
Molecular Analysis of ZNF71 KRAB in Non-Small-Cell Lung Cancer

 , ,
, ,  ,
,  , and
, and
Abstract
:1. Introduction
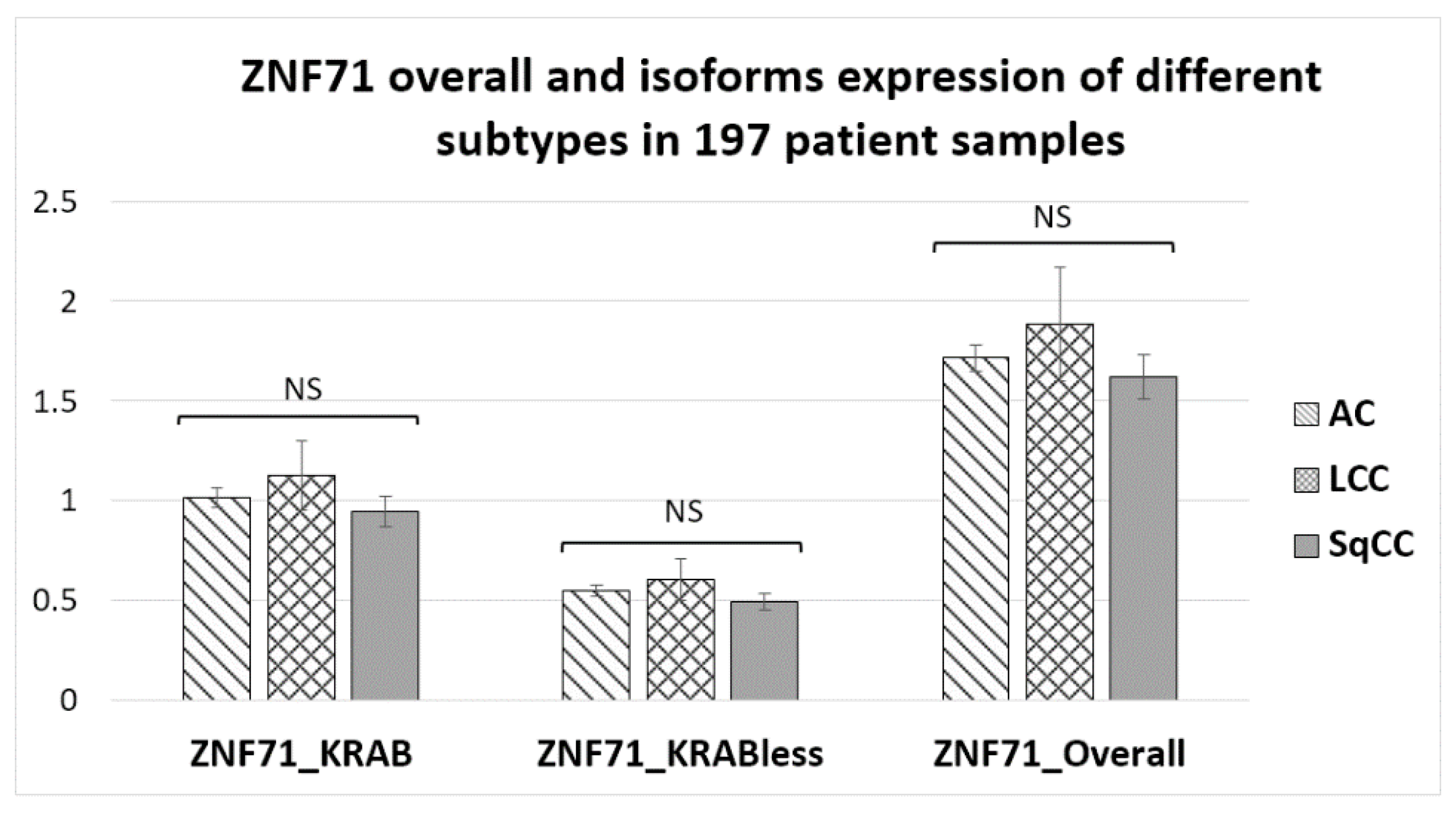
2. Results
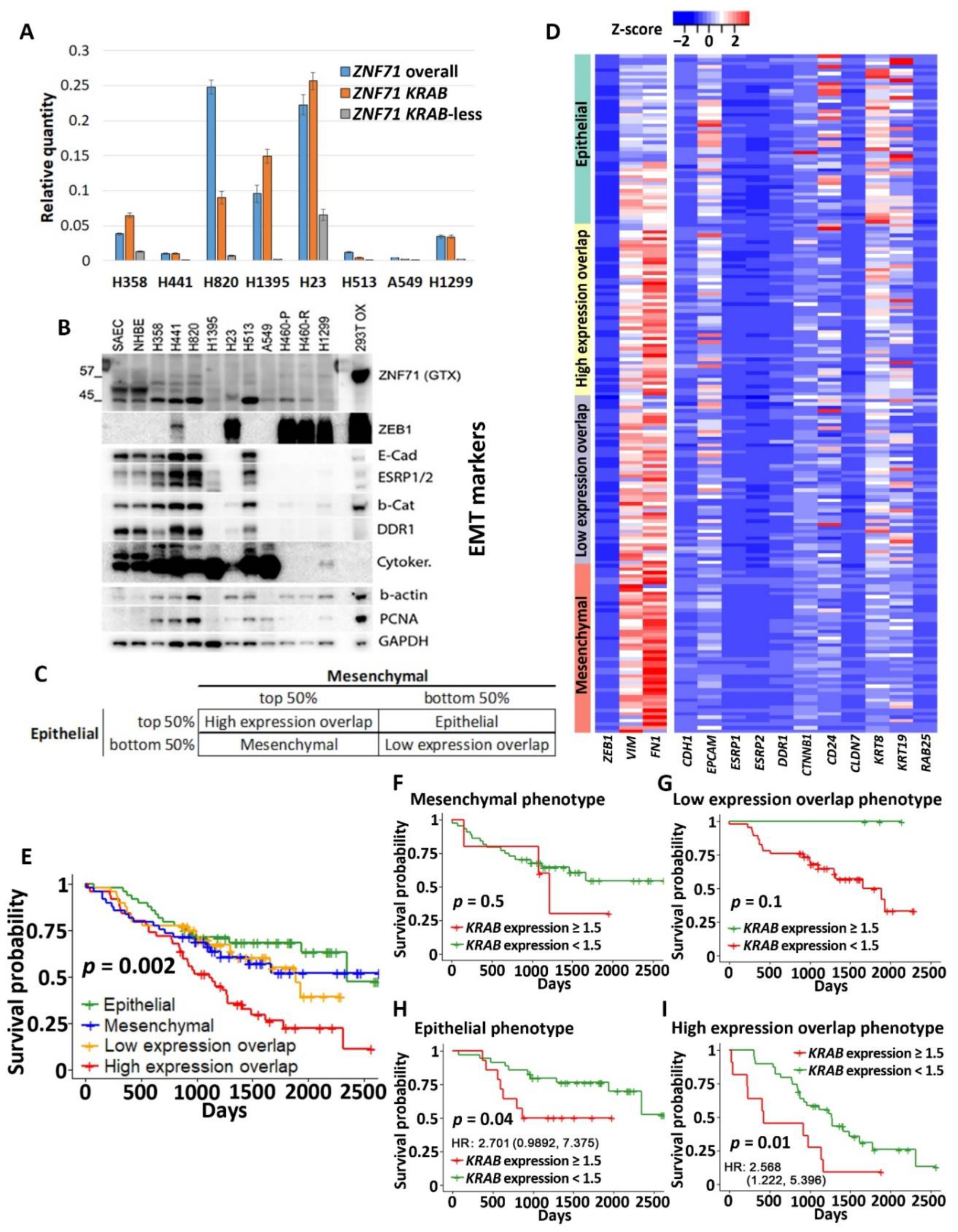
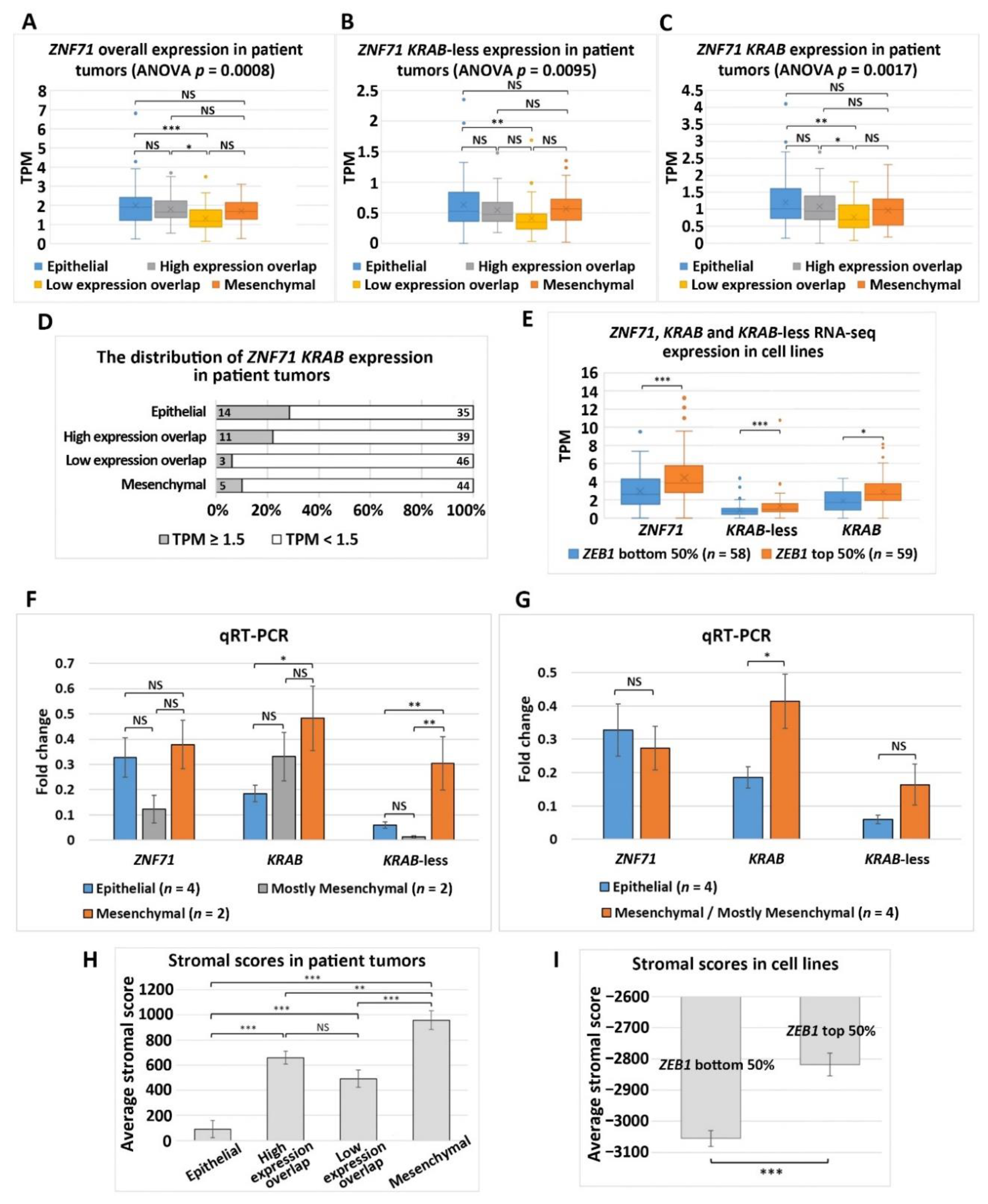
2.1. Expression of ZNF71 Isoforms in NSCLC Tumors and Cell Lines
2.2. Association of ZNF71 KRAB Isoform with EMT
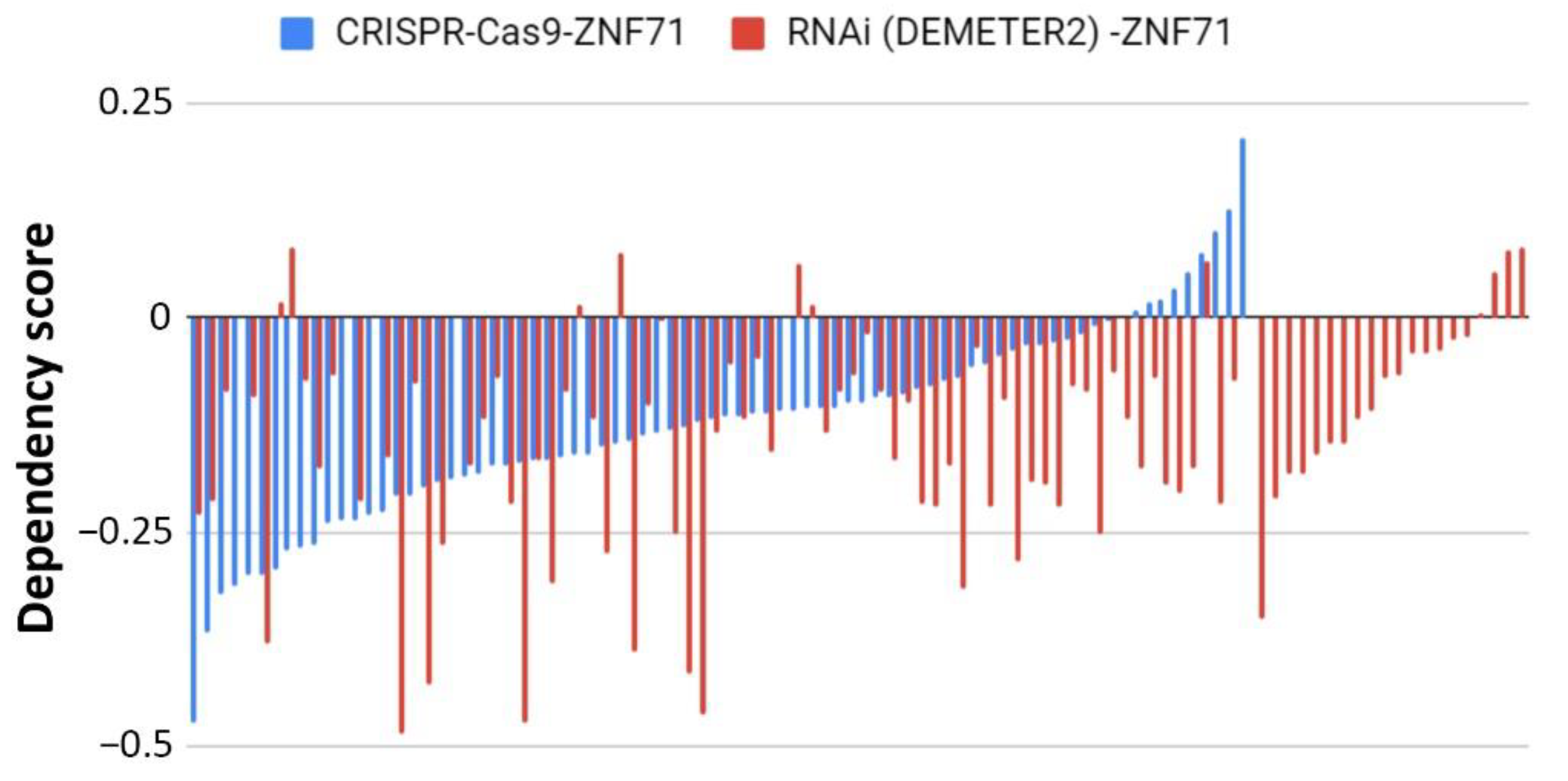
2.3. Functional Analysis of ZNF71 in NSCLC Cell Lines
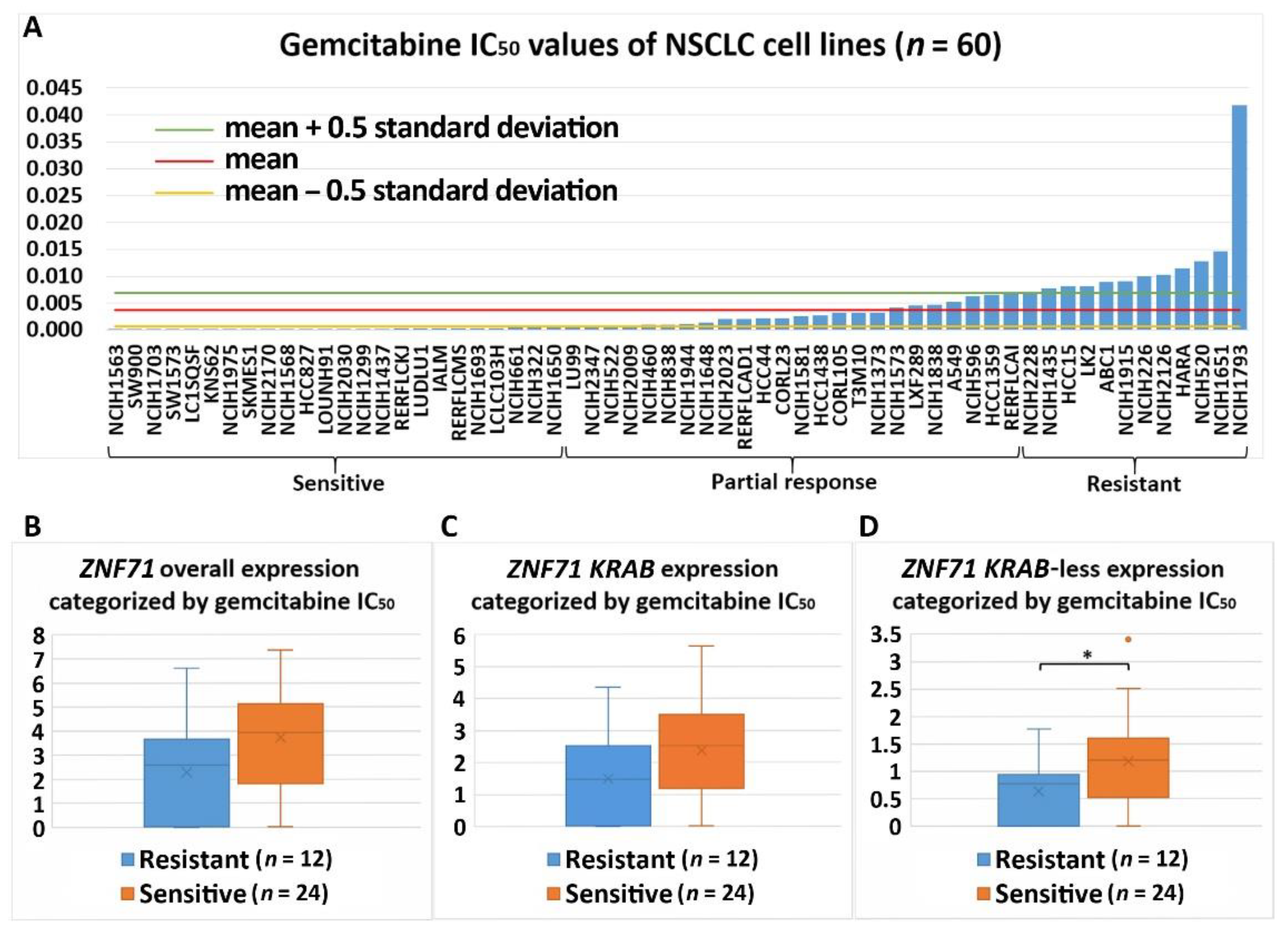
2.4. Association of ZNF71 Isoforms with Chemoresponse
3. Discussion
4. Materials and Methods
4.1. NSCLC Patient Samples and RNA-Seq Data
4.2. Cancer Cell Line Encyclopedia (CCLE)
