
Altered Task-Evoked Corticolimbic Responsivity in Generalized Anxiety Disorder

{kind=link}
Abstract
:1. Introduction
2. Neural Circuitry for Emotional Reactivity and Regulation
2.1. Neuroanatomy of the Corticolimbic Circuit
2.2. Corticolimbic Circuit and Anxiety
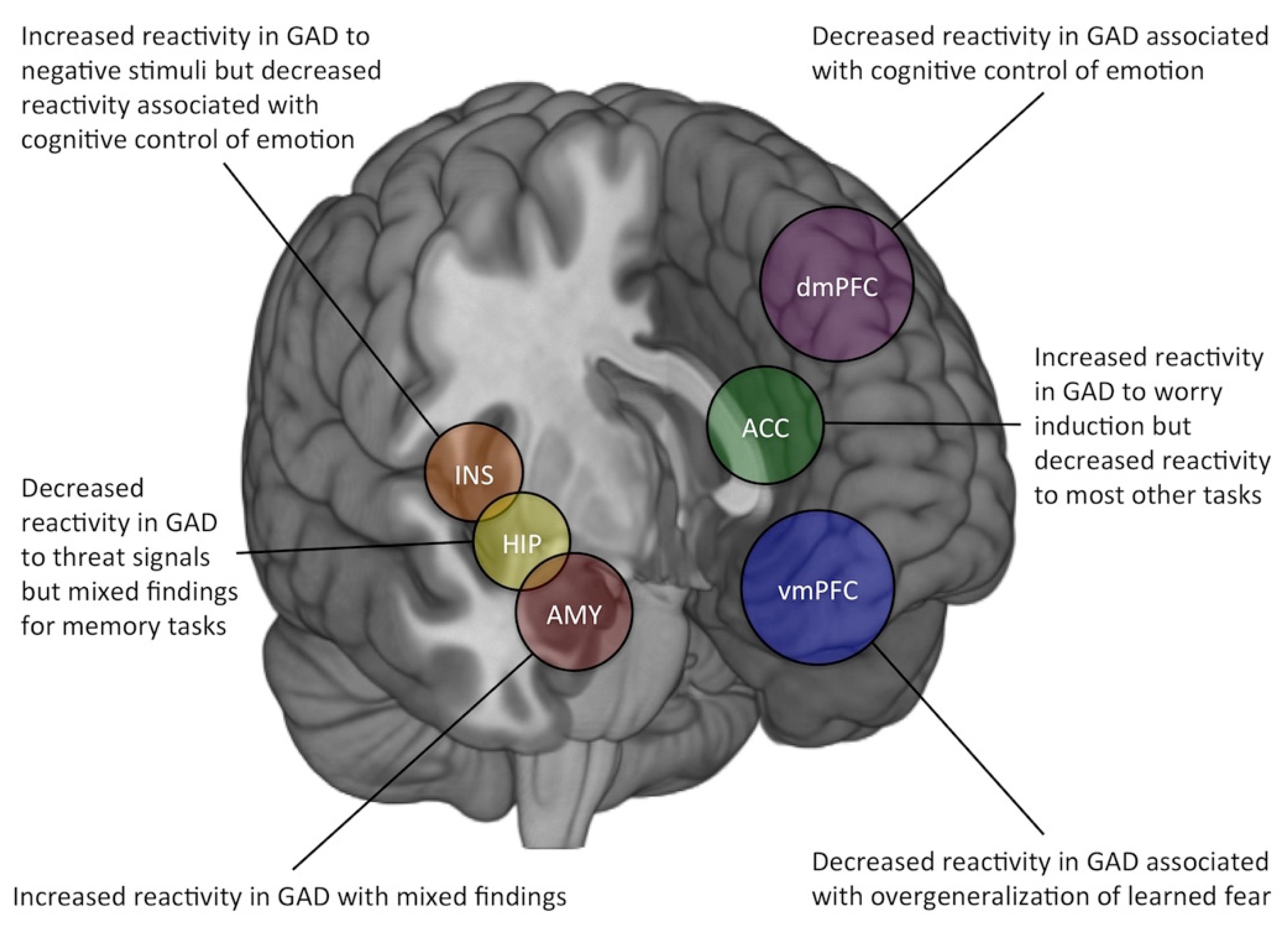
3. Altered BOLD Responsivity in GAD
3.1. Amygdala
3.2. Anterior Cingulate Cortex
3.3. Prefrontal Cortex
3.4. Other Brain Regions
4. Factors Contributing to the Mixed Findings in Task-Based fMRI Studies of GAD
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Comer, J.S.; Kendall, P.C.; Franklin, M.E.; Hudson, J.L.; Pimentel, S.S. Obsessing/worrying about the overlap between obsessive-compulsive disorder and generalized anxiety disorder in youth. Clin. Psychol. Rev. 2004, 24, 663–683. [Google Scholar] [CrossRef]
- Mineka, S.; Hendersen, R.W. Controllability and predictability in acquired motivation. Annu. Rev. Psychol. 1985, 36, 495–529. [Google Scholar] [CrossRef]
- Ruscio, A.M.; Gentes, E.L.; Jones, J.D.; Hallion, L.S.; Coleman, E.S.; Swendsen, J. Rumination predicts heightened responding to stressful life events in major depressive disorder and generalized anxiety disorder. J. Abnorm. Psychol. 2015, 124, 17–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabat-Zinn, J.; Massion, A.O.; Kristeller, J.; Peterson, L.G.; Fletcher, K.E.; Pbert, L.; Lenderking, W.R.; Santorelli, S.F. Effectiveness of a meditation-based stress reduction program in the treatment of anxiety disorder. Am. J. Psychiatry 1992, 149, 936–943. [Google Scholar] [PubMed]
- Zugman, A.; Harrewijn, A.; Cardinale, E.M.; Zwiebel, H.; Freitag, G.F.; Werwath, K.E.; Bas-Hoogendam, J.M.; Groenewold, N.A.; Aghajani, M.; Hilbert, K.; et al. Mega-analysis methods in ENIGMA: The experience of the generalized anxiety disorder working group. Hum. Brain Mapp. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kolesar, T.; Bilevicius, E.; Wilson, A.D.; Kornelsen, J. Systematic review and meta-analyses of neural structural and functional differences in generalized anxiety disorder and healthy controls using magnetic resonance imaging. NeuroImage Clin. 2019, 24, 102016. [Google Scholar] [CrossRef] [PubMed]
- Madonna, D.; Delvecchio, G.; Soares, J.C.; Brambilla, P. Structural and functional neuroimaging studies in generalized anxiety disorder: A systematic review. Braz. J. Psychiatry 2019, 41, 336–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szucs, D.; Ioannidis, J.P. Sample size evolution in neuroimaging research: An evaluation of highly-cited studies (1990-2012) and of latest practices (2017-2018) in high-impact journals. NeuroImage 2020, 221, 117164. [Google Scholar] [CrossRef] [PubMed]
- Elliott, M.L.; Knodt, A.R.; Ireland, D.; Morris, M.L.; Poulton, R.; Ramrakha, S.; Sison, M.L.; Moffitt, T.E.; Caspi, A.; Hariri, A.R. What is the test-retest reliability of common task-functional MRI measures? New empirical evidence and a meta-analysis. Psychol. Sci. 2020, 31, 792–806. [Google Scholar] [CrossRef]
- Infantolino, Z.P.; Luking, K.R.; Sauder, C.K.; Curtin, J.J.; Hajcak, G. Robust is not necessarily reliable: From within-subjects fMRI contrasts to between-subjects comparisons. NeuroImage 2018, 173, 146–152. [Google Scholar] [CrossRef]
- Bishop, S.J. Neurocognitive mechanisms of anxiety: An integrative account. Trends Cogn. Sci. 2007, 11, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Grupe, D.W.; Nitschke, J.B. Uncertainty and anticipation in anxiety: An integrated neurobiological and psychological perspective. Nat. Rev. Neurosci. 2013, 14, 488–501. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Loucks, R.A.; Palmer, A.L.; Brown, A.C.; Solomon, K.M.; Marchante, A.N.; Whalen, P.J. The structural and functional connectivity of the amygdala: From normal emotion to pathological anxiety. Behav. Brain Res. 2011, 223, 403–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggleton, J.P. The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction; Wiley-Liss, Inc.: New York, NY, USA, 1992. [Google Scholar]
- Whalen, P.J.; Phelps, E.A. The Human Amygdala; The Guilford Press: New York, NY, USA, 2009. [Google Scholar]
- LeDoux, J.E. Emotion circuits in the brain. Annu. Rev. Neurosci. 2000, 23, 155–184. [Google Scholar] [CrossRef] [PubMed]
- Adolphs, R.; Tranel, D.; Damasio, H.; Damasio, A.R. Fear and the human amygdala. J. Neurosci. 1995, 15, 5879–5891. [Google Scholar] [CrossRef]
- Davis, M.; Whalen, P.J. The amygdala: Vigilance and emotion. Mol. Psychiatry 2001, 6, 13–34. [Google Scholar] [CrossRef] [Green Version]
- Hariri, A.R.; Mattay, V.S.; Tessitore, A.; Kolachana, B.; Fera, F.; Goldman, D.; Egan, M.F.; Weinberger, D.R. Serotonin transporter genetic variation and the response of the human amygdala. Science 2002, 297, 400–403. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Zelano, C.; Gottfried, J.A.; Mohanty, A. Human amygdala represents the complete spectrum of subjective valence. J. Neurosci. 2015, 35, 15145–15156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson-Mendenhall, C.D.; Barrett, L.F.; Barsalou, L.W. Neural evidence that human emotions share core affective properties. Psychol. Sci. 2013, 24, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Adolphs, R. What does the amygdala contribute to social cognition? Ann. N. Y. Acad. Sci. 2010, 1191, 42–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghashghaei, H.T.; Hilgetag, C.C.; Barbas, H. Sequence of information processing for emotions based on the anatomic dialogue between prefrontal cortex and amygdala. NeuroImage 2007, 34, 905–923. [Google Scholar] [CrossRef] [Green Version]
- Milad, M.R.; Quirk, G.J. Neurons in medial prefrontal cortex signal memory for fear extinction. Nature 2002, 420, 70–74. [Google Scholar] [CrossRef]
- McRae, K.; Gross, J.J. Emotion regulation. Emotion 2020, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, K.C.; Jazaieri, H.; Gross, J.J. Emotion regulation: A transdiagnostic perspective on a new RDoC domain. Cognit. Ther. Res. 2016, 40, 426–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morawetz, C.; Riedel, M.C.; Salo, T.; Berboth, S.; Eickoff, S.B.; Laird, A.R.; Kohn, N. Multiple large-scale neural networks underlying emotion regulation. Neurosci. Biobehav. Rev. 2020, 116, 382–395. [Google Scholar] [CrossRef]
- Ochsner, K.N.; Gross, J.J. The cognitive control of emotion. Trends Cogn. Sci. 2005, 9, 242–249. [Google Scholar] [CrossRef]
- Denny, B.T.; Inhoff, M.C.; Zerubavel, N.; Davachi, L.; Ocshner, K.N. Getting over it: Long-lasting effects of emotion regulation on amygdala response. Psychol. Sci. 2015, 26, 1377–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhle, J.T.; Silvers, J.A.; Wager, T.D.; Lopez, R.; Onyemekwu, C.; Kober, H.; Weber, J.; Ochsner, K.N. Cognitive reappraisal of emotion: A meta-analysis of human neuroimaging studies. Cereb. Cortex 2014, 24, 2981–2990. [Google Scholar] [CrossRef] [PubMed]
- Hartley, C.A.; Phelps, E.A. Changing fear: The neurocircuitry of emotion regulation. Neuropsychopharmacology 2009, 35, 136–146. [Google Scholar] [CrossRef] [Green Version]
- Quirk, G.J.; Beer, J.S. Prefrontal involvement in the regulation of emotion: Convergence of rat and human studies. Curr. Opin. Neurobiol. 2006, 16, 723–727. [Google Scholar] [CrossRef]
- Barbas, H.; De Olmos, J. Projections from the amygdala to basoventral and mediodorsal prefrontal regions in the rhesus monkey. J. Comp. Neurol. 1990, 300, 549–571. [Google Scholar] [CrossRef]
- Carmichael, S.T.; Price, J.L. Limbic connections of the orbital and medial prefrontal cortex in macaque monkeys. J. Comp. Neurol. 1995, 363, 615–641. [Google Scholar] [CrossRef]
- Aggleton, J.P.; Burton, M.J.; Passingham, R.E. Cortical and subcortical afferents to the amygdala of the rhesus monkey (Macaca mulatta). Brain Res. 1980, 190, 347–368. [Google Scholar] [CrossRef]
- Calhoon, G.G.; Tye, K.M. Resolving the neural circuits of anxiety. Nat. Neurosci. 2015, 18, 1394–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tovote, P.; Fadok, J.P.; Lüthi, A. Neuronal circuits for fear and anxiety. Nat. Rev. Neurosci. 2015, 16, 317–331. [Google Scholar] [CrossRef]
- Etkin, A.; Wager, T.D. Functional neuroimaging of anxiety: A meta-analysis of emotional processing in PTSD, social anxiety disorder, and specific phobia. Am. J. Psychiatry 2007, 164, 1476–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, L.M.; Liberzon, I. The neurocircuitry of fear, stress, and anxiety. Neuropsychopharmacology 2010, 35, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Bishop, S.; Duncan, J.; Brett, M.; Lawrence, A.D. Prefrontal cortical function and anxiety: Controlling attention to threat-related stimuli. Nat. Neurosci. 2004, 7, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Chey, J.; Chung, A.; Bae, S.; Khang, H.; Ham, B.; Yoon, S.J.; Jeong, D.-U.; Lyoo, I.K. Diminished rostral anterior cingulate activity in response to threat-related events in posttraumatic stress disorder. J. Psychiatry Res. 2008, 42, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Hariri, A.R.; Mattay, V.S.; Tessitore, A.; Fera, F.; Weinberger, D.R. Neocortical modulation of the amygdala response to fearful stimuli. Biol. Psychiatry 2003, 53, 494–501. [Google Scholar] [CrossRef]
- Pezawas, L.; Meyer-Lindenberg, A.; Drabant, E.M.; Verchinski, B.A.; Munoz, K.E.; Kolachana, B.S.; Egan, M.F.; Mattay, V.S.; Hariri, A.R.; Weinberger, D.R. 5-HTTLPR polymorphism impacts human cingulate-amygdala interactions: A genetic susceptibility mechanism for depression. Nat. Neurosci. 2005, 8, 828–834. [Google Scholar] [CrossRef]
- Fitzgerald, J.M.; Phan, K.L.; Kennedy, A.E.; Shankman, S.A.; Langenecker, S.A.; Klumpp, H. Prefrontal and amygdala engagement during emotional reactivity and regulation in generalized anxiety disorder. J. Affect. Disord. 2017, 218, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Price, R.B.; Eldreth, D.A.; Mohlman, J. Deficient prefrontal attentional control in late-life generalized anxiety disorder: An fMRI investigation. Transl. Psychiatry 2011, 1, e46. [Google Scholar] [CrossRef] [Green Version]
- Mohlman, J.; Eldreth, D.A.; Price, R.B.; Staples, A.M.; Hanson, C. Prefrontal-limbic connectivity during worry in older adults with generalized anxiety disorder. Aging Ment. Health 2017, 21, 426–438. [Google Scholar] [CrossRef]
- Buff, C.; Schmidt, C.; Brinkmann, L.; Gathmann, B.; Tupak, S.; Straube, T. Directed threat imagery in generalized anxiety disorder. Psychol. Med. 2018, 48, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Nitschke, J.B.; Sarinopoulos, I.; Oathes, D.J.; Johnstone, T.; Whalen, P.J.; Davidson, R.J.; Kalin, N.H. Anticipatory activation in the amygdala and anterior cingulate in generalized anxiety disorder and prediction of treatment response. Am. J. Psychiatry 2009, 166, 302–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, K.S.; Geraci, M.; Smith, B.W.; Hollon, N.; DeVido, J.; Otero, M.; Pine, D.S. Reduced dorsal anterior cingulate cortical activity during emotional regulation and top-down attentional control in generalized social phobia, generalized anxiety disorder, and comorbid generalized social phobia/generalized anxiety disorder. Biol. Psychiatry 2012, 72, 476–482. [Google Scholar] [CrossRef] [Green Version]
- Carlson, J.M.; Rubin, D.; Mujica-Parodi, L.R. Lost emotion: Disrupted brain-based tracking of dynamic affective episodes in anxiety and depression. Psychiatry Res. Neuroimaging 2017, 260, 37–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yassa, M.A.; Hazlett, R.L.; Stark, C.E.L.; Hoehn-Saric, R. Functional MRI of the amygdala and bed nucleus of the stria terminalis during conditions of uncertainty in generalized anxiety disorder. J. Psychiatr. Res. 2012, 46, 1045–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClure, E.B.; Monk, C.S.; Nelson, E.E.; Parrish, J.M.; Adler, A.; Blair, R.J.R.; Fromm, S.; Charney, D.S.; Leibenluft, E.; Ernst, M.; et al. Abnormal attention modulation of fear circuit function in pediatric generalized anxiety disorder. Arch. Gen. Psychiatry 2007, 64, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Monk, C.S.; Telzer, E.H.; Mogg, K.; Bradley, B.P.; Mai, X.; Louro, H.M.C.; Chen, G.; McClure, E.B.; Ernst, M.; Pine, D.S. Amygdala and ventrolateral prefrontal cortex activation to masked angry faces in children and adolescents with generalized anxiety disorder. Arch. Gen. Psychiatry 2008, 65, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Fonzo, G.A.; Ramsawh, H.J.; Flagan, T.M.; Sullivan, S.G.; Simmons, A.N.; Paulus, M.P.; Stein, M.B. Cognitive-behavioral therapy for generalized anxiety disorder is associated with attenuation of limbic activation to threat-related facial emotions. J. Affect. Disord. 2014, 169, 76–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonzo, G.A.; Ramsawh, H.J.; Flagan, T.M.; Sullivan, S.G.; Letamendi, A.; Simmons, A.N.; Paulus, M.P.; Stein, M.B. Common and disorder-specific neural responses to emotional faces in generalised anxiety, social anxiety and panic disorders. Br. J. Psychiatry 2015, 206, 206–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hariri, A.R.; Whalen, P.J. The amygdala: Inside and out. F1000 Biol. Rep. 2011, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Whalen, P.J. Fear, vigilance, and ambiguity: Initial neuroimaging studies of the human amygdala. Curr. Dir. Psychol. Sci. 1998, 7, 177–188. [Google Scholar] [CrossRef]
- Hölzel, B.K.; Hoge, E.A.; Greve, D.N.; Gard, T.; Creswell, J.D.; Brown, K.W.; Barrett, L.F.; Schwartz, C.; Vaitl, D.; Lazar, S.W. Neural mechanisms of symptom improvements in generalized anxiety disorder following mindfulness training. NeuroImage Clin. 2013, 2, 448–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, K.; Shaywitz, J.; Smith, B.W.; Rhodes, R.; Geraci, M.; Jones, M.; McCaffrey, D.; Vythilingam, M.; Finger, E.; Mondillo, K.; et al. Response to emotional expressions in generalized social phobia and generalized anxiety disorder: Evidence for separate disorders. Am. J. Psychiatry 2008, 165, 1193–1202. [Google Scholar] [CrossRef] [Green Version]
- Etkin, A.; Egner, T.; Peraza, D.M.; Kandel, E.R.; Hirsch, J. Resolving emotional conflict: A role for the rostral anterior cingulate cortex in modulating activity in the amygdala. Neuron 2006, 51, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Bush, G.; Luu, P.; Posner, M.I. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn. Sci. 2000, 4, 215–222. [Google Scholar] [CrossRef]
- Li, J.; Zhong, Y.; Ma, Z.; Wu, Y.; Pang, M.; Wang, C.; Liu, N.; Wang, C.; Zhang, N. Emotion reactivity-related brain network analysis in generalized anxiety disorder: A task fMRI study. BMC Psychiatry 2020, 20, 429. [Google Scholar] [CrossRef]
- Palm, M.E.; Elliott, R.; McKie, S.; Deakin, J.F.W.; Anderson, I.M. Attenuated responses to emotional expressions in women with generalized anxiety disorder. Psychol. Med. 2011, 41, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Ball, T.M.; Ramsawh, H.J.; Campbell-Sills, L.; Paulus, M.P.; Stein, M.B. Prefrontal dysfunction during emotion regulation in generalized anxiety and panic disorders. Psychol. Med. 2013, 43, 1475–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Via, E.; Fullana, M.A.; Goldberg, X.; Tinoco-González, D.; Martínez-Zalacaín, I.; Soriano-Mas, C.; Davey, C.G.; Menchón, J.M.; Straube, B.; Kircher, T.; et al. Ventromedial prefrontal cortex activity and pathological worry in generalised anxiety disorder. Br. J. Psychiatry 2018, 213, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Paulesu, E.; Sambugaro, E.; Torti, T.; Danelli, L.; Ferri, F.; Scialfa, G.; Sberna, M.; Ruggiero, G.M.; Bottini, G.; Sassaroli, S. Neural correlates of worry in generalized anxiety disorder and in normal controls: A functional MRI study. Psychol. Med. 2010, 40, 117–124. [Google Scholar] [CrossRef]
- Andreescu, C.; Gross, J.J.; Lenze, E.; Edelman, K.D.; Snyder, S.; Tanase, C.; Aizenstein, H. Altered cerebral blood flow patterns associated with pathologic worry in the elderly. Depress. Anxiety 2011, 28, 202–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable intrinsic connectivity networks for salience processing and executive control. J. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef]
- Cha, J.; Greenberg, T.; Carlson, J.M.; DeDora, D.J.; Hajcak, G.; Mujica-Parodi, L.R. Circuit-wide structural and functional measures predict ventromedial prefrontal cortex fear generalization: Implications for generalized anxiety disorder. J. Neurosci. 2014, 34, 4043–4053. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, T.; Carlson, J.M.; Cha, J.; Hajcak, G.; Mujica-Parodi, L.R. Ventromedial prefrontal cortex reactivity is altered in generalized anxiety disorder during fear generalization. Depress. Anxiety 2013, 30, 242–250. [Google Scholar] [CrossRef]
- Diwadkar, V.A.; Re, M.; Cecchetto, F.; Garzitto, M.; Piccin, S.; Bonivento, C.; Maieron, M.; D’Agostini, S.; Balestreri, M.; Brambilla, P. Attempts at memory control induce dysfunctional brain activation profiles in generalized anxiety disorder: An exploratory fMRI study. Psychiatry Res. Neuroimaging 2017, 266, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Etkin, A.; Prater, K.E.; Hoeft, F.; Menon, V.; Schatzberg, A.F. Failure of anterior cingulate activation and connectivity with the amygdala during implicit regulation of emotional processing in generalized anxiety disorder. Am. J. Psychiatry 2010, 167, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Moon, C.M.; Jeong, G.W. Functional and morphological alterations associated with working memory dysfunction in patients with generalized anxiety disorder. Acta Radiol. 2017, 58, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.M.; Jeong, G.W. Functional neuroanatomy on the working memory under emotional distraction in patients with generalized anxiety disorder. Psychiatry Clin. Neurosci. 2015, 69, 609–619. [Google Scholar] [CrossRef]
- Moon, C.M.; Sundaram, T.; Choi, N.G.; Jeong, G.W. Working memory dysfunction associated with brain functional deficits and cellular metabolic changes in patients with generalized anxiety disorder. Psychiatry Res. Neuroimaging 2016, 254, 137–144. [Google Scholar] [CrossRef]
- Blair, K.S.; Otero, M.; Teng, C.; Geraci, M.; Ernst, M.; Blair, R.J.R.; Pine, D.S.; Grillon, C. Reduced optimism and a heightened neural response to everyday worries are specific to generalized anxiety disorder, and not seen in social anxiety. Psychol. Med. 2017, 47, 1806–1815. [Google Scholar] [CrossRef] [PubMed]
- Buff, C.; Brinkmann, L.; Neumeister, P.; Feldker, K.; Heitmann, C.; Gathmann, B.; Andor, T.; Straube, T. Specifically altered brain responses to threat in generalized anxiety disorder relative to social anxiety disorder and panic disorder. NeuroImage Clin. 2016, 12, 698–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, C.M.; Yang, J.C.; Jeong, G.W. Functional neuroanatomy associated with the interaction between emotion and cognition in explicit memory tasks in patients with generalized anxiety disorder. Acta Radiol. 2017, 58, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.M.; Yang, J.C.; Jeong, G.W. Explicit verbal memory impairments associated with brain functional deficits and morphological alterations in patients with generalized anxiety disorder. J. Affect. Disord. 2015, 186, 328–336. [Google Scholar] [CrossRef]
- Strawn, J.R.; Bitter, S.M.; Weber, W.A.; Chu, W.J.; Whitsel, R.M.; Adler, C.; Cerullo, M.A.; Eliassen, J.; Strakowski, S.M.; Delbello, M.P. Neurocircuitry of generalized anxiety disorder in adolescents: A pilot functional neuroimaging and functional connectivity study. Depress. Anxiety 2012, 29, 939–947. [Google Scholar] [CrossRef]
- Monk, C.S.; Nelson, E.E.; McClure, E.B.; Mogg, K.; Bradley, B.P.; Leibenluft, E.; Blair, R.J.R.; Chen, G.; Charney, D.S.; Ernst, M.; et al. Ventrolateral prefrontal cortex activation and attentional bias in response to angry faces in adolescents with generalized anxiety disorder. Am. J. Psychiatry 2006, 163, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Zhang, B.; Li, W.; Li, H.; Pang, J.; Hu, Q.; Zhang, L.; Tang, Y.; Yang, Z.; Wang, J.; et al. Insula shows abnormal task-evoked and resting-state activity in first-episode drug-naïve generalized anxiety disorder. Depress. Anxiety 2020, 37, 632–644. [Google Scholar] [CrossRef]
- Tan, Y.; Wei, D.; Zhang, M.; Yang, J.; Jelincic, V.; Qiu, J. The role of mid-insula in the relationship between cardiac interoceptive attention and anxiety: Evidence from an fMRI study. Sci. Rep. 2018, 8, 17280. [Google Scholar] [CrossRef] [PubMed]
- Laufer, O.; Israeli, D.; Paz, R. Behavioral and neural mechanisms of overgeneralization in anxiety. Curr. Biol. 2016, 26, 713–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, J.; Greenberg, T.; Song, I.; Blair Simpson, H.; Posner, J.; Mujica-Parodi, L.R. Abnormal hippocampal structure and function in clinical anxiety and comorbid depression. Hippocampus 2016, 26, 545–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, J.; Adolphs, R. Building a science of individual differences from fMRI. Trends Cogn. Sci. 2016, 20, 425–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grady, C.L.; Rieck, J.R.; Nichol, D.; Rodrigue, K.M.; Kennedy, K.M. Influence of sample size and analytic approach on stability and interpretation of brain-behavior correlations in task-related fMRI data. Hum. Brain Mapp. 2021, 42, 204–219. [Google Scholar] [CrossRef]
- Turner, B.O.; Paul, E.J.; Miller, M.B.; Barbey, A.K. Small sample sizes reduce the replicability of task-based fMRI studies. Commun. Biol. 2018, 1, 62. [Google Scholar] [CrossRef] [Green Version]
- Kragel, P.A.; Han, X.; Kraynak, T.E.; Gianaros, P.J.; Wager, T.D. Functional MRI can be highly reliable, but it depends on what you measure: A Commentary on Elliott et al. (2020). Psychol. Sci. 2021. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, N.; Kim, M.J. Altered Task-Evoked Corticolimbic Responsivity in Generalized Anxiety Disorder. Int. J. Mol. Sci. 2021, 22, 3630. https://doi.org/10.3390/ijms22073630
Kim N, Kim MJ. Altered Task-Evoked Corticolimbic Responsivity in Generalized Anxiety Disorder. International Journal of Molecular Sciences. 2021; 22(7):3630. https://doi.org/10.3390/ijms22073630
Chicago/Turabian StyleKim, Nayoung, and M. Justin Kim. 2021. "Altered Task-Evoked Corticolimbic Responsivity in Generalized Anxiety Disorder" International Journal of Molecular Sciences 22, no. 7: 3630. https://doi.org/10.3390/ijms22073630
APA StyleKim, N., & Kim, M. J. (2021). Altered Task-Evoked Corticolimbic Responsivity in Generalized Anxiety Disorder. International Journal of Molecular Sciences, 22(7), 3630. https://doi.org/10.3390/ijms22073630