
Proteomic Comparison of Bone Marrow Derived Osteoblasts and Mesenchymal Stem Cells

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Global Proteomic Profiles of Osteoblasts and MSCs
2.2. Qualitative Proteomic Differences between Osteoblasts and MSCs: A Minor Protein Subset Is Only Detected in Osteoblasts and Reflects Their Differentiation and/or Specialized Functions
2.3. A Minor Subset of Diverse Proteins Reach Quantifiable Levels Only for MSCs
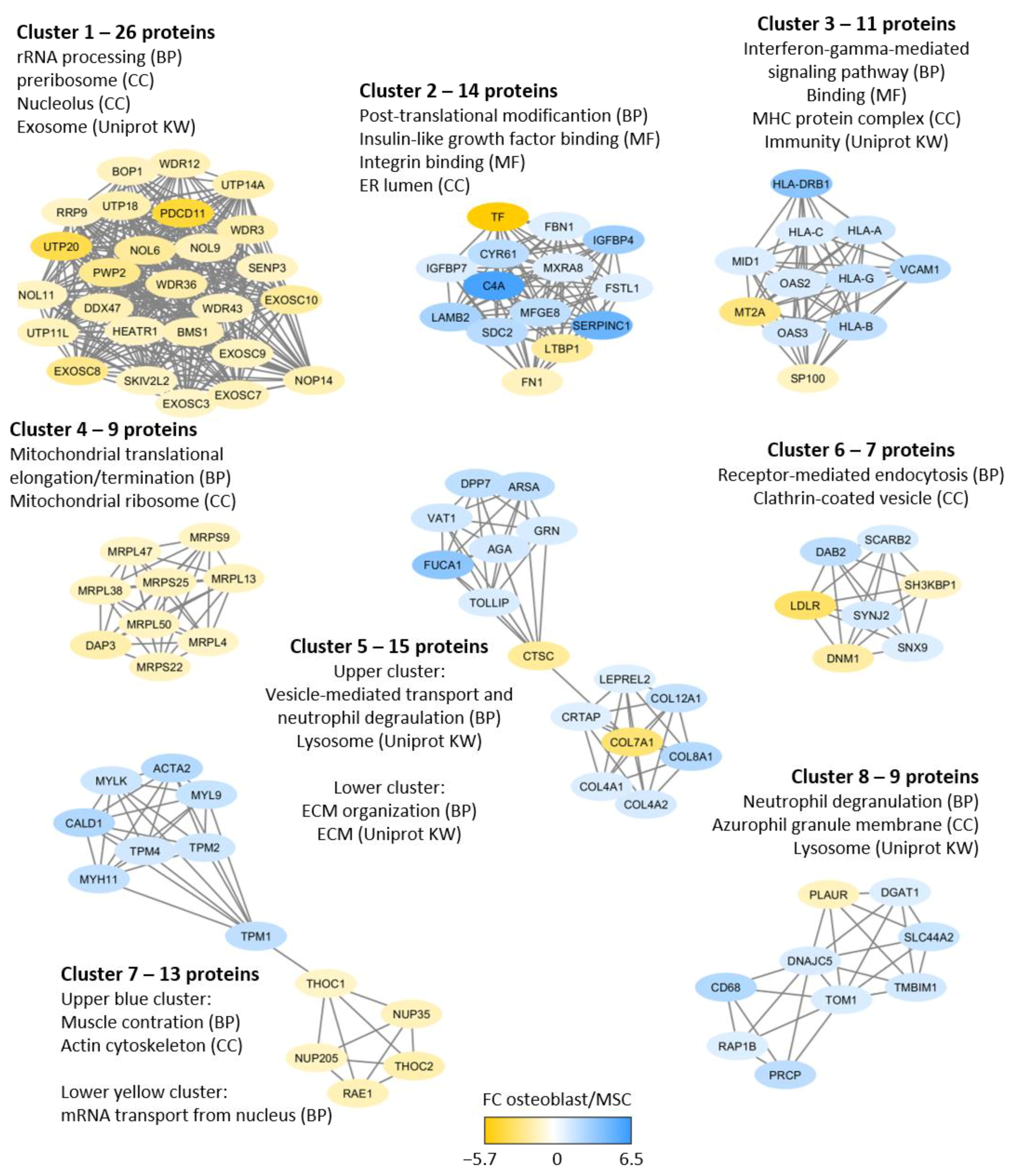
2.4. Quantitative Proteomic Differences between Osteoblasts and MSCs: Proteins Showing Increased Levels in Osteoblasts Reflect Cellular Differentiation and/or Specialized Functions whereas Proteins Increased in MSCs Reflect Differences in Transcriptional Regulation
- Formation, modulation, or binding to extracellular matrix molecules: CYR61, ENPPI, FBN1, FHL2, GPNMB, ITGA11, and LOX.