4.3. CRISPR-Cas9 Assays
4.4. RNAi Functional Assays
4.5. Drug Response
4.6. RNA Extraction, Quality Assessment, and qRT-PCR
4.7. Western Blots
4.8. Responses to Taxol with Annexin V Staining for Apoptosis Using Flow Cytometry
4.9. Statistical Analysis
4.10. Assessment of Stromal Infiltration and Immune Infiltration
4.11. ZNF71 Overexpression
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Variable | Count | |
|---|---|---|
| Stage | 1a | 70 |
| 1b | 44 | |
| 2a | 25 | |
| 2b | 23 | |
| 3a | 32 | |
| 4 | 3 | |
| Histology | Adenocarcinoma | 108 |
| Large cell carcinoma | 23 | |
| Squamous cell carcinoma | 66 | |
| Gender | Female | 102 |
| Male | 95 | |
| Status | Alive | 104 |
| Dead | 93 | |
| Smoking | Current | 94 |
| Ex > 1 year | 84 | |
| Never | 19 | |
| EMT Phenotype | |||||
|---|---|---|---|---|---|
| Stage | Epithelial | High Expression Overlap | Low Expression Overlap | Mesenchymal | Total |
| 1 | 28 | 30 | 28 | 28 | 114 |
| 2 | 11 | 11 | 12 | 14 | 48 |
| 3 | 9 | 7 | 9 | 7 | 32 |
| 4 | 1 | 2 | 0 | 0 | 3 |
| Total | 49 | 50 | 49 | 49 | 197 |
| EMT Phenotype | |||||
|---|---|---|---|---|---|
| ZNF71 KRAB Expression | Epithelial | High Expression Overlap | Low Expression Overlap | Mesenchymal | Total |
| TPM ≥ 1.5 | 14 | 11 | 3 | 5 | 33 |
| TPM < 1.5 | 35 | 39 | 46 | 44 | 164 |
| Total | 49 | 50 | 49 | 49 | 197 |
| Correlation with Immune Scores | Correlation with Stromal Scores | Partial Correlation with Immune Scores (Removing the Effect of Stromal Scores) | Partial Correlation with Stromal Scores (Removing the Effect of Immune Scores) | |
|---|---|---|---|---|
| ZEB1 | 0.13 | 0.55 *** | −0.235 *** | 0.573 *** |
| VIM | 0.21 ** | 0.3 *** | 0 | 0.447 *** |
| FN1 | 0.11 | 0.34 *** | −0.29 *** | 0.595 *** |
| CD24 | −0.23 ** | −0.2 ** | −0.175 * | 0.022 |
| CDH1 | −0.18 * | −0.12 | −0.157 * | −0.05 |
| CLDN7 | −0.12 | −0.12 | −0.07 | −0.079 |
| CTNNB1 | −0.083 | 0.041 | −0.189 ** | 0.19 ** |
| DDR1 | −0.38 *** | −0.36 *** | −0.29 *** | −0.137 |
| EPCAM | −0.24 *** | −0.25 *** | −0.167 * | −0.178 * |
| ESRP1 | −0.56 *** | −0.42 *** | −0.404 *** | −0.184 ** |
| ESRP2 | −0.32 ** | −0.25 *** | −0.117 | −0.159 * |
| KRT8 | −0.32 ** | −0.19 ** | −0.114 | −0.103 |
| KRT19 | −0.17 * | −0.28 *** | 0.043 | −0.089 |
| RAB25 | −0.25 *** | −0.26 *** | −0.151 * | −0.175 * |
| ZNF71 overall | −0.12 | −0.17 * | 0.015 | −0.059 |
| ZNF71 KRAB | −0.11 | −0.16 * | −0.157 | −0.055 |
| ZNF71 KRAB-less | −0.085 | −0.11 | −0.114 | 0.005 |
| Cell Line | Location of the EGFR Mutation | EGFR Variant Classification |
|---|---|---|
| HCC1359 | chr7: 55087037-55087037 | Missense_Mutation |
| HCC4006 | chr7: 55242466-55242474 | In_Frame_Del |
| HCC827 | chr7: 55242466-55242480 | In_Frame_Del |
| LOUNH91 | chr7: 55242470-55242487 | In_Frame_Del |
| chr7: 55242494-55242494 | Missense_Mutation | |
| NCIH1355 | chr7: 55273154-55273154 | Missense_Mutation |
| NCIH1568 | chr7: 55233029-55233029 | Silent |
| chr7: 55273056-55273056 | Missense_Mutation | |
| NCIH1650 | chr7: 55242465-55242479 | In_Frame_Del |
| NCIH1793 | chr7: 55223565-55223565 | Missense_Mutation |
| NCIH1975 | chr7: 55249071-55249071 | Missense_Mutation |
| chr7: 55259515-55259515 | Missense_Mutation | |
| NCIH226 | chr7: 55223558-55223558 | Missense_Mutation |
| PC14 | chr7: 55242465-55242479 | In_Frame_Del |
| SKMES1 | chr7: 55240751-55240751 | Silent |
| chr7: 55223558-55223558 | Missense_Mutation |
| Sensitive | Resistant | ||
|---|---|---|---|
| Gefitinib | IC50 | HCC4006, HCC827, LOUNH91, NCIH1355, NCIH1568 | HCC1359, NCIH1793, NCIH226, SKMES1 |
| EC50 | HCC4006, HCC827, NCIH1355, NCIH1568, NCIH1975, NCIH226, PC14 | HCC1359, SKMES1 | |
| Erlotinib | IC50 | HCC4006, HCC827, NCIH1355, NCIH1650, NCIH1793 | |
| EC50 | HCC1359, HCC4006, HCC827, NCIH1355, NCIH1650, NCIH1975, PC14 | NCIH226, SKMES1 | |


References
- Lung Cancer-Non-Small Cell: Statistics. Available online: https://www.cancer.net/cancer-types/lung-cancer-non-small-cell/statistics and https://www.cancer.net/cancer-types/melanoma/statistics (accessed on 2 October 2020).
- Hoffman, P.C.; Mauer, A.M.; Vokes, E.E. Lung cancer. Lancet 2000, 355, 479–485. [Google Scholar] [CrossRef]
- Zheng, Y.; Jaklitsch, M.T.; Bueno, R. Neoadjuvant Therapy in Non-Small Cell Lung Cancer. Surg. Oncol. Clin. North Am. 2016, 25, 567–584. [Google Scholar] [CrossRef]
- Crinò, L.; Weder, W.; Van Meerbeeck, J.; Felip, E. Early stage and locally advanced (non-metastatic) non-small-cell lung cancer: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2010, 21, v103–v115. [Google Scholar] [CrossRef] [PubMed]
- Byron, E.; Pinder-Schenck, M. Systemic and targeted therapies for early-stage lung cancer. Cancer Control. 2014, 21, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Guo, N.L.; Dowlati, A.; Raese, R.A.; Dong, C.; Chen, G.; Beer, D.G.; Shaffer, J.; Singh, S.; Bokhary, U.; Liu, L.; et al. A Predictive 7-Gene Assay and Prognostic Protein Biomarkers for Non-small Cell Lung Cancer. EBioMedicine 2018, 32, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Laity, J.H.; Lee, B.M.; Wright, P.E. Zinc finger proteins: New insights into structural and functional diversity. Curr. Opin. Struct. Biol. 2001, 11, 39–46. [Google Scholar] [CrossRef]
- Cassandri, M.; Smirnov, A.; Novelli, F.; Pitolli, C.; Agostini, M.; Malewicz, M.; Melino, G.; Raschellà, G. Zinc-finger proteins in health and disease. Cell Death Discov. 2017, 3, 17071. [Google Scholar] [CrossRef] [Green Version]
- Huntley, S.; Baggott, D.M.; Hamilton, A.T.; Tran-Gyamfi, M.; Yang, S.; Kim, J.; Gordon, L.; Branscomb, E.; Stubbs, L. A comprehensive catalog of human KRAB-associated zinc finger genes: Insights into the evolutionary history of a large family of transcriptional repressors. Genome Res. 2006, 16, 669–677. [Google Scholar] [CrossRef] [Green Version]
- Bruno, M.; Mahgoub, M.; Macfarlan, T.S. The Arms Race Between KRAB-Zinc Finger Proteins and Endogenous Retroelements and Its Impact on Mammals. Annu. Rev. Genet. 2019, 53, 393–416. [Google Scholar] [CrossRef]
- Ecco, G.; Imbeault, M.; Trono, D. KRAB zinc finger proteins. Development 2017, 144, 2719–2729. [Google Scholar] [CrossRef] [Green Version]
- Lupo, A.; Cesaro, E.; Montano, G.; Zurlo, D.; Izzo, P.; Costanzo, P. KRAB-Zinc Finger Proteins: A Repressor Family Displaying Multiple Biological Functions. Curr. Genom. 2013, 14, 268–278. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T.; Mataki, C.; Nagao, C.; Umetani, M.; Wada, Y.; Ishii, M.; Tsutsumi, S.; Kohro, T.; Saiura, A.; Aburatani, H.; et al. The Gene Expression Profile of Human Umbilical Vein Endothelial Cells Stimulated by Tumor Necrosis Factor a using DNA Microarray Analysis. J. Atheroscler. Thromb. 2000, 7, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-H.; Cheong, H.S.; Park, J.S.; Jang, A.-S.; Uh, S.-T.; Kim, Y.-H.; Kim, M.-K.; Choi, I.S.; Cho, S.H.; Choi, B.W.; et al. A Genome-Wide Association Study of Total Serum and Mite-Specific IgEs in Asthma Patients. PLoS ONE 2013, 8, e71958. [Google Scholar] [CrossRef]
- Roche, J. The Epithelial-to-Mesenchymal Transition in Cancer. Cancers 2018, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Nieto, M.A.; Huang, R.Y.-J.; Jackson, R.A.; Thiery, J.P. EMT: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging Biological Principles of Metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [Green Version]
- Moustakas, A.; De Herreros, A.G. Epithelial-mesenchymal transition in cancer. Mol. Oncol. 2017, 11, 715–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, C.J.; Dyson, G.; Campeanu, I.J.; Watza, D.; Ratnam, M.; Wu, G. A stroma-corrected ZEB1 transcriptional signature is inversely associated with antitumor immune activity in breast cancer. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshihara, K.; Shahmoradgoli, M.; Martínez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Treviño, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Azeredo-Tseng, C.; Luo, M.; Randall, N.; Hong, T. Integrative Transcriptomic Analysis Reveals a Multiphasic Epithelial-Mesenchymal Spectrum in Cancer and Non-tumorigenic Cells. Front. Oncol. 2020, 9, 1479. [Google Scholar] [CrