
Design and In Vitro Evaluation of Splice-Switching Oligonucleotides Bearing Locked Nucleic Acids, Amido-Bridged Nucleic Acids, and Guanidine-Bridged Nucleic Acids


Abstract
:1. Introduction
2. Results
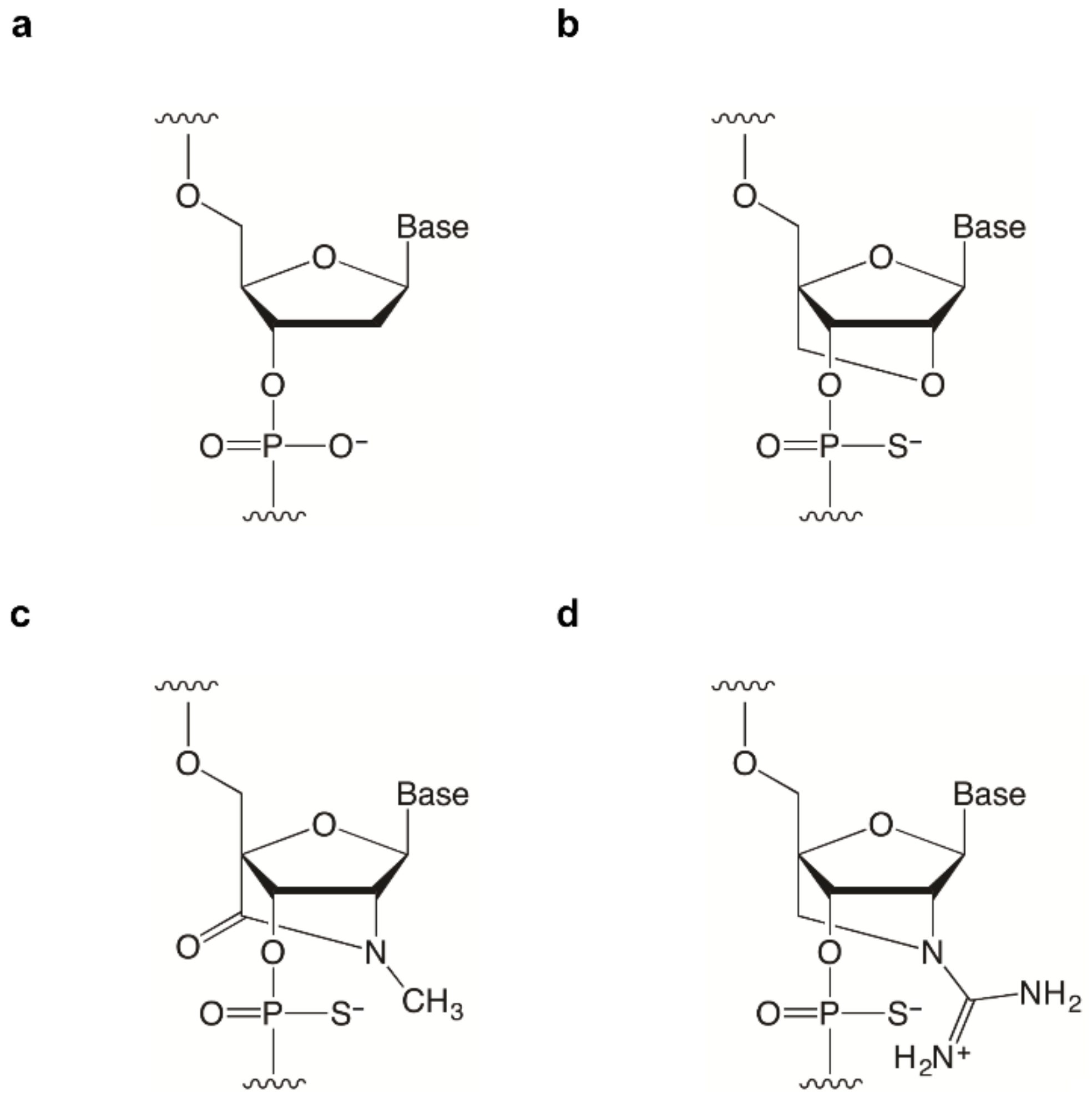
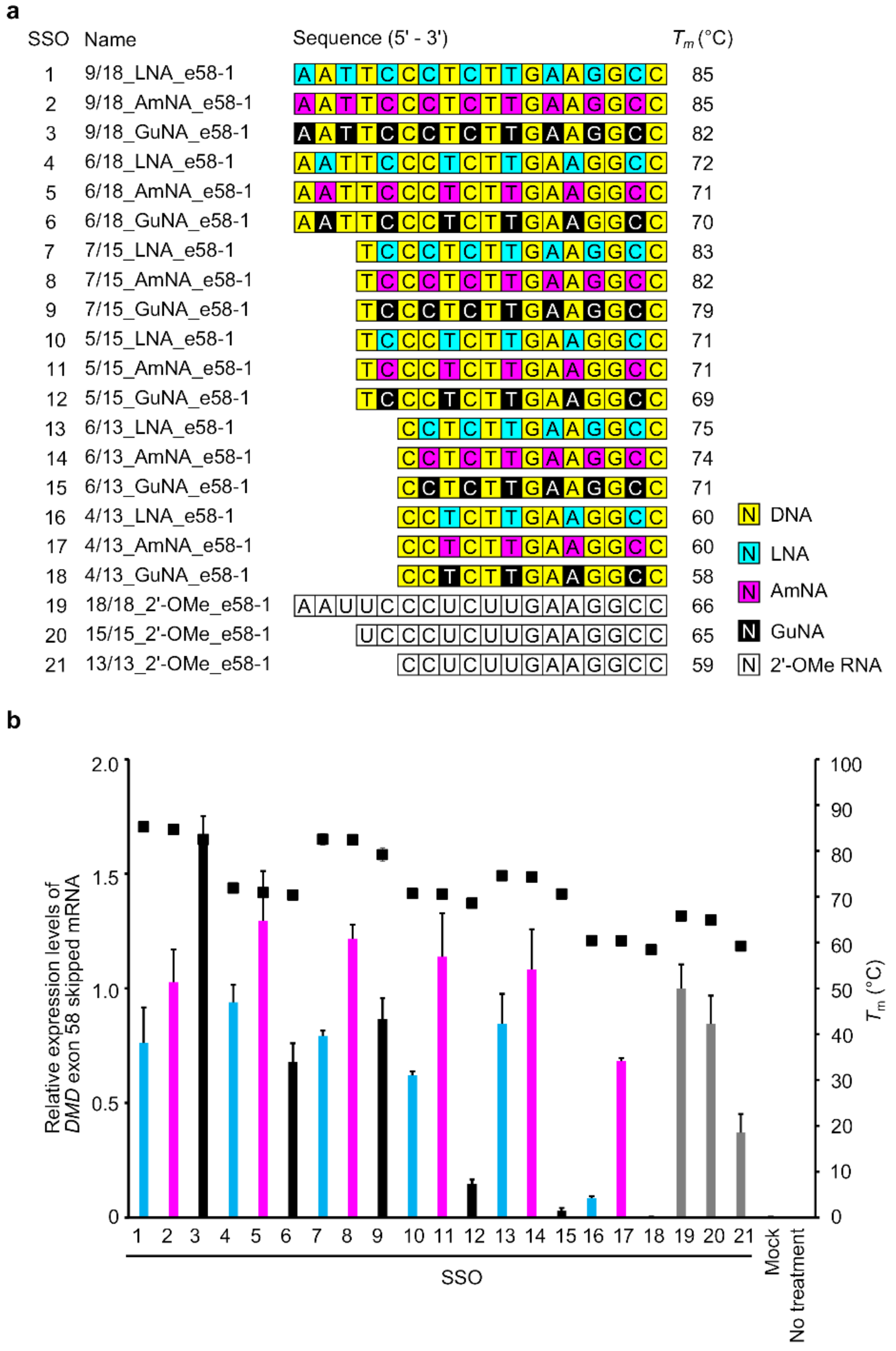
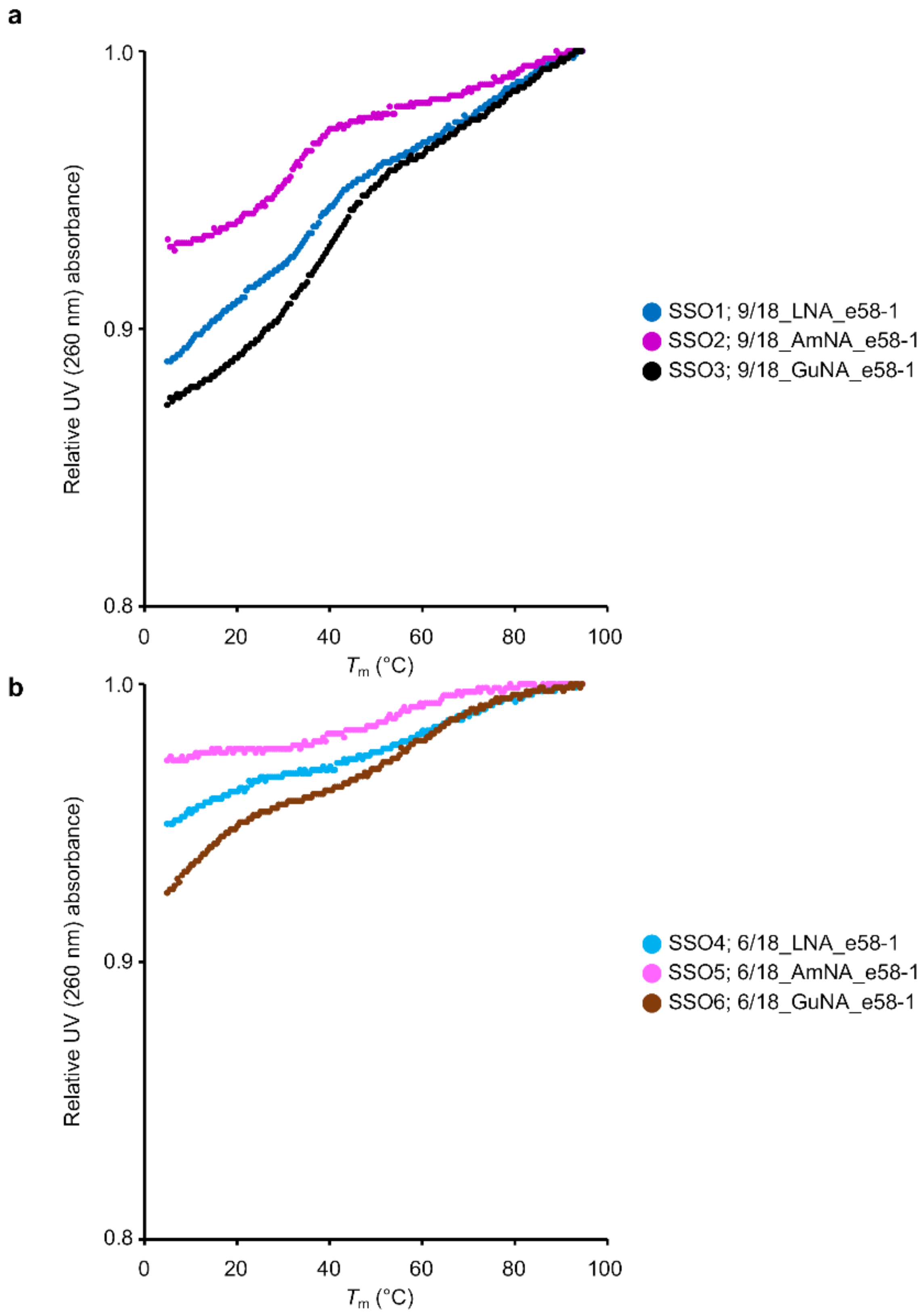
2.1. Design of BNA-Modified SSOs Targeting DMD Exon 58 and Evaluation Using a Stably Transfected Cell Line
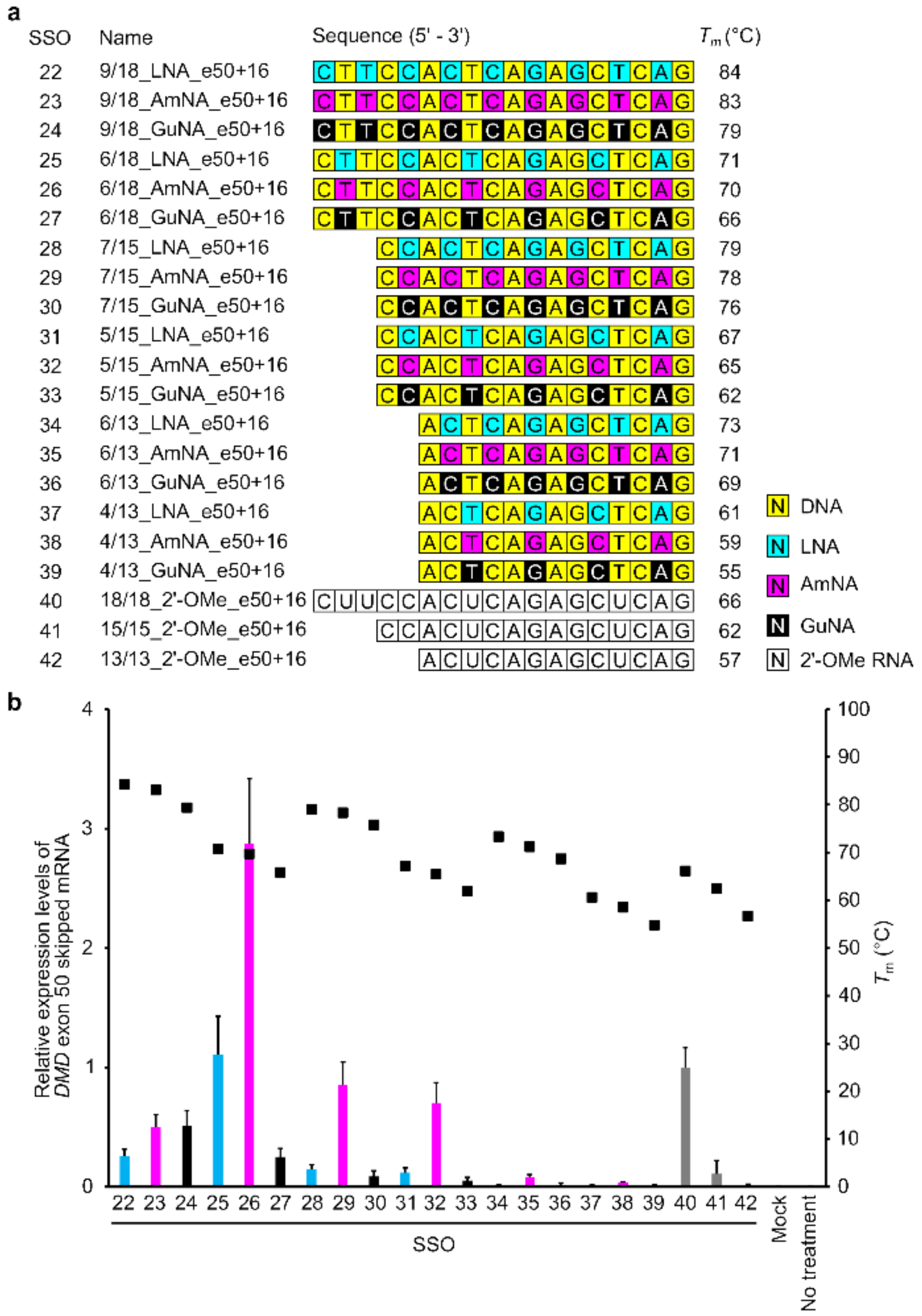
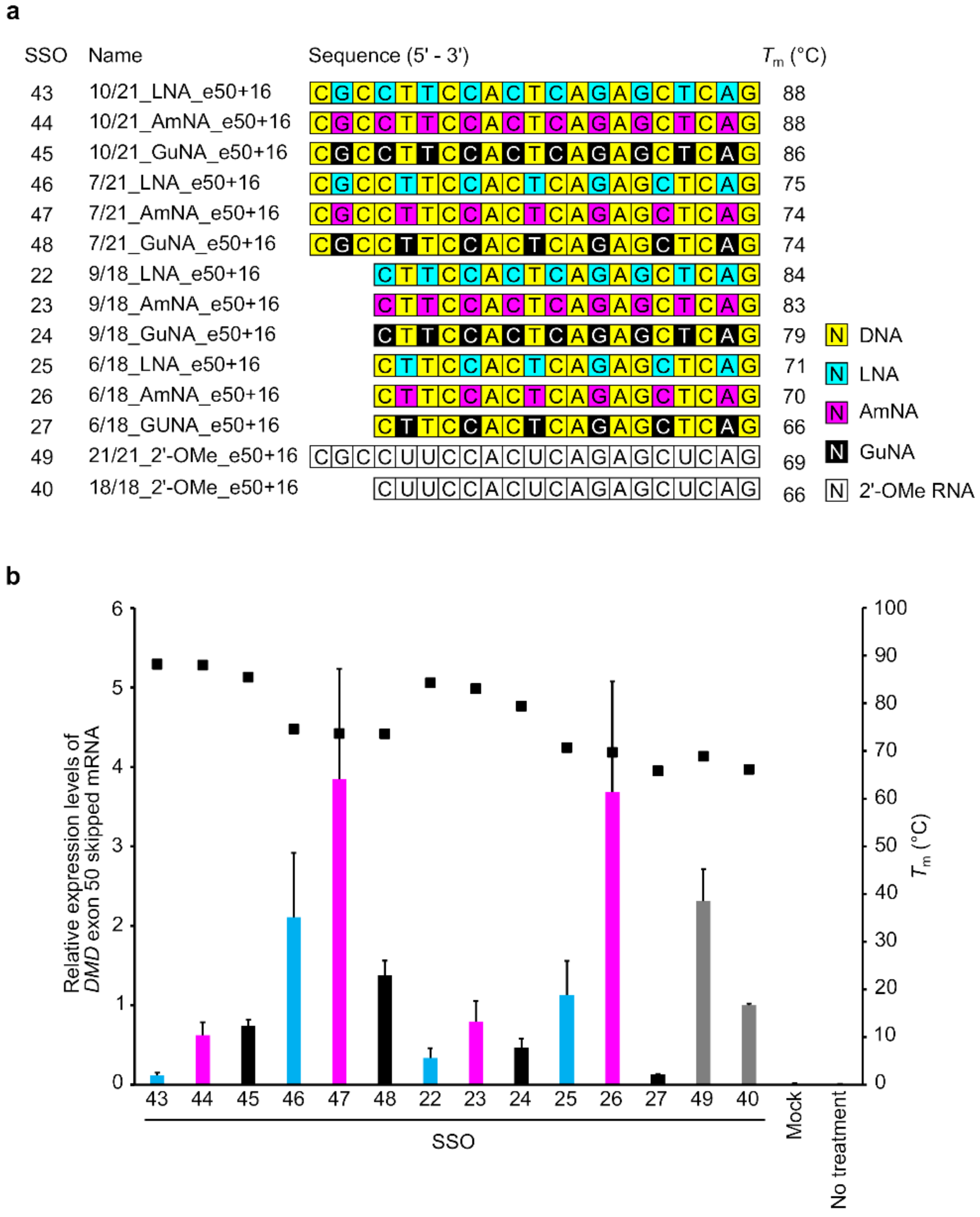
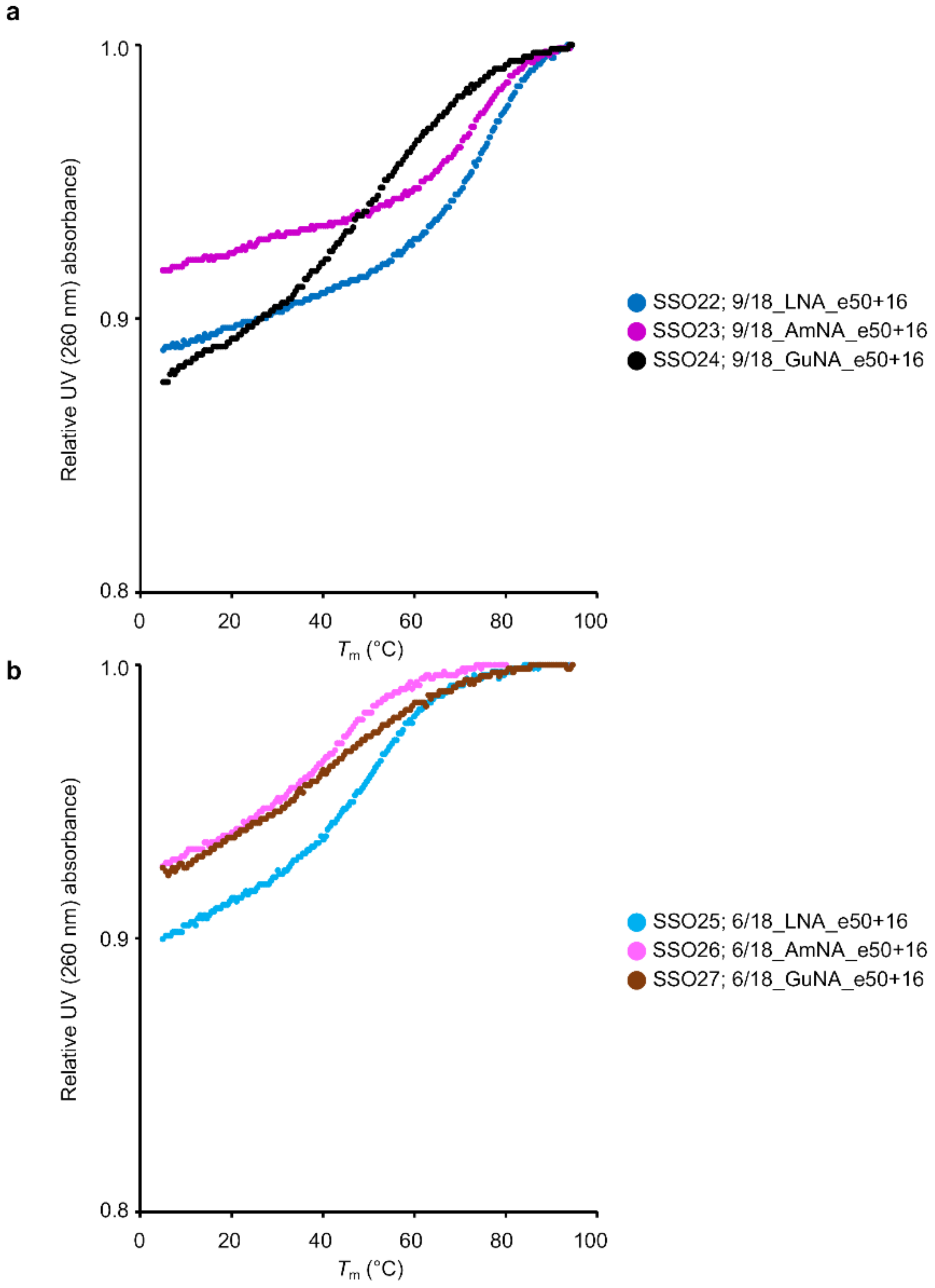
2.2. Confirmation of the General Design of BNA-Modified SSOs Targeting DMD Exon 50 Using a Stably Transfected Cell Line
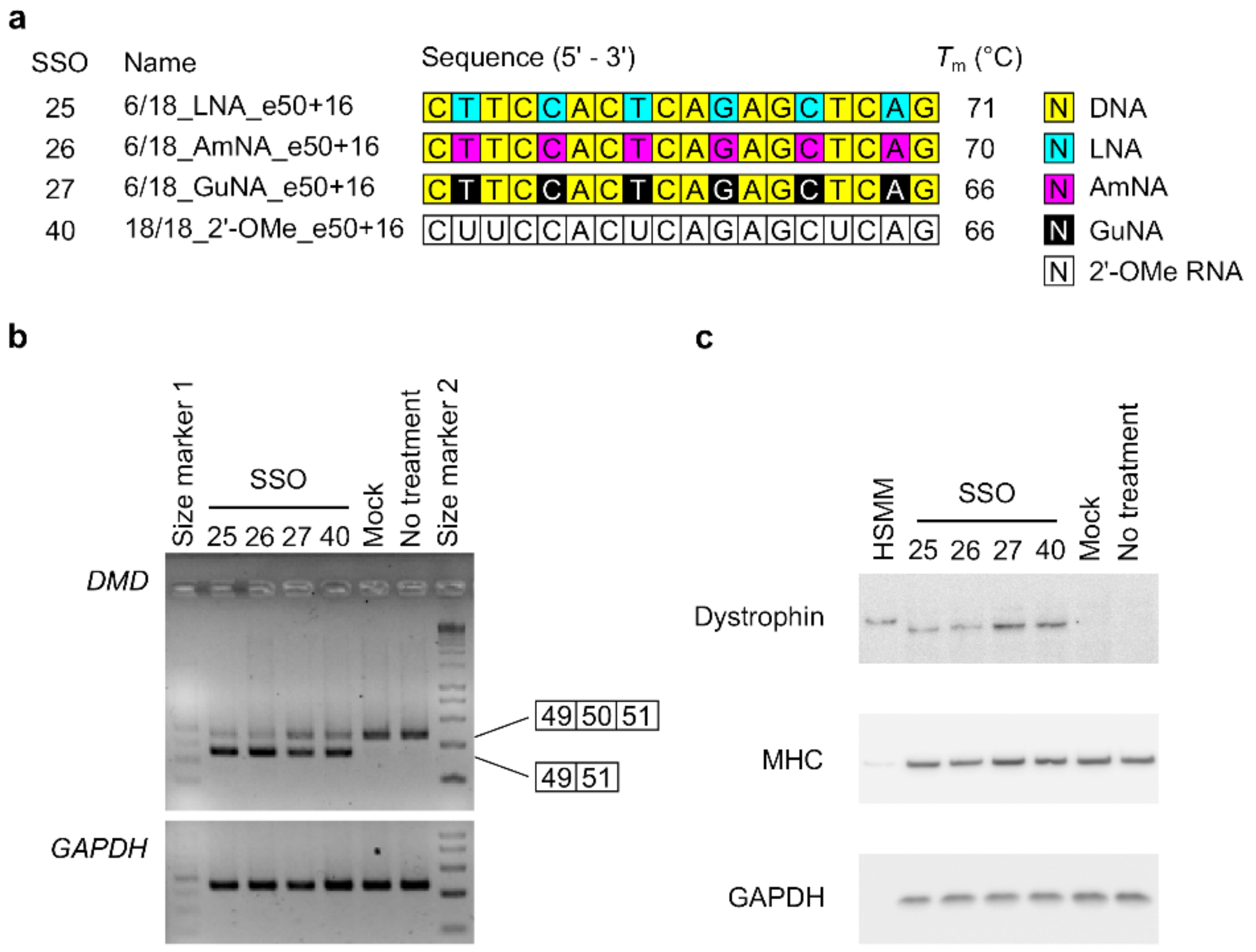
2.3. Investigating the Effect on Dystrophin Protein Restoration of BNA-Modified SSOs Using DMD Model Cells
3. Discussion
4. Materials and Methods
4.1. Synthesis of Oligonucleotides
4.2. UV Melting Analysis
4.3. Cell Culture
4.4. SSO Transfection
4.5. RNA Isolation and cDNA Synthesis
4.6. RT-PCR Analysis
4.7. Quantitative RT-PCR Analysis
4.8. Immunoblot Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hwang, J.; Yokota, T. Recent advancements in exon-skipping therapies using antisense oligonucleotides and genome editing for the treatment of various muscular dystrophies. Expert Rev. Mol. Med. 2019, 21, e5. [Google Scholar] [CrossRef]
- Havens, M.A.; Hastings, M.L. Splice-switching antisense oligonucleotides as therapeutic drugs. Nucleic Acids Res. 2016, 44, 6549–6563. [Google Scholar] [CrossRef]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in oligonucleotide drug delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Rigo, F.; Bennett, C.F.; Krainer, A.R.; Hua, Y. Comparison of the efficacy of MOE and PMO modifications of systemic antisense oligonucleotides in a severe SMA mouse model. Nucleic Acids Res. 2020, 48, 2853–2865. [Google Scholar] [CrossRef]
- Shimo, T.; Tachibana, K.; Saito, K.; Yoshida, T.; Tomita, E.; Waki, R.; Yamamoto, T.; Doi, T.; Inoue, T.; Kawakami, J.; et al. Design and evaluation of locked nucleic acid-based splice-switching oligonucleotides in vitro. Nucleic Acids Res. 2014, 42, 8174–8187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimo, T.; Tachibana, K.; Kawawaki, Y.; Watahiki, Y.; Ishigaki, T.; Nakatsuji, Y.; Hara, T.; Kawakami, J.; Obika, S. Enhancement of exon skipping activity by reduction in the secondary structure content of LNA-based splice-switching oligonucleotides. Chem. Commun. 2019, 55, 6850–6853. [Google Scholar] [CrossRef] [PubMed]
- Le, B.T.; Murayama, K.; Shabanpoor, F.; Asanuma, H.; Veedu, R.N. Antisense oligonucleotide modified with serinol nucleic acid (SNA) induces exon skipping in mdx myotubes. RSC Adv. 2017, 7, 34049–34052. [Google Scholar] [CrossRef] [Green Version]
- Le, B.T.; Adams, A.M.; Fletcher, S.; Wilton, S.D.; Veedu, R.N. Rational Design of Short Locked Nucleic Acid-Modified 2’-O-Methyl Antisense Oligonucleotides for Efficient Exon-Skipping In Vitro. Mol. Ther. Nucleic Acids 2017, 9, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Le, B.T.; Chakravarthy, M.; Kosbar, T.R.; Veedu, R.N. Systematic evaluation of 2’-Fluoro modified chimeric antisense oligonucleotide-mediated exon skipping in vitro. Sci. Rep. 2019, 9, 6078. [Google Scholar] [CrossRef]
- Le, B.T.; Chen, S.; Abramov, M.; Herdewijn, P.; Veedu, R.N. Evaluation of anhydrohexitol nucleic acid, cyclohexenyl nucleic acid and d-altritol nucleic acid-modified 2’-O-methyl RNA mixmer antisense oligonucleotides for exon skipping in vitro. Chem. Commun. 2016, 52, 13467–13470. [Google Scholar] [CrossRef]
- Roberts, J.; Palma, E.; Sazani, P.; Orum, H.; Cho, M.; Kole, R. Efficient and persistent splice switching by systemically delivered LNA oligonucleotides in mice. Mol. Ther. 2006, 14, 471–475. [Google Scholar] [CrossRef]
- Surono, A.; Van Khanh, T.; Takeshima, Y.; Wada, H.; Yagi, M.; Takagi, M.; Koizumi, M.; Matsuo, M. Chimeric RNA/ethylene-bridged nucleic acids promote dystrophin expression in myocytes of duchenne muscular dystrophy by inducing skipping of the nonsense mutation-encoding exon. Hum. Gene Ther. 2004, 15, 749–757. [Google Scholar] [CrossRef]
- Jarver, P.; O’Donovan, L.; Gait, M.J. A chemical view of oligonucleotides for exon skipping and related drug applications. Nucleic Acid Ther. 2014, 24, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Rigo, F.; Hua, Y.; Chun, S.J.; Prakash, T.P.; Krainer, A.R.; Bennett, C.F. Synthetic oligonucleotides recruit ILF2/3 to RNA transcripts to modulate splicing. Nat. Chem. Biol. 2012, 8, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, A.M.; Casper, M.D.; Freier, S.M.; Lesnik, E.A.; Zounes, M.C.; Cummins, L.L.; Gonzalez, C.; Cook, P.D. Uniformly modified 2’-deoxy-2’-fluoro phosphorothioate oligonucleotides as nuclease-resistant antisense compounds with high affinity and specificity for RNA targets. J. Med. Chem. 1993, 36, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Yahara, A.; Shrestha, A.R.; Yamamoto, T.; Hari, Y.; Osawa, T.; Yamaguchi, M.; Nishida, M.; Kodama, T.; Obika, S. Amido-bridged nucleic acids (AmNAs): Synthesis, duplex stability, nuclease resistance, and in vitro antisense potency. Chembiochem 2012, 13, 2513–2516. [Google Scholar] [CrossRef]
- Shrestha, A.R.; Kotobuki, Y.; Hari, Y.; Obika, S. Guanidine bridged nucleic acid (GuNA): An effect of a cationic bridged nucleic acid on DNA binding affinity. Chem. Commun. 2014, 50, 575–577. [Google Scholar] [CrossRef] [PubMed]
- Popplewell, L.J.; Trollet, C.; Dickson, G.; Graham, I.R. Design of phosphorodiamidate morpholino oligomers (PMOs) for the induction of exon skipping of the human DMD gene. Mol. Ther. 2009, 17, 554–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Disterer, P.; Kryczka, A.; Liu, Y.; Badi, Y.E.; Wong, J.J.; Owen, J.S.; Khoo, B. Development of therapeutic splice-switching oligonucleotides. Hum. Gene. Ther. 2014, 25, 587–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popplewell, L.J.; Malerba, A.; Dickson, G. Optimizing antisense oligonucleotides using phosphorodiamidate morpholino oligomers. Methods Mol. Biol. 2012, 867, 143–167. [Google Scholar]
- Shimo, T.; Obika, S. Optimization of 2’,4’-BNA/LNA-Based Oligonucleotides for Splicing Modulation In Vitro. Methods Mol. Biol. 2018, 1828, 395–411. [Google Scholar]
- Graziewicz, M.A.; Tarrant, T.K.; Buckley, B.; Roberts, J.; Fulton, L.; Hansen, H.; Orum, H.; Kole, R.; Sazani, P. An Endogenous TNF-alpha Antagonist Induced by Splice-switching Oligonucleotides Reduces Inflammation in Hepatitis and Arthritis Mouse Models. Mol. Ther. 2008, 16, 1316–1322. [Google Scholar] [CrossRef]
- Bestas, B.; Moreno, P.M.; Blomberg, K.E.; Mohammad, D.K.; Saleh, A.F.; Sutlu, T.; Nordin, J.Z.; Guterstam, P.; Gustafsson, M.O.; Kharazi, S. Splice-correcting oligonucleotides restore BTK function in X-linked agammaglobulinemia model. J. Clin. Investig. 2014, 124, 4067–4081. [Google Scholar] [CrossRef] [Green Version]
- Aartsma-Rus, A.; De Winter, C.L.; Janson, A.A.; Kaman, W.E.; Van Ommen, G.J.; Den Dunnen, J.T.; Van Deutekom, J.C. Functional analysis of 114 exon-internal AONs for targeted DMD exon skipping: Indication for steric hindrance of SR protein binding sites. Oligonucleotides 2005, 15, 284–297. [Google Scholar] [CrossRef]
- Wu, B.; Lu, P.; Cloer, C.; Shaban, M.; Grewal, S.; Milazi, S.; Shah, S.N.; Moulton, H.M.; Lu, Q.L. Long-term rescue of dystrophin expression and improvement in muscle pathology and function in dystrophic mdx mice by peptide-conjugated morpholino. Am. J. Pathol. 2012, 181, 392–400. [Google Scholar] [CrossRef] [Green Version]
- Shimo, T.; Hosoki, K.; Nakatsuji, Y.; Yokota, T.; Obika, S. A novel human muscle cell model of Duchenne muscular dystrophy created by CRISPR/Cas9 and evaluation of antisense-mediated exon skipping. J. Hum. Genet. 2018, 63, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Scharner, J.; Ma, W.K.; Zhang, Q.; Lin, K.T.; Rigo, F.; Bennett, C.F.; Krainer, A.R. Hybridization-mediated off-target effects of splice-switching antisense oligonucleotides. Nucleic Acids Res. 2020, 48, 802–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, V.B.; Simoes, R.; Mamchaoui, K.; Carvalho, C.; Carmo-Fonseca, M. Short (16-mer) locked nucleic acid splice-switching oligonucleotides restore dystrophin production in Duchenne Muscular Dystrophy myotubes. PLoS ONE 2017, 12, e0181065. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Yahara, A.; Waki, R.; Yasuhara, H.; Wada, F.; Harada-Shiba, M.; Obika, S. Amido-bridged nucleic acids with small hydrophobic residues enhance hepatic tropism of antisense oligonucleotides in vivo. Org. Biomol. Chem. 2015, 13, 3757–3765. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, S.; Sawamoto, H.; Takegawa-Araki, T.; Arai, Y.; Yamakoshi, S.; Yamada, K.; Ohta, T.; Kawanishi, E.; Horie, N.; Yamaguchi, T.; et al. Synthesis and properties of GuNA purine/pyrimidine nucleosides and oligonucleotides. Org. Biomol. Chem. 2020, 18, 9461–9472. [Google Scholar] [CrossRef] [PubMed]
- Shimo, T.; Tachibana, K.; Obika, S. Construction of a tri-chromatic reporter cell line for the rapid and simple screening of splice-switching oligonucleotides targeting DMD exon 51 using high content screening. PLoS ONE 2018, 13, e0197373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of BNA | Target Exon | Minimal Length for Effective Exon Skipping | Tm (°C) for Effective Exon Skipping | Key Features |
|---|---|---|---|---|
| LNA | DMD exon 58 | 13 | 72 to 85 | – |
| DMD exon 50 | 18 | 71 to 75 | ||
| AmNA | DMD exon 58 | 13 | 71 to 85 | Higher activity than the corresponding LNA-modified SSOs |
| DMD exon 50 | 15 | 65 to 78 | ||
| GuNA | DMD exon 58 | 15 | 79 to 82 | Longer SSO is more active |
| DMD exon 50 | 21 | 74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimo, T.; Nakatsuji, Y.; Tachibana, K.; Obika, S. Design and In Vitro Evaluation of Splice-Switching Oligonucleotides Bearing Locked Nucleic Acids, Amido-Bridged Nucleic Acids, and Guanidine-Bridged Nucleic Acids. Int. J. Mol. Sci. 2021, 22, 3526. https://doi.org/10.3390/ijms22073526
Shimo T, Nakatsuji Y, Tachibana K, Obika S. Design and In Vitro Evaluation of Splice-Switching Oligonucleotides Bearing Locked Nucleic Acids, Amido-Bridged Nucleic Acids, and Guanidine-Bridged Nucleic Acids. International Journal of Molecular Sciences. 2021; 22(7):3526. https://doi.org/10.3390/ijms22073526
Chicago/Turabian StyleShimo, Takenori, Yusuke Nakatsuji, Keisuke Tachibana, and Satoshi Obika. 2021. "Design and In Vitro Evaluation of Splice-Switching Oligonucleotides Bearing Locked Nucleic Acids, Amido-Bridged Nucleic Acids, and Guanidine-Bridged Nucleic Acids" International Journal of Molecular Sciences 22, no. 7: 3526. https://doi.org/10.3390/ijms22073526
APA StyleShimo, T., Nakatsuji, Y., Tachibana, K., & Obika, S. (2021). Design and In Vitro Evaluation of Splice-Switching Oligonucleotides Bearing Locked Nucleic Acids, Amido-Bridged Nucleic Acids, and Guanidine-Bridged Nucleic Acids. International Journal of Molecular Sciences, 22(7), 3526. https://doi.org/10.3390/ijms22073526