- Bone mineralization: ENPP1, EPHA2, FBN1, FHL2, GPNMB, LOX, and TPM4.
- Interactions with integrins: CYR61, FHL2, and ITGA11.
- Intracellular signaling important for osteoblastic differentiation and/or functions: PTH signaling (CYR61, TMEM119), Wnt/β-catenin signaling (CYR61, FHL2, GTPBP4, ITGA11, TMEM119), BMP2 (CYR61, FBN1, TMEM119), and RhoA signaling (EPHA2, GTPBP4).
- Transcription: Only a minority of the proteins are regarded as transcriptional regulators (decreased levels of CEBPB, DDX21, EPHA2; increased levels of LOX), are important for cellular metabolism (FASN2) or are involved in exosomal communication between cells (EPHA2).
2.5. Only a Small Number of Osteoblast Proteins Show Significant Differences when Comparing Male Vs. Female Osteoblast Donors and Elderly Vs. Younger Donors
2.6. Osteoblast Responses to Extended Culture Including Suboptimal In Vitro Culture Conditions
2.7. MSC Responses to Suboptimal In Vitro Culture Conditions
3. Discussion
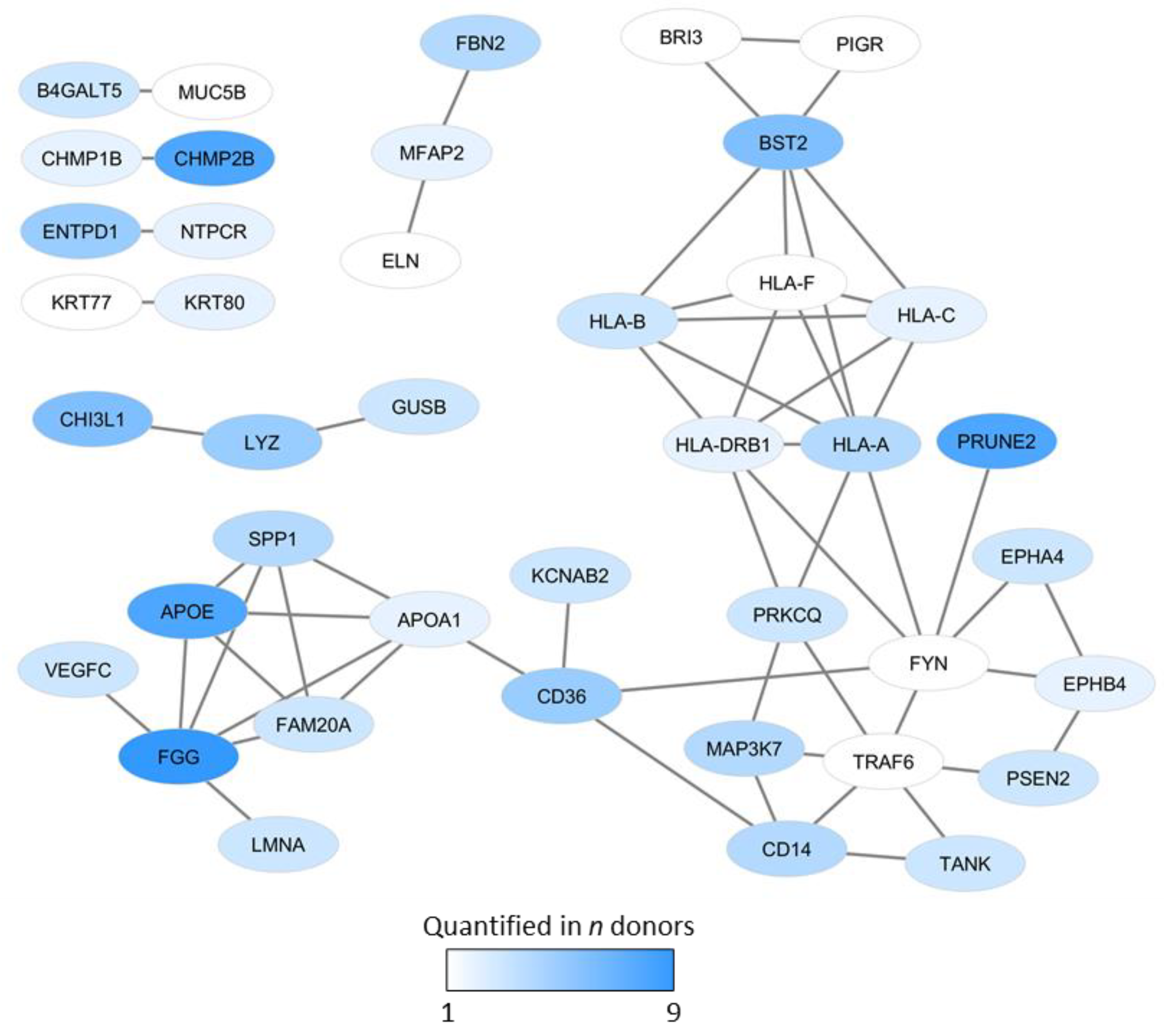
- Osteoblast differentiation: BST2, CHI3L1, FBN2, FYN, PRKCQ, PRUNE2, PSEN2, VEGFC.
