
CD34+ Stromal Cells/Telocytes as a Source of Cancer-Associated Fibroblasts (CAFs) in Invasive Lobular Carcinoma of the Breast

 ,
, 
Abstract
:1. Introduction
2. Results
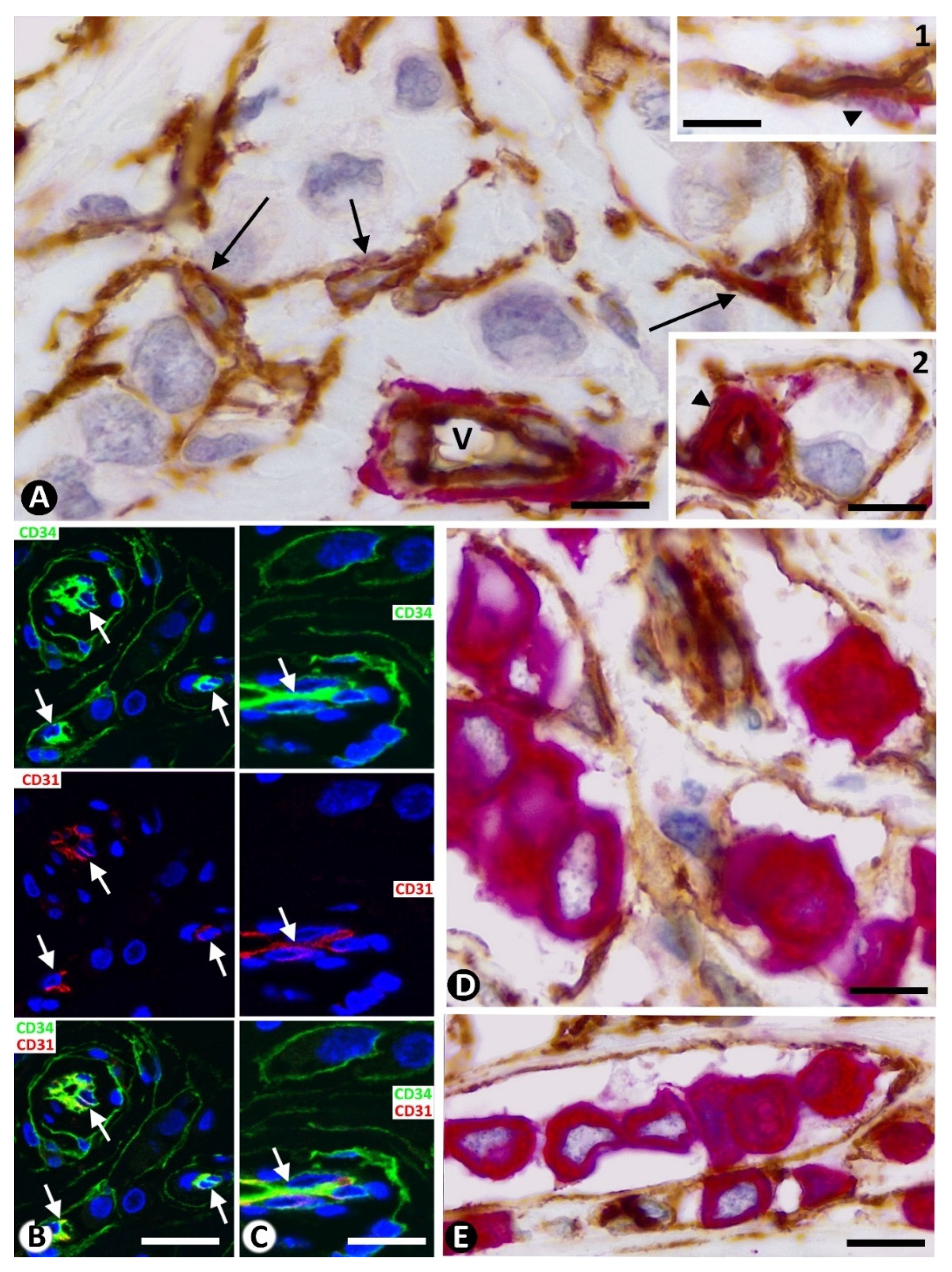
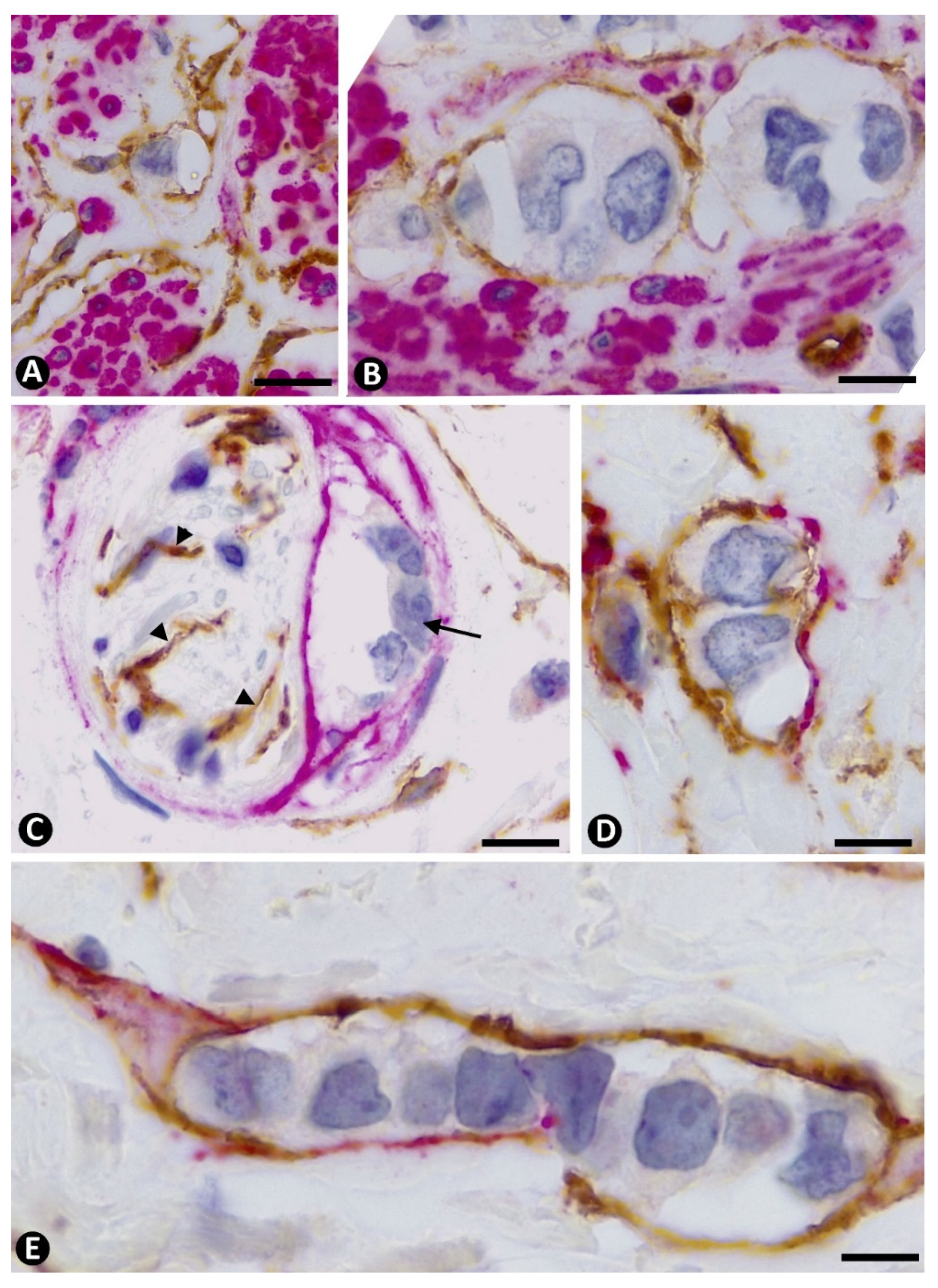
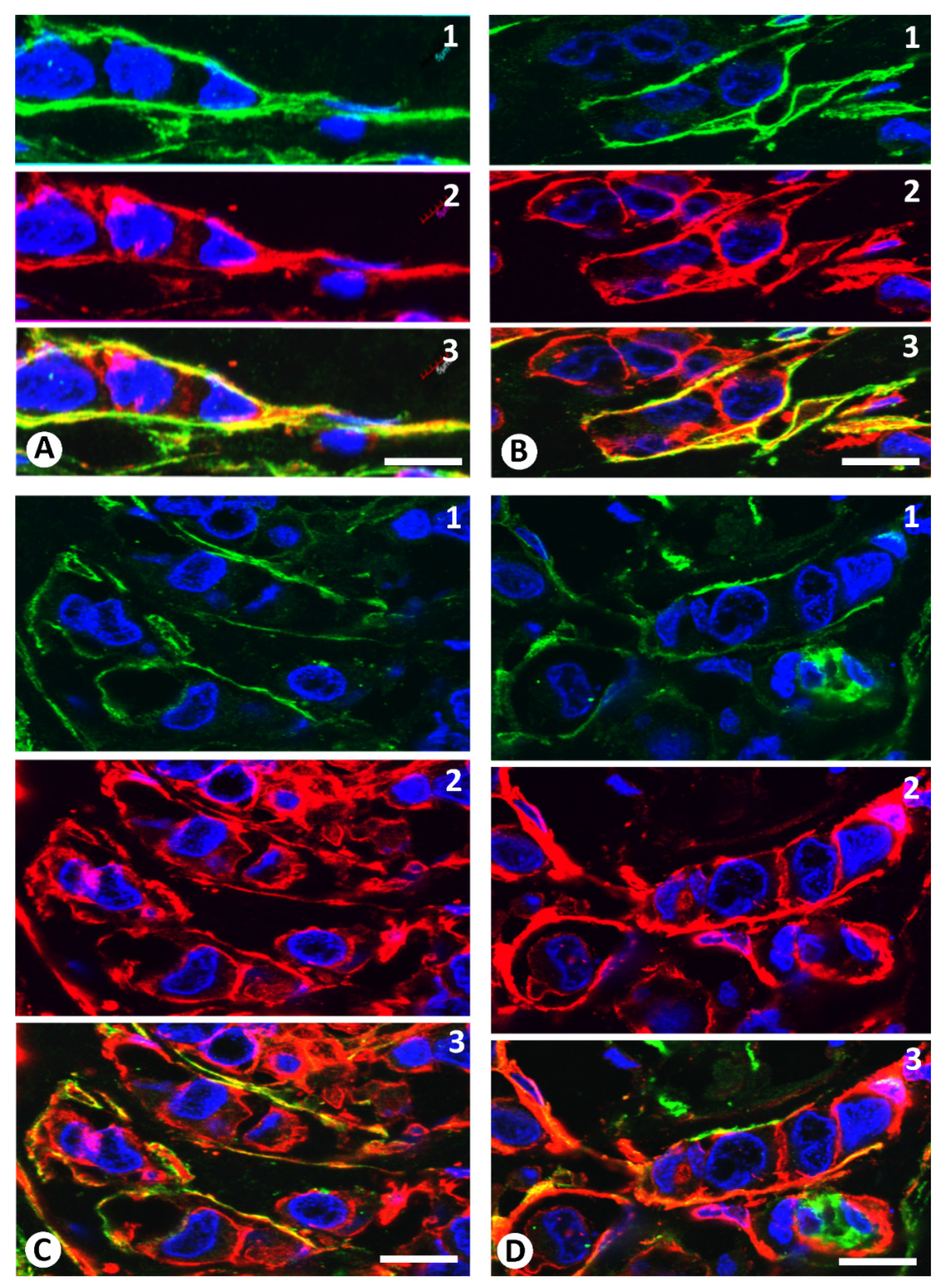
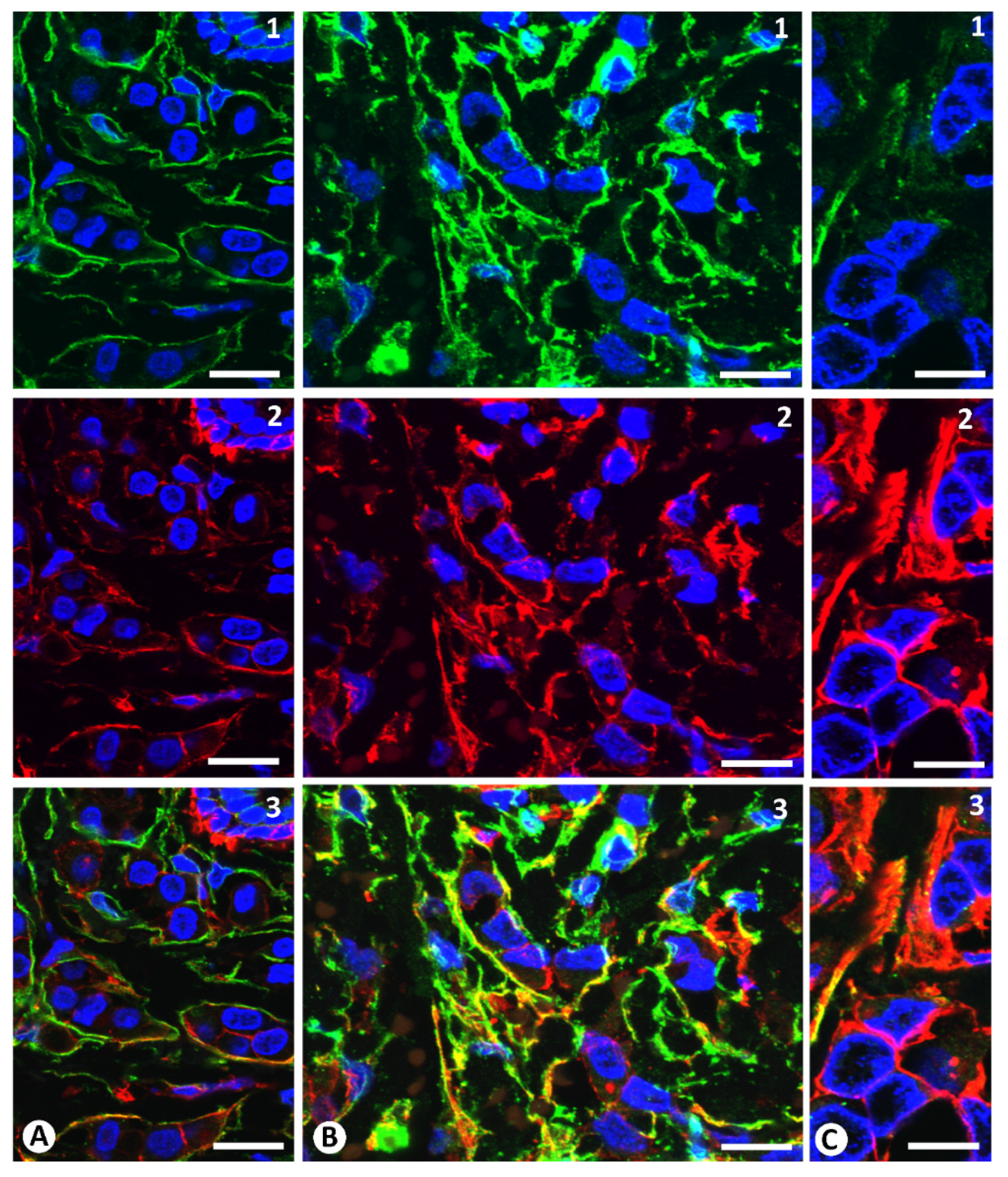
2.1. Stromal Cells around Strands and Nests of Neoplastic Cells in Invasive Lobular Carcinoma
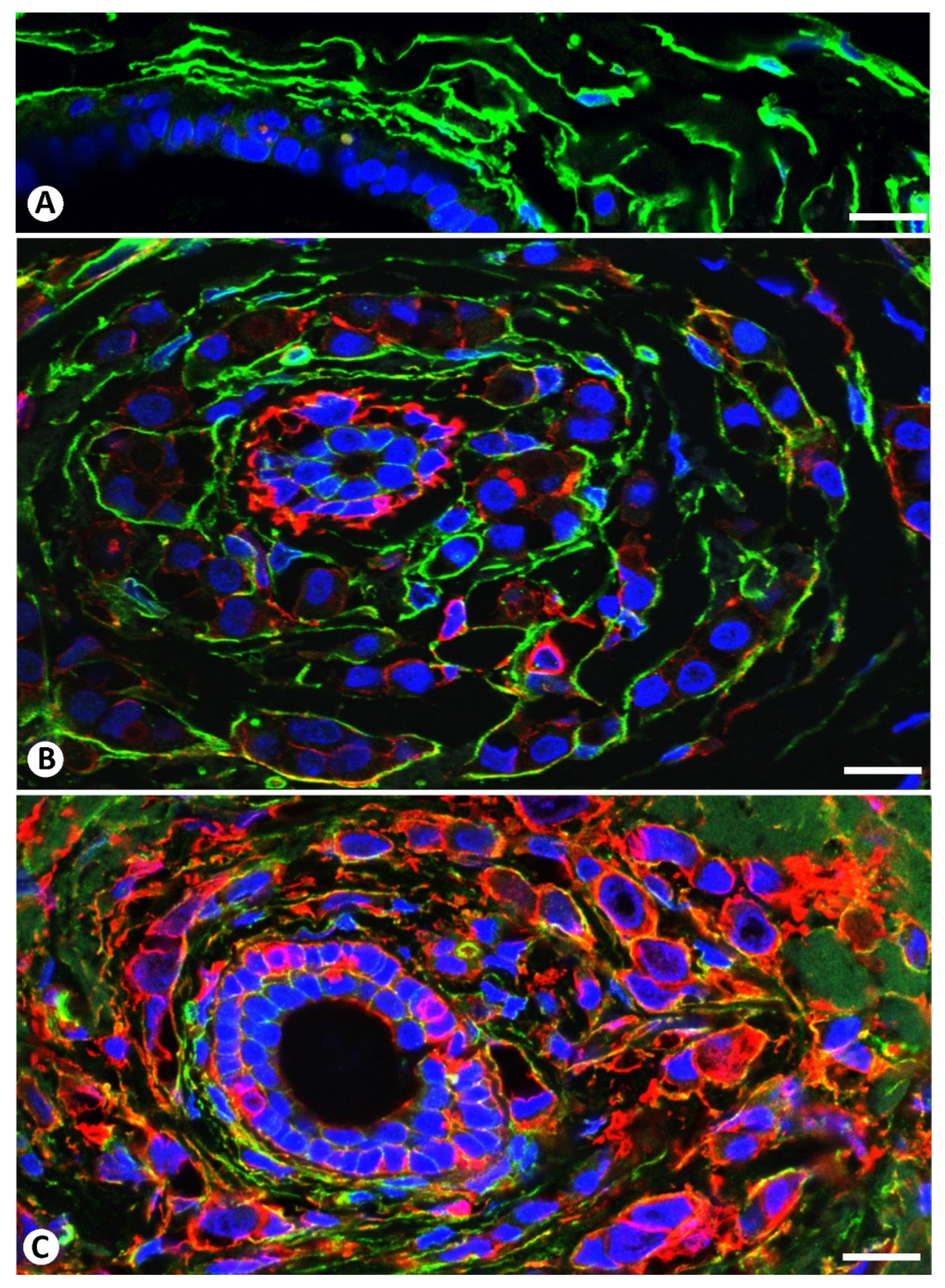
2.2. The Patterns of Arrangement and Location of Stromal Cells (CD34+ and αSMA+ Stromal Cells) in Invasive Lobular Carcinoma Are Similar to Those of CD34+SCs/TCs in the Normal Breast
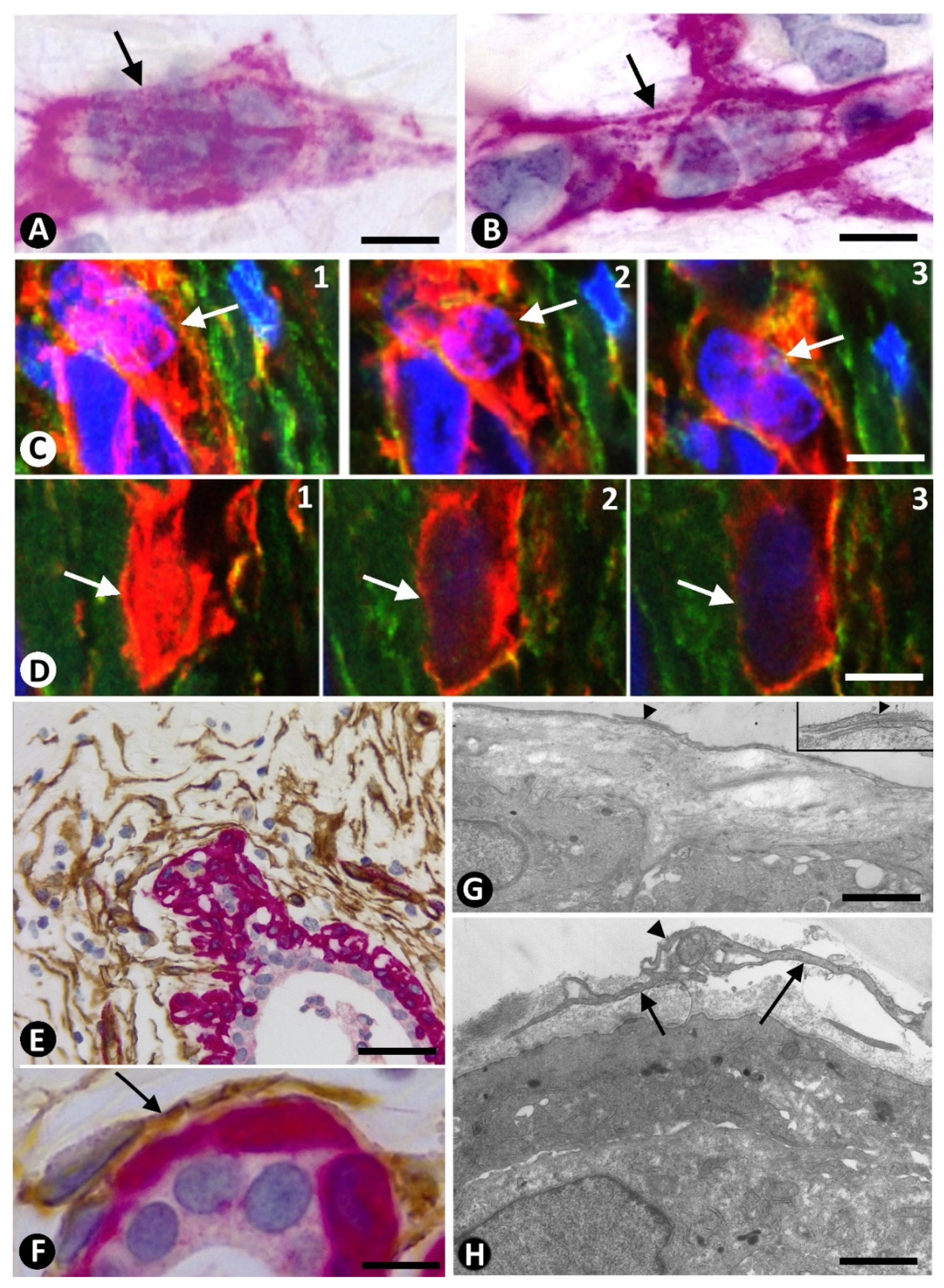
2.3. Presence of Both Types of Stromal Cells (CD34+ and αSMA+ Stromal Cells) around the Same Strand and Nest of Neoplastic Cells
2.4. Coexpression of CD34 and αSMA in Stromal Cells of Invasive Lobular Carcinoma
3. Discussion
4. Material and Methods
4.1. Tissue Samples
4.2. Light Microscopy
4.3. Immunohistochemistry
4.4. Immunofluorescence in Confocal Microscopy
4.5. Electron Microscopy
4.6. Semiquantitative Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faussone-Pellegrini, M.S.; Popescu, L.M. Telocytes. BioMol. Concepts 2011, 2, 481–489. [Google Scholar] [CrossRef]
- Popescu, L.M.; Faussone-Pellegrini, M.S. Telocytes—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to telocytes. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; Sáez, F.J.; Díaz-Flores, L., Jr.; Valladares, F.; Madrid, J.F. CD34+ stromal cells/fibroblasts/fibrocytes/telocytes as a tissue reserve and a principal source of mesenchymal cells. Location, morphology, function and role in pathology. Histol. Histopathol. 2014, 29, 831–870. [Google Scholar] [PubMed]
- Manetti, M.; Tani, A.; Rosa, I.; Chellini, F.; Squecco, R.; Idrizaj, E.; Zecchi-Orlandini, S.; Ibba-Manneschi, L.; Sassoli, C. Morphological evidence for telocytes as stromal cells supporting satellite cell activation in eccentric contraction-induced skeletal muscle injury. Sci. Rep. 2019, 9, 14515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusu, M.C.; Cretoiu, D.; Vrapciu, A.D.; Hostiuc, S.; Dermengiu, D.; Manoiu, V.S.; Cretoiu, S.M.; Mirancea, N. Telocytes of the human adult trigeminal ganglion. Cell Biol. Toxicol. 2016, 32, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Hummel, E.; Zimmermann, H.; Gherghiceanu, M.; Popescu, L.M. Human cardiac telocytes: 3D imaging by FIB-SEM tomography. J. Cell. Mol. Med. 2014, 18, 2157–2164. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Gherghiceanu, M.; Hummel, E.; Zimmermann, H.; Simionescu, O.; Popescu, L.M. FIB-SEM tomography of human skin telocytes and their extracellular vesicles. J. Cell. Mol. Med. 2015, 19, 714–722. [Google Scholar] [CrossRef] [Green Version]
- Cretoiu, D. The Third Dimension of Telocytes Revealed by FIB-SEM Tomography. Adv. Exp. Med. Biol. 2016, 913, 325–334. [Google Scholar]
- Barth, P.J.; Ramaswamy, A.; Moll, R. CD34(+) fibrocytes in normal cervical stroma, cervical intraepithelial neoplasia III, and invasive squamous cell carcinoma of the cervix uteri. Virch. Arch. 2002, 441, 564–568. [Google Scholar] [CrossRef]
- Barth, P.J.; Ebrahimsade, S.; Ramaswamy, A.; Moll, R. CD34+ fibrocytes in invasive ductal carcinoma, ductal carcinoma in situ, and benign breast lesions. Virch. Arch. 2002, 440, 298–303. [Google Scholar] [CrossRef]
- Barth, P.J.; Ebrahimsade, S.; Hellinger, A.; Moll, R.; Ramaswamy, A. CD34+ fibrocytes in neoplastic and inflammatory pancreatic lesions. Virch. Arch. 2002, 440, 128–133. [Google Scholar] [CrossRef]
- Barth, P.J.; Schenck zu Schweinsberg, T.; Ramaswamy, A.; Moll, R. CD34+ fibrocytes, alpha-smooth muscle antigen-positive myofibroblasts, and CD117 expression in the stroma of invasive squamous cell carcinomas of the oral cavity, pharynx, and larynx. Virch. Arch. 2004, 444, 231–234. [Google Scholar] [CrossRef]
- Barth, P.J.; Moll, R.; Ramaswamy, A. Stromal remodeling and SPARC (secreted protein acid rich in cysteine) expression in invasive ductal carcinomas of the breast. Virch. Arch. 2005, 446, 532–536. [Google Scholar] [CrossRef]
- Chauhan, H.; Abraham, A.; Phillips, J.R.; Pringle, J.H.; Walker, R.A.; Jones, J.L. There is more than one kind of myofibroblast: Analysis of CD34 expression in benign, in situ, and invasive breast lesions. J. Clin. Pathol. 2003, 56, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, N.; Nakayama, H.; Miyazaki, E.; Toi, M.; Hiroi, M.; Enzan, H. The distribution of CD34-positive stromal cells and myofibroblasts in colorectal carcinoid tumors. Histol. Histopathol. 2005, 20, 27–33. [Google Scholar]
- Kuroda, N.; Guo, L.; Miyazaki, E.; Hamauzu, T.; Toi, M.; Hiroi, M.; Enzan, H. The appearance of myofibroblasts and the disappearance of CD34-positive stromal cells in the area adjacent to xanthogranulomatous foci of chronic cholecystitis. Histol. Histopathol. 2005, 20, 127–133. [Google Scholar]
- Nakayama, H.; Enzan, H.; Miyazaki, E.; Kuroda, N.; Naruse, K.; Hiroi, M. Differential expression of CD34 in normal colorectal tissue, peritumoral inflammatory tissue, and tumour stroma. J. Clin. Pathol. 2000, 53, 626–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, H.; Enzan, H.; Miyazaki, E.; Kuroda, N.; Naruse, K.; Kiyoku, H.; Toi, M.; Hiroi, M. CD34 positive stromal cells in gastric adenocarcinomas. J. Clin. Pathol. 2001, 54, 846–848. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, H.; Enzan, H.; Yamamoto, M.; Miyazaki, E.; Hidaka, C.; Okumichi, T.; Okumichi, A.; Kajihara, H. CD34-positive stromal cells in primary lung carcinomas. Oncol. Rep. 2003, 10, 1313–1316. [Google Scholar] [CrossRef] [PubMed]
- Nimphius, W.; Moll, R.; Olbert, P.; Ramaswamy, A.; Barth, P.J. CD34+ fibrocytes in chronic cystitis and noninvasive and invasive urothelial carcinomas of the urinary bladder. Virch. Arch. 2007, 450, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, A.; Moll, R.; Barth, P.J. CD34+ fibrocytes in tubular carcinomas and radial scars of the breast. Virch. Arch. 2003, 443, 536–540. [Google Scholar] [CrossRef]
- Wessel, C.; Westhoff, C.C.; Nowak, K.; Moll, I.; Barth, P.J. CD34(+) fibrocytes in melanocytic nevi and malignant melanomas of the skin. Virch. Arch. 2008, 453, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Anderberg, C.; Pietras, K. On the origin of cancer-associated fibroblasts. Cell Cycle 2009, 8, 1461–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartoschek, M.; Oskolkov, N.; Bocci, M.; Lövrot, J.; Larsson, C.; Sommarin, M.; Madsen, C.D.; Lindgren, D.; Pekar, G.; Karlsson, G.; et al. Spatially and functionally distinct subclasses of breast cancer-associated fibroblasts revealed by single cell RNA sequencing. Nat. Commun. 