 1744–1755. [Google Scholar] [CrossRef] [Green Version]
- Oud, M.M.; van Bon, B.W.; Bongers, E.M.; Hoischen, A.; Marcelis, C.L.; de Leeuw, N.; Mol, S.J.; Mortier, G.; Knoers, N.V.; Brunner, H.G.; et al. Early presentation of cystic kidneys in a family with a homozygous INVS mutation. Am. J. Med. Genet. Part A 2014, 164A, 1627–1634. [Google Scholar] [CrossRef]
- Wang, J.; Sinha, T.; Wynshaw-Boris, A. Wnt signaling in mammalian development: Lessons from mouse genetics. Cold Spring Harb. Perspect. Biol. 2012, 4, a007963. [Google Scholar] [CrossRef]
- Ganner, A.; Lienkamp, S.; Schafer, T.; Romaker, D.; Wegierski, T.; Park, T.J.; Spreitzer, S.; Simons, M.; Gloy, J.; Kim, E.; et al. Regulation of ciliary polarity by the APC/C. Proc. Natl. Acad. Sci. USA 2009, 106, 17799–17804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schueler, M.; Braun, D.A.; Chandrasekar, G.; Gee, H.Y.; Klasson, T.D.; Halbritter, J.; Bieder, A.; Porath, J.D.; Airik, R.; Zhou, W.; et al. DCDC2 mutations cause a renal-hepatic ciliopathy by disrupting Wnt signaling. Am. J. Hum. Genet. 2015, 96, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Wynshaw-Boris, A. Dishevelled: In vivo roles of a multifunctional gene family during development. Curr. Top. Dev. Biol. 2012, 101, 213–235. [Google Scholar] [CrossRef]
- Lienkamp, S.; Ganner, A.; Walz, G. Inversin, Wnt signaling and primary cilia. Differentiation 2012, 83, S49–S55. [Google Scholar] [CrossRef]
- Bellavia, S.; Dahan, K.; Terryn, S.; Cosyns, J.P.; Devuyst, O.; Pirson, Y. A homozygous mutation in INVS causing juvenile nephronophthisis with abnormal reactivity of the Wnt/beta-catenin pathway. Nephrol. Dial. Transplant. 2010, 25, 4097–4102. [Google Scholar] [CrossRef] [Green Version]
- Gerdes, J.M.; Liu, Y.; Zaghloul, N.A.; Leitch, C.C.; Lawson, S.S.; Kato, M.; Beachy, P.A.; Beales, P.L.; DeMartino, G.N.; Fisher, S.; et al. Disruption of the basal body compromises proteasomal function and perturbs intracellular Wnt response. Nat. Genet. 2007, 39, 1350–1360. [Google Scholar] [CrossRef]
- Saito, S.; Tampe, B.; Muller, G.A.; Zeisberg, M. Primary cilia modulate balance of canonical and non-canonical Wnt signaling responses in the injured kidney. Fibrogenesis Tissue Repair 2015, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Hertig, A.; Verine, J.; Mougenot, B.; Jouanneau, C.; Ouali, N.; Sebe, P.; Glotz, D.; Ancel, P.Y.; Rondeau, E.; Xu-Dubois, Y.C. Risk factors for early epithelial to mesenchymal transition in renal grafts. Am. J. Transplant. 2006, 6, 2937–2946. [Google Scholar] [CrossRef]
- Fabbri, L.; Bost, F.; Mazure, N.M. Primary Cilium in Cancer Hallmarks. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivante, A.; Mark-Danieli, M.; Davidovits, M.; Harari-Steinberg, O.; Omer, D.; Gnatek, Y.; Cleper, R.; Landau, D.; Kovalski, Y.; Weissman, I.; et al. Renal hypodysplasia associates with a WNT4 variant that causes aberrant canonical WNT signaling. J. Am. Soc. Nephrol. 2013, 24, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Eley, L.; Turnpenny, L.; Yates, L.M.; Craighead, A.S.; Morgan, D.; Whistler, C.; Goodship, J.A.; Strachan, T. A perspective on inversin. Cell Biol. Int. 2004, 28, 119–124. [Google Scholar] [CrossRef]
- Papakrivopoulou, E.; Dean, C.H.; Copp, A.J.; Long, D.A. Planar cell polarity and the kidney. Nephrol. Dial. Transplant. 2014, 29, 1320–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loughna, P.; Chitty, L.; Evans, T.; Chudleigh, T. Fetal size and dating: Charts recommended for clinical obstetric practice. Ultrasound 2009, 17. [Google Scholar] [CrossRef]
- Williams, J.R. The Declaration of Helsinki and public health. Bull. World Health Organ. 2008, 86, 650–652. [Google Scholar] [CrossRef] [PubMed]
- Vukojevic, K.; Skobic, H.; Saraga-Babic, M. Proliferation and differentiation of glial and neuronal progenitors in the development of human spinal ganglia. Differentiation 2009, 78, 91–98. [Google Scholar] [CrossRef]
- Racetin, A.; Raguz, F.; Durdov, M.G.; Kunac, N.; Saraga, M.; Sanna-Cherchi, S.; Soljic, V.; Martinovic, V.; Petricevic, J.; Kostic, S.; et al. Immunohistochemical expression pattern of RIP5, FGFR1, FGFR2 and HIP2 in the normal human kidney development. Acta Histochem. 2019, 121, 531–538. [Google Scholar] [CrossRef]
- Vukojevic, K.; Kero, D.; Novakovic, J.; Kalibovic Govorko, D.; Saraga-Babic, M. Cell proliferation and apoptosis in the fusion of human primary and secondary palates. Eur. J. Oral Sci. 2012, 120, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Vukusic Pusic, T.; Janjic, T.; Dujmovic, I.; Poljicanin, A.; Soljic, V.; Saraga-Babic, M.; Vukojevic, K. The involvement of proliferation and apoptosis in the early human gonad development. J. Mol. Histol. 2013, 44, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Lenkkeri, U.; Mannikko, M.; McCready, P.; Lamerdin, J.; Gribouval, O.; Niaudet, P.M.; Antignac, C.K.; Kashtan, C.E.; Homberg, C.; Olsen, A.; et al. Structure of the gene for congenital nephrotic syndrome of the finnish type (NPHS1) and characterization of mutations. Am. J. Hum. Genet. 1999, 64, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | α-Tubulin | Inversin | DVL-1 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GW/ystructure * | 14 | 15 | 16 | 21 | 38 | 1.5 | 7 | 14 | 15 | 16 | 21 | 38 | 1.5 | 7 | 14 | 15 | 16 | 21 | 38 | 1.5 | 7 |
| Pct | 3 | 3 | 3 | 3 | 2 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 3 | 2 | 1 |
| Dct | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 2 | 2 | 2 | 2 | 2 | 2 | 3 | 1 | 2 | 2 | 2 | 3 | 1 | 1 |
| G | 2 | 1 | 2 | 3 | 1 | 2 | 2 | 2 | 3 | 3 | 3 | 3 | 3 | 3 | 1 | 1 | 1 | 3 | 3 | 3 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solic, I.; Racetin, A.; Filipovic, N.; Mardesic, S.; Bocina, I.; Galesic-Ljubanovic, D.; Glavina Durdov, M.; Saraga-Babić, M.; Vukojevic, K. Expression Pattern of α-Tubulin, Inversin and Its Target Dishevelled-1 and Morphology of Primary Cilia in Normal Human Kidney Development and Diseases. Int. J. Mol. Sci. 2021, 22, 3500. https://doi.org/10.3390/ijms22073500
Solic I, Racetin A, Filipovic N, Mardesic S, Bocina I, Galesic-Ljubanovic D, Glavina Durdov M, Saraga-Babić M, Vukojevic K. Expression Pattern of α-Tubulin, Inversin and Its Target Dishevelled-1 and Morphology of Primary Cilia in Normal Human Kidney Development and Diseases. International Journal of Molecular Sciences. 2021; 22(7):3500. https://doi.org/10.3390/ijms22073500
Chicago/Turabian StyleSolic, Ivana, Anita Racetin, Natalija Filipovic, Snjezana Mardesic, Ivana Bocina, Danica Galesic-Ljubanovic, Meri Glavina Durdov, Mirna Saraga-Babić, and Katarina Vukojevic. 2021. "Expression Pattern of α-Tubulin, Inversin and Its Target Dishevelled-1 and Morphology of Primary Cilia in Normal Human Kidney Development and Diseases" International Journal of Molecular Sciences 22, no. 7: 3500. https://doi.org/10.3390/ijms22073500
APA StyleSolic, I., Racetin, A., Filipovic, N., Mardesic, S., Bocina, I., Galesic-Ljubanovic, D., Glavina Durdov, M., Saraga-Babić, M., & Vukojevic, K. (2021). Expression Pattern of α-Tubulin, Inversin and Its Target Dishevelled-1 and Morphology of Primary Cilia in Normal Human Kidney Development and Diseases. International Journal of Molecular Sciences, 22(7), 3500. https://doi.org/10.3390/ijms22073500