- Transcriptional regulation: Regulators or targets of Runx2 (EPHB4, SPP1) and NFκB, (PRUNE2, PSEN2). LMAA is also regarded as a transcriptional regulator.
- Intracellular signaling pathways: the network included regulators of Wnt (BRI3), BMP (BST2, FBN2, CHMP1B, MAP3K7), RhoA (EPHB4, PRUNE2), Notch (PRUNE2, PSEN2), TGF (FBN2, ELN, MAP3K7) and TNF signaling (MAP3K7, TANK, TRAF6).
- Intracellular trafficking/transport. Osteoblasts are characterized by an extensive Golgi apparatus and endoplasmatic reticulum [27], and the interaction network included proteins that are important for the Golgi system (B4GALT5, FAM20A), intracellular trafficking (CHMP2B), and cytoskeleton (KRT77, KRT80).
- Bone metabolism/mineralization: B4GALT5, ELN, FBN2, TLR4, EPHB4, FGG.
- Bone remodeling/homeostasis: EPHB4, LMNA, TRAF6, TANK, SPP1, VEGFC.
4. Materials and Methods
4.1. Human MSCs and Osteoblasts
4.2. Preparation of Cell Samples: Original Cells and In Vitro Incubated Cell Samples
4.3. Proteomics Sample Preparation
4.4. Liquid Chromatography (LC) Tandem Mass Spectrometry (MS) Analysis
4.5. Statistical and Bioinformatical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Domingues, M.J.; Cao, H.; Heazlewood, S.Y.; Cao, B.; Nilsson, S.K. Niche Extracellular Matrix Components and Their Influence on HSC. J. Cell. Biochem. 2017, 118, 1984–1993. [Google Scholar] [CrossRef] [PubMed]
- Sangaletti, S.; Chiodoni, C.; Tripodo, C.; Colombo, M.P. Common extracellular matrix regulation of myeloid cell activity in the bone marrow and tumor microenvironments. Cancer Immunol. Immunother. 2017, 66, 1059–1067. [Google Scholar] [CrossRef] [Green Version]
- Klamer, S.; Voermans, C. The role of novel and known extracellular matrix and adhesion molecules in the homeostatic and regenerative bone marrow microenvironment. Cell Adh. Migr. 2014, 8, 563–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedi, A.; Santoro, A.; Dunant, C.F.; Dick, J.E.; Laurenti, E. Molecular landscapes of human hematopoietic stem cells in health and leukemia. Ann. N. Y. Acad. Sci. 2016, 1370, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Yu, V.W.; Scadden, D.T. Hematopoietic Stem Cell and Its Bone Marrow Niche. Curr. Top. Dev. Biol. 2016, 118, 21–44. [Google Scholar]
- Ehninger, A.; Trumpp, A. The bone marrow stem cell niche grows up: Mesenchymal stem cells and macrophages move in. J. Exp. Med. 2011, 208, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Kunisaki, Y.; Bruns, I.; Scheiermann, C.; Ahmed, J.; Pinho, S.; Zhang, D.; Mizoguchi, T.; Wei, Q.; Lucas, D.; Ito, K.; et al. Arteriolar niches maintain haematopoietic stem cell quiescence. Nature 2013, 502, 637–643. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhang, W.; Ran, Q.; Xiang, Y.; Zhong, J.F.; Li, S.C.; Li, Z. The Differentiation Balance of Bone Marrow Mesenchymal Stem Cells Is Crucial to Hematopoiesis. STEM Cells Int. 2018, 2018, 1540148. [Google Scholar] [CrossRef]
- Garg, P.; Mazur, M.M.; Buck, A.C.; Wandtke, M.E.; Liu, J.; Ebraheim, N.A. Prospective Review of Mesenchymal Stem Cells Differentiation into Osteoblasts. Orthop. Surg. 2017, 9, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Shou, P.; Zheng, C.; Jiang, M.; Cao, G.; Yang, Q.; Cao, J.; Xie, N.; Velletri, T.; Zhang, X.; et al. Fate decision of mesenchymal stem cells: Adipocytes or osteoblasts? Cell Death Differ. 2016, 23, 1128–1139. [Google Scholar] [CrossRef] [Green Version]
- Brenner, A.K.; Andersson Tvedt, T.H.; Bruserud, Ø. The Complexity of Targeting PI3K-Akt-mTOR Signalling in Human Acute Myeloid Leukaemia: The Importance of Leukemic Cell Heterogeneity, Neighbouring Mesenchymal Stem Cells and Immunocompetent Cells. Molecules 2016, 21, 1512. [Google Scholar] [CrossRef]
- Bergfeld, S.A.; DeClerck, Y.A. Bone marrow-derived mesenchymal stem cells and the tumor microenvironment. Cancer Metastasis Rev. 2010, 29, 249–261. [Google Scholar] [CrossRef]
- Shupp, A.B.; Kolb, A.D.; Mukhopadhyay, D.; Bussard, K.M. Cancer Metastases to Bone: Concepts, Mechanisms, and Interactions with Bone Osteoblasts. Cancers 2018, 10, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clézardin, P. Pathophysiology of bone metastases from solid malignancies. Joint Bone Spine 2017, 84, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Esposito, M.; Kang, Y. Bone metastasis and the metastatic niche. J. Mol. Med. 2015, 93, 1203–1212. [Google Scholar] [CrossRef] [Green Version]
- Bruserud, Ø.; Ryningen, A.; Wergeland, L.; Glenjen, N.I.; Gjertsen, B.T. Osteoblasts increase proliferation and release of pro-angiogenic interleukin 8 by native human acute myelogenous leukemia blasts. Haematologica 2004, 89, 391–402. [Google Scholar] [PubMed]
- Brenner, A.K.; Nepstad, I.; Bruserud, Ø. Mesenchymal Stem Cells Support Survival and Proliferation of Primary Human Acute Myeloid Leukemia Cells through Heterogeneous Molecular Mechanisms. Front. Immunol. 2017, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- Kornblau, S.M.; Ruvolo, P.P.; Wang, R.Y.; Battula, V.L.; Shpall, E.J.; Ruvolo, V.R.; McQueen, T.; Qui, Y.; Zeng, Z.; Pierce, S.; et al. Distinct protein signatures of acute myeloid leukemia bone marrow-derived stromal cells are prognostic for patient survival. Haematologica 2018, 103, 810–821. [Google Scholar] [CrossRef]
- Zheng, H.; Bae, Y.; Kasimir-Bauer, S.; Tang, R.; Chen, J.; Ren, G.; Yuan, M.; Esposito, M.; Li, W.; Wei, Y.; et al. Therapeutic Antibody Targeting Tumor- and Osteoblastic Niche-Derived Jagged1 Sensitizes Bone Metastasis to Chemotherapy. Cancer Cell 2017, 32, 731–747. [Google Scholar] [CrossRef] [Green Version]
- Brommage, R. New Targets and Emergent Therapies for Osteoporosis. Handb. Exp. Pharmacol. 2020, 262, 451–473. [Google Scholar]
- Wu, M.; Chen, G.; Li, Y.P. TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef] [PubMed]
- Aslani, S.; Abhari, A.; Sakhinia, E.; Sanajou, D.; Rajabi, H.; Rahimzadeh, S. Interplay between microRNAs and Wnt, transforming growth factor-β, and bone morphogenic protein signaling pathways promote osteoblastic differentiation of mesenchymal stem cells. J. Cell. Physiol. 2019, 234, 8082–8093. [Google Scholar] [CrossRef] [PubMed]
- Eppert, K.; Takenaka, K.; Lechman, E.R.; Waldron, L.; Nilsson, B.; van Galen, P.; Metzeler, K.H.; Poeppl, A.; Ling, V.; Beyene, J.; et al. Stem cell gene expression programs influence clinical outcome in human leukemia. Nat. Med. 2011, 17, 1086–1093. [Google Scholar] [CrossRef]
- Kaushansky, K.; Zhan, H. The marrow stem cell niche in normal and malignant hematopoiesis. Ann. N. Y. Acad. Sci. 2020, 1466, 17–23. [Google Scholar] [CrossRef]
- Kumar, B.; Garcia, M.; Weng, L.; Jung, X.; Murakami, J.L.; Hu, X.; McDonald, T.; Lin, A.; Kumar, A.R.; DiGiusto, D.L.; et al. Acute myeloid leukemia transforms the bone marrow niche into a leukemia-permissive microenvironment through exosome secretion. Leukemia 2018, 32, 575–587. [Google Scholar] [CrossRef]