2018, 9, 5150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochet, L.; Lehuédé, C.; Dauvillier, S.; Wang, Y.Y.; Dirat, B.; Laurent, V.; Dray, C.; Guiet, R.; Maridonneau-Parini, I.; Le Gonidec, S.; et al. Adipocyte-derived fibroblasts promote tumor progression and contribute to the desmoplastic reaction in breast cancer. Cancer Res. 2013, 73, 5657–5668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzobo, K.; Dandara, C. Architecture of Cancer-Associated Fibroblasts in Tumor Microenvironment: Mapping Their Origins, Heterogeneity, and Role in Cancer Therapy Resistance. OMICS 2020, 24, 314–339. [Google Scholar] [CrossRef] [PubMed]
- Kurashige, M.; Kohara, M.; Ohshima, K.; Tahara, S.; Hori, Y.; Nojima, S.; Wada, N.; Ikeda, J.I.; Miyamura, K.; Ito, M.; et al. Origin of cancer-associated fibroblasts and tumor-associated macrophages in humans after sex-mismatched bone marrow transplantation. Commun. Biol. 2018, 1, 131. [Google Scholar] [CrossRef] [Green Version]
- Ostman, A.; Augsten, M. Cancer-associated fibroblasts and tumor growth--bystanders turning into key players. Curr. Opin. Genet. Dev. 2009, 19, 67–73. [Google Scholar] [CrossRef]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Barth, P.J.; Westhoff, C.C. CD34+ fibrocytes: Morphology, histogenesis and function. Curr. Stem Cell Res. Ther. 2007, 2, 221–227. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; González, M.; Sáez, F.J.; Aparicio, F.; Díaz-Flores, L., Jr.; Madrid, J.F. Human resident CD34+ stromal cells/telocytes have progenitor capacity and are a source of αSMA+ cells during repair. Histol. Histopathol. 2015, 30, 615–627. [Google Scholar] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; Lizartza, K.; Goméz, M.G.; García, M.P.; Sáez, F.J.; Díaz-Flores, L., Jr.; Madrid, J.F. Behavior of in situ human native adipose tissue CD34+ stromal/progenitor cells during different stages of repair. Tissue-resident CD34+ stromal cells as a source of myofibroblasts. Anat. Rec. 2015, 298, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; González-Gómez, M.; Carrasco, J.L.; Alvarez-Argüelles, H.; Díaz-Flores, L., Jr. Telocytes/CD34+ Stromal Cells in Pathologically Affected White Adipose Tissue. Int. J. Mol. Sci. 2020, 21, 9694. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimsade, S.; Westhoff, C.C.; Barth, P.J. CD34+ fibrocytes are preserved in most invasive lobular carcinomas of the breast. Pathol. Res. Prac. 2007, 203, 695–698. [Google Scholar] [CrossRef] [PubMed]
- El-Tahawy, N.F.G.; Rifaai, R.A. Immunohistochemical and ultrastructural evidence for telocytes in the different physiological stages of the female rat mammary gland. Life Sci. 2019, 231, 116521. [Google Scholar] [CrossRef] [PubMed]
- Gherghiceanu, M.; Popescu, L.M. Interstitial Cajal-like cells (ICLC) in human resting mammary gland stroma. Transmission electron microscope (TEM) identification. J. Cell. Mol. Med. 2005, 9, 893–910. [Google Scholar] [CrossRef] [PubMed]
- Hostiuc, S.; Negoi, I.; Perlea, P. Morphology of telocytes from the mammary gland. A review. Gineco.eu 2018, 14, 36–38. [Google Scholar]
- Janas, P.; Kucybała, I.; Radoń-Pokracka, M.; Huras, H. Telocytes in the female reproductive system: An overview of up-to-date knowledge. Adv. Clin. Exp. Med. 2018, 27, 559–565. [Google Scholar] [CrossRef]
- Mou, Y.; Wang, Y.; Li, J.; Lü, S.; Duan, C.; Du, Z.; Yang, G.; Chen, W.; Zhao, S.; Zhou, J.; et al. Immunohistochemical characterization and functional identification of mammary gland telocytes in the self-assembly of reconstituted breast cancer tissue in vitro. J. Cell. Mol. Med. 2013, 17, 65–75. [Google Scholar] [CrossRef]
- Petre, N.; Rusu, M.C.; Pop, F.; Jianu, A.M. Telocytes of the mammary gland stroma. Folia Morphol. 2016, 75, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Radu, E.; Regalia, T.; Ceafalan, L.; Andrei, F.; Cretoiu, D.; Popescu, L.M. Cajal-type cells from human mammary gland stroma: Phenotype characteristics in cell culture. J. Cell. Mol. Med. 2005, 9, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Westhoff, C.C.; Jank, P.; Jacke, C.O.; Albert, U.S.; Ebrahimsade, S.; Barth, P.J.; Moll, R. Prognostic relevance of the loss of stromal CD34 positive fibroblasts in invasive lobular carcinoma of the breast. Virch. Arch. 2020, 477, 717–724. [Google Scholar] [CrossRef]
- Marini, M.; Rosa, I.; Ibba-Manneschi, L.; Manetti, M. Telocytes in skeletal, cardiac and smooth muscle interstitium: Morphological and functional aspects. Histol. Histopathol. 2018, 33, 1151–1165. [Google Scholar] [PubMed]
- Albulescu, R.; Tanase, C.; Codrici, E.; Popescu, D.I.; Cretoiu, S.M.; Popescu, L.M. The secretome of myocardial telocytes modulates the activity of cardiac stem cells. J. Cell. Mol. Med. 2015, 19, 1783–1794. [Google Scholar] [CrossRef] [PubMed]
- Bani, D.; Formigli, L.; Gherghiceanu, M.; Faussone-Pellegrini, M.S. Telocytes as supporting cells for myocardial tissue organization in developing and adult heart. J. Cell. Mol. Med. 2010, 14, 2531–2538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cretoiu, S.M.; Popescu, L.M. Telocytes revisited. BioMol. Concepts 2014, 5, 353–369. [Google Scholar] [CrossRef]
- Popescu, L.M. The tandem: Telocytes–stem cells. Int. J. Biol. Biomed. Eng. 2011, 5, 83–92. [Google Scholar]
- Rosa, I.; Taverna, C.; Novelli, L.; Marini, M.; Ibba-Manneschi, L.; Manetti, M. Telocytes constitute a widespread interstitial meshwork in the lamina propria and underlying striated muscle of human tongue. Sci. Rep. 2019, 9, 5858. [Google Scholar] [CrossRef] [Green Version]
- Rosa, I.; Faussone-Pellegrini, M.S.; Romano, E.; Ibba-Manneschi, L.; Matucci-Cerinic, M.; Manetti, M. Impairment in the telocyte/CD34+ stromal cell network in human rheumatoid arthritis synovium. J. Cell. Mol. Med. 2021, 25, 2274–2278. [Google Scholar] [CrossRef] [PubMed]
- San Martin, R.; Barron, D.A.; Tuxhorn, J.A.; Ressler, S.J.; Hayward, S.W.; Shen, X.; Laucirica, R.; Wheeler, T.M.; Gutierrez, C.; Ayala, G.E.; et al. Recruitment of CD34(+) fibroblasts in tumor-associated reactive stroma: The reactive microvasculature hypothesis. Am. J. Pathol. 2014, 184, 1860–1870. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Han, C.; Wang, S.; Fang, P.; Ma, Z.; Xu, L.; Yin, R. Cancer-associated fibroblasts: An emerging target of anti-cancer immunotherapy. J. Hematol. Oncol. 2019, 12, 86. [Google Scholar] [CrossRef]
- Costa, A.; Kieffer, Y.; Scholer-Dahirel, A.; Pelon, F.; Bourachot, B.; Cardon, M.; Sirven, P.; Magagna, I.; Fuhrmann, L.; Bernard, C.; et al. Fibroblast Heterogeneity and Immunosuppressive Environment in Human Breast Cancer. Cancer Cell 2018, 33, 463–479.e10. [Google Scholar] [CrossRef] [Green Version]
- Higgins, S.P.; Tang, Y.; Higgins, C.E.; Mian, B.; Zhang, W.; Czekay, R.P.; Samarakoon, R.; Conti, D.J.; Higgins, P.J. TGF-β1/p53 signaling in renal fibrogenesis. Cell Signal. 2018, 43, 1–10. [Google Scholar] [CrossRef]
- McKeown-Longo, P.J.; Higgins, P.J. Hyaluronan, Transforming Growth Factor β, and Extra Domain A-Fibronectin: A Fibrotic Triad. Adv. Wound Care 2021, 10, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Gifford, C.C.; Samarakoon, R.; Higgins, P.J. Deregulation of Negative Controls on TGF-β1 Signaling in Tumor Progression. Cancers 2018, 10, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, X.; Young, C.D.; Zhou, H.; Wang, X. Transforming Growth Factor-β Signaling in Fibrotic Diseases and Cancer-Associated Fibroblasts. Biomolecules 2020, 10, 1666. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Li, Y.; Zhu, S.; Yu, J.; Zhang, B.; Chen, X.; Zhang, Z.; Ma, Y.; Niu, Y.; Shang, Z. YAP1 plays a key role of the conversion of normal fibroblasts into cancer-associated fibroblasts that contribute to prostate cancer progression. J. Exp. Clin. Cancer Res. 2020, 39, 36. [Google Scholar] [CrossRef]
- Schulze, A.B.; Schmidt, L.H.; Heitkötter, B.; Huss, S.; Mohr, M.; Marra, A.; Hillejan, L.; Görlich, D.; Barth, P.J.; Rehkämper, J.; et al. Prognostic impact of CD34 and SMA in cancer-associated fibroblasts in stage I-III NSCLC. Thorac. Cancer 2020, 11, 120–129. [Google Scholar] [CrossRef]
- Cangkrama, M.; Wietecha, M.; Mathis, N.; Okumura, R.; Ferrarese, L.; Al-Nuaimi, D.; Antsiferova, M.; Dummer, R.; Innocenti, M.; Werner, S. A paracrine activin A-mDia2 axis promotes squamous carcinogenesis via fibroblast reprogramming. EMBO Mol. Med. 2020, 12, e11466. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Breast | Invasive Lobular Carcinoma of the Breast | |||||
|---|---|---|---|---|---|---|
| N° and Percentage of Cases | Percentage of CD34+SCs/TCs | Percentage of αSMA+ Stromal Cells | N° and Percentage of Cases | Percentage of CD34+SCs/TCs | Percentage of αSMA+ Stromal Cells | |
| Predominance of CD34+SCs/TCs | 6 (100%) | 100% | 0% | 11 (26.19%) | >70% | <30% |
| Balanced Proportion of CD34+ and αSMA Stromal Cells | 0% | 0% | 0% | 18 (42.85%) | 30–70% | 30–70% |
| Predominance of αSMA+ Stromal Cells | 0% | 0% | 0% | 13 (30.95%) | <30% | >70% |
| Normal Breast | Invasive Lobular Carcinoma | ||||
|---|---|---|---|---|---|
| CD34+ Stromal Cells | αSMA+ Stromal Cells | Predominance of CD34+ Stromal Cells | Balanced Proportion of CD34+ and αSMA+ Stromal Cells | Predominance of αSMA+ Stromal Cells | |
| N° and % | 6 (100%) | 0 (0%) | 11 (26.19%) | 18 (42.85%) | 13 (30.95%) |
| Correspondence in Location and Arrangement of CD34+ and αSMA+ Stromal Cells | Periductal, Perivascular, Interlobular interstitium, septal and intralobular adipose tissue smooth muscle of the nipples | Coincidence with normal breast | Coincidence with normal breast | Coincidence with normal breast | |
| Coincidence of Both Types of Stromal Cells in Neoplastic Nests/Strands | 0 | 0 | 26.09 ± 4.39 | 32,27 ± 8.44 | 22.07 ± 5.23 |
| Coexpression of CD34+ and αSMA+ in Stromal Cells (mean ± s.d.) | 0 | 0 | 27.63 ± 3.11 | 29.77 ± 6.50 | 25.25 ± 4.28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Flores, L.; Gutiérrez, R.; González-Gómez, M.; García, M.P.; Díaz-Flores, L., Jr.; Carrasco, J.L.; Martín-Vasallo, P. CD34+ Stromal Cells/Telocytes as a Source of Cancer-Associated Fibroblasts (CAFs) in Invasive Lobular Carcinoma of the Breast. Int. J. Mol. Sci. 2021, 22, 3686. https://doi.org/10.3390/ijms22073686
Díaz-Flores L, Gutiérrez R, González-Gómez M, García MP, Díaz-Flores L Jr., Carrasco JL, Martín-Vasallo P. CD34+ Stromal Cells/Telocytes as a Source of Cancer-Associated Fibroblasts (CAFs) in Invasive Lobular Carcinoma of the Breast. International Journal of Molecular Sciences. 2021; 22(7):3686. https://doi.org/10.3390/ijms22073686
Chicago/Turabian StyleDíaz-Flores, Lucio, Ricardo Gutiérrez, Miriam González-Gómez, Maria Pino García, Lucio Díaz-Flores, Jr., José Luís Carrasco, and Pablo Martín-Vasallo. 2021. "CD34+ Stromal Cells/Telocytes as a Source of Cancer-Associated Fibroblasts (CAFs) in Invasive Lobular Carcinoma of the Breast" International Journal of Molecular Sciences 22, no. 7: 3686. https://doi.org/10.3390/ijms22073686
APA StyleDíaz-Flores, L., Gutiérrez, R., González-Gómez, M., García, M. P., Díaz-Flores, L., Jr., Carrasco, J. L., & Martín-Vasallo, P. (2021). CD34+ Stromal Cells/Telocytes as a Source of Cancer-Associated Fibroblasts (CAFs) in Invasive Lobular Carcinoma of the Breast. International Journal of Molecular Sciences, 22(7), 3686. https://doi.org/10.3390/ijms22073686