- Kolb, A.D.; Bussard, K.M. The Bone Extracellular Matrix as an Ideal Milieu for Cancer Cell Metastases. Cancers 2019, 11, 1020. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Malaval, L.; Aubin, J.E. The mature osteoblast phenotype is characterized by extensive plasticity. Exp. Cell Res. 1997, 232, 97–105. [Google Scholar] [CrossRef]
- Kenkre, J.S.; Bassett, J. The bone remodelling cycle. Ann. Clin. Biochem. 2018, 55, 308–327. [Google Scholar] [CrossRef]
- Noh, J.Y.; Yang, Y.; Jung, H. Molecular Mechanisms and Emerging Therapeutics for Osteoporosis. Int. J. Mol. Sci. 2020, 21, 7623. [Google Scholar] [CrossRef]
- Arthur, A.; Gronthos, S. Eph-Ephrin Signaling Mediates Cross-Talk Within the Bone Microenvironment. Front. Cell Dev. Biol. 2021, 9, 598612. [Google Scholar] [CrossRef]
- Ono, T.; Takayanagi, H. Osteoimmunology in Bone Fracture Healing. Curr. Osteoporos. Rep. 2017, 15, 367–375. [Google Scholar] [CrossRef]
- Shaw, A.T.; Gravallese, E.M. Mediators of inflammation and bone remodeling in rheumatic disease. Semin. Cell Dev. Biol. 2016, 49, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Brylka, L.J.; Schinke, T. Chemokines in Physiological and Pathological Bone Remodeling. Front. Immunol. 2019, 10, 2182. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Pérez, A.; Franco-Trepat, E.; Guillán-Fresco, M.; Jorge-Mora, A.; López, V.; Pino, J.; Gualillo, O.; Gómez, R. Role of Toll-Like Receptor 4 on Osteoblast Metabolism and Function. Front. Physiol. 2018, 9, 504. [Google Scholar] [CrossRef] [PubMed]
- Bruserud, Ø.; Aarstad, H.H.; Tvedt, T.H.A. Combined C-Reactive Protein and Novel Inflammatory Parameters as a Predictor in Cancer-What Can We Learn from the Hematological Experience? Cancers 2020, 12, 1966. [Google Scholar] [CrossRef]
- Liu, B.; Shao, Y.; Fu, R. Current research status of HLA in immune-related diseases. Immun. Inflamm. Dis. 2021. epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Aasebø, E.; Birkeland, E.; Selheim, F.; Berven, F.; Brenner, A.K.; Bruserud, Ø. The Extracellular Bone Marrow Microenvironment-A Proteomic Comparison of Constitutive Protein Release by In Vitro Cultured Osteoblasts and Mesenchymal Stem Cells. Cancers 2020, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.G.; Cerchio, K.; Stoch, S.A.; Gottesdiener, K.; Wu, M.; Recker, R.; L-000845704 Study Group. Effect of L-000845704, an alphaVbeta3 integrin antagonist, on markers of bone turnover and bone mineral density in postmenopausal osteoporotic women. J. Clin. Endocrinol. Metab. 2005, 90, 2022–2028. [Google Scholar]
- Pennypacker, B.; Shea, M.; Liu, Q.; Masarachia, P.; Saftig, P.; Rodan, S.; Rodan, G.; Kimmel, D. Bone density, strength, and formation in adult cathepsin K (−/−) mice. Bone 2009, 44, 199–207. [Google Scholar] [CrossRef]
- Brommage, R.; Liu, J.; Vogel, P.; Mseeh, F.; Thompson, A.Y.; Potter, D.G.; Shadoan, M.K.; Hansen, G.M.; Jeter-Jones, S.; Cui, J.; et al. NOTUM inhibition increases endocortical bone formation and bone strength. Bone Res. 2019, 7, 2. [Google Scholar]
- Brommage, R.; Liu, J.; Hansen, G.M.; Kirkpatrick, L.L.M.; Potter, D.G.; Sands, A.T.; Zambrowicz, B.; Powell, D.R.; Vogel, P. High-throughput screening of mouse gene knockouts identifies established and novel skeletal phenotypes. Bone Res. 2014, 2, 14034. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.; Boyette, L.B.; Tuan, R.S. Characterization of bone marrow-derived mesenchymal stem cells in aging. Bone 2015, 70, 37–47. [Google Scholar] [CrossRef]
- Aasebø., E.; Hernandez-Valladares, M.; Selheim, F.; Berven, F.S.; Brenner, A.K.; Bruserud, Ø. Proteomic Profiling of Primary Human Acute Myeloid Leukemia Cells Does Not Reflect Their Constitutive Release of Soluble Mediators. Proteomes 2018, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruserud, Ø.; Frostad, S.; Foss, B. In Vitro culture of acute myelogenous leukemia blasts: A comparison of four different culture media. J. Hematother. 1999, 8, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Frostad, S.; Bjerknes, R.; Abrahamsen, J.F.; Olweus, J.; Bruserud, Ø. Insulin-like growth factor-1 (IGF-1) has a costimulatory effect on proliferation of committed progenitors derived from human umbilical cord CD34+ cells. STEM Cells 1998, 16, 334–342. [Google Scholar] [CrossRef]
- Olweus, J.; Terstappen, L.W.; Thompson, P.A.; Lund-Johansen, F. Expression and function of receptors for stem cell factor and erythropoietin during lineage commitment of human hematopoietic progenitor cells. Blood 1996, 88, 1594–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruserud, Ø.; Tronstad, K.J.; Berge, R. In vitro culture of human osteosarcoma cell lines: A comparison of functional characteristics for cell lines cultured in medium without and with fetal calf serum. J. Cancer Res. Clin. Oncol. 2005, 131, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Gasson, S.B.; Dobson, L.K.; Chow, L.; Dow, S.; Gregory, C.A.; Saunders, W.B. Optimizing In Vitro Osteogenesis in Canine Autologous and Induced Pluripotent Stem Cell-Derived Mesenchymal Stromal Cells with Dexamethasone and BMP-2. STEM Cells Dev. 2021, 30, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Mott, A.; Mitchell, A.; McDaid, C.; Harden, M.; Grupping, R.; Dean, A.; Byrne, A.; Doherty, L.; Sharma, H. Systematic review assessing the evidence for the use of stem cells in fracture healing. Bone Jt Open 2020, 1, 628–638. [Google Scholar]
- Desai, S.; Jayasuriya, C.T. Implementation of Endogenous and Exogenous Mesenchymal Progenitor Cells for Skeletal Tissue Regeneration and Repair. Bioengineering 2020, 7, 86. [Google Scholar] [CrossRef]
- Dumont, N.; Boyer, L.; Émond, H.; Celebi-Saltik, B.; Pasha, R.; Bazin, R.; Mantovani, D.; Roy, D.C.; Pineault, N. Medium conditioned with mesenchymal stromal cell-derived osteoblasts improves the expansion and engraftment properties of cord blood progenitors. Exp. Hematol. 2014, 42, 741–752. [Google Scholar] [CrossRef]
- Xie, Y.; Hu, J.H.; Wu, H.; Huang, Z.Z.; Yan, H.W.; Shi, Z.Y. Bone marrow stem cells derived exosomes improve osteoporosis by promoting osteoblast proliferation and inhibiting cell apoptosis. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1214–1220. [Google Scholar]
- Kim, S.J.; Shin, Y.W.; Yang, K.H.; Kim, S.B.; Yoo, M.J.; Han, S.K.; Im, S.A.; Won, Y.D.; Sung, Y.B.; Jeon, T.S.; et al. A multi-center, randomized, clinical study to compare the effect and safety of autologous cultured osteoblast (Ossron) injection to treat fractures. BMC Musculoskelet. Disord. 2009, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Hauzeur, J.P.; Lechanteur, C.; Baudoux, E.; De Maertelaer, V.; Pather, S.; Katz, R.; Malaise, M.; Ino, J.; Beguin, Y. Did Osteoblastic Cell Therapy Improve the Prognosis of Pre-fracture Osteonecrosis of the Femoral Head? A Randomized, Controlled Trial. Clin. Orthop. Relat. Res. 2020, 478, 1307–1315. [Google Scholar] [CrossRef]
- Hu, L.; Yin, C.; Zhao, F.; Ali, A.; Ma, J.; Qian, A. Mesenchymal Stem Cells: Cell Fate Decision to Osteoblast or Adipocyte and Application in Osteoporosis Treatment. Int. J. Mol. Sci. 2018, 19, 360. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.J.; Tang, P.C.; Chen, Y.H.; Chu, F.H.; Kang, T.C.; Chen, L.R.; Yang, J.R. Porcine induced pluripotent stem cell-derived osteoblast-like cells prevent glucocorticoid-induced bone loss in Lanyu pigs. PLoS ONE 2018, 13, e0202155. [Google Scholar] [CrossRef]
- Yamada, M.; Watanabe, J.; Ueno, T.; Ogawa, T.; Egusa, H. Cytoprotective Preconditioning of Osteoblast-Like Cells with N-Acetyl L-Cysteine for Bone Regeneration in Cell Therapy. Int. J. Mol. Sci. 2019, 20, 5199. [Google Scholar] [CrossRef] [Green Version]
- Cahill, R.A.; Jones, O.Y.; Klemperer, M.; Steele, A.; Mueller, T.O.; el-Badri, N.; Chang, Y.; Good, R.A. Replacement of recipient stromal/mesenchymal cells after bone marrow transplantation using bone fragments and cultured osteoblast-like cells. Biol. Blood Marrow Transplant. 2004, 10, 709–717. [Google Scholar] [CrossRef] [Green Version]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal Stem Cell Secretome: Toward Cell-Free Therapeutic Strategies in Regenerative Medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef] [Green Version]
- Squecco, R.; Tani, A.; Chellini, F.; Garella, R.; Idrizaj, E.; Rosa, I.; Zecchi-Orlandini, S.; Manetti, M.; Sassoli, C. Bone Marrow-Mesenchymal Stromal Cell Secretome as Conditioned Medium Relieves Experimental Skeletal Muscle Damage Induced by Ex Vivo Eccentric Contraction. Int. J. Mol. Sci. 2021, 22, 3645. [Google Scholar] [CrossRef] [PubMed]
- d’Angelo, M.; Cimini, A.; Castelli, V. Insights into the Effects of Mesenchymal Stem Cell-Derived Secretome in Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 5241. [Google Scholar] [CrossRef]
- Hernandez-Valladares, M.; Aasebø, E.; Mjaavatten, O.; Vaudel, M.; Bruserud, Ø.; Berven, F.; Selheim, F. Reliable FASP-based procedures for optimal quantitative proteomic and phosphoproteomic analysis on samples from acute myeloid leukemia patients. Biol. Proced. Online 2016, 18, 13. [Google Scholar] [CrossRef] [Green Version]
- Hughes, C.S.; Moggridge, S.; Müller, T.; Sorensen, P.H.; Morin, G.B.; Krijgsveld, J. Single-pot, solid-phase-enhanced sample preparation for proteomics experiments. Nat. Protoc. 2019, 14, 68–85. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Arntzen, M.Ø.; Koehler, C.J.; Barsnes, H.; Berven, F.S.; Treumann, A.; Thiede, B. IsobariQ: Software for isobaric quantitative proteomics using IPTL, iTRAQ, and TMT. J. Proteome Res. 2011, 10, 913–920. [Google Scholar] [CrossRef]
- Scholz, C.; Lyon, D.; Refsgaard, J.C.; Jensen, L.J.; Choudhary, C.; Weinert, B.T. Avoiding abundance bias in the functional annotation of post-translationally modified proteins. Nat. Methods 2015, 12, 1003–1004. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Bader, G.D.; Hogue, C.W. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Aasebø, E.; Brenner, A.K.; Birkeland, E.; Tvedt, T.H.A.; Selheim, F.; Berven, F.S.; Bruserud, Ø. The Constitutive Extracellular Protein Release by Acute Myeloid Leukemia Cells-A Proteomic Study of Patient Heterogeneity and Its Modulation by Mesenchymal Stromal Cells. Cancers 2021, 13, 1509. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO Term | Name | Matching Protein in Osteoblasts Only |
|---|---|---|
| GO:0031012 (198/700) | Extracellular matrix (CC) | 21 proteins: ANGPTL3, APOA1, APOE, BCAM, CHI3L1, COL5A3, CRISPLD2, CXCL12, DMBT1, ELN, F9, FBN2, FGG, LGALS9, LRRC32, MFAP2, MUC5B, NTN4, PODN, TFPI2, TINAGL1 |
| GO:0030198 (159/404) | Extracellular matrix organization (BP) | 8 proteins: COL5A3, CRISPLD2, ELN, FBN2, FGG, MFAP2, NTN4, SPP1 |
| GO:0001503 (105/297) | Ossification (BP) | SPP1, TRAF6 |
| GO:0030278 (29/122) | Regulation of ossification | FBN2 |
| GO:0045667 (39/127) | Regulation of osteoblast differentiation | FBN2, HDAC7, JAG1, VEGFC |
| GO:0007229 (48/203) | Integrin-mediated signaling pathway | ANGPTL3, APOA1 |
| GO:2001044 (10/15) | Regulation of integrin-mediated signaling pathway | LIMS2 |
| GO:0016055 (140/442) | Wnt signaling pathway | DAAM2, MAP3K7 |
| GO:0030111 (144/406) | Regulation of Wnt signaling pathway | APOE, DAAM2, IGFBP2 |
| GO:0007219 (34/165) | Notch signaling pathway | JAG1, PSEN2 |
| GO:0008593 (27/110) | Regulation of Notch signaling pathway | JAG1 |
| Uniprot Keyword | S-Value | Foreground Count | FDR |
|---|---|---|---|
| Signal | 0.69 | 25 | 0.0008 |
| Disulfide bond | 0.66 | 24 | 0.0008 |
| Nucleus | −0.57 | 6 | 0.003 |
| Glycoprotein | 0.55 | 25 | 0.002 |
| Phosphoprotein | −0.45 | 24 | 0.040 |
| Acetylation | −0.45 | 13 | 0.022 |
| Secreted | 0.39 | 16 | 0.002 |
| Cell membrane | 0.24 | 18 | 0.047 |
| Adaptive immunity | 0.17 | 6 | 0.003 |
| Donors | Gender | Age (Years) | Lot Number |
|---|---|---|---|
| Osteoblasts | |||
| hOB Donor 1 | Male | 54 | 422Z050 |
| hOB Donor 2 | Male | 58 | 422Z047.2 |
| hOB Donor 3 | Male | 63 | 415Z007.2 |
| hOB Donor 4 | Female | 71 | 443Z004.2 |
| hOB Donor 5 | Female | 62 | 413Z026.2 |
| * hOB Donor 6 | Female | 73 | 427Z010.2 |
| * hOB Donor 7 | Male | 58 | 422Z051 |
| * hOB Donor 8 | Male | 64 | 427Z036 |
| * hOB Donor 9 | Female | 64 | 422Z031.2 |
| * hOB Donor 10 | Female | 56 | 415Z011 |
| MSCs | |||
| MSC Donor 1 | Male | 47 | 402Z027 |
| MSC Donor 2 | Male | 62 | 413Z021.4 |
| MSC Donor 3 | Male | 63 | 411Z011.4 |
| MSC Donor 4 | Female | 57 | 409Z018.1 |
| MSC Donor 5 | Female | 66 | 421Z029.3 |
| * MSC Donor 6 | Male | 70 | 429Z022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aasebø, E.; Brenner, A.K.; Hernandez-Valladares, M.; Birkeland, E.; Berven, F.S.; Selheim, F.; Bruserud, Ø. Proteomic Comparison of Bone Marrow Derived Osteoblasts and Mesenchymal Stem Cells. Int. J. Mol. Sci. 2021, 22, 5665. https://doi.org/10.3390/ijms22115665
Aasebø E, Brenner AK, Hernandez-Valladares M, Birkeland E, Berven FS, Selheim F, Bruserud Ø. Proteomic Comparison of Bone Marrow Derived Osteoblasts and Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2021; 22(11):5665. https://doi.org/10.3390/ijms22115665
Chicago/Turabian StyleAasebø, Elise, Annette K. Brenner, Maria Hernandez-Valladares, Even Birkeland, Frode S. Berven, Frode Selheim, and Øystein Bruserud. 2021. "Proteomic Comparison of Bone Marrow Derived Osteoblasts and Mesenchymal Stem Cells" International Journal of Molecular Sciences 22, no. 11: 5665. https://doi.org/10.3390/ijms22115665
APA StyleAasebø, E., Brenner, A. K., Hernandez-Valladares, M., Birkeland, E., Berven, F. S., Selheim, F., & Bruserud, Ø. (2021). Proteomic Comparison of Bone Marrow Derived Osteoblasts and Mesenchymal Stem Cells. International Journal of Molecular Sciences, 22(11), 5665. https://doi.org/10.3390/ijms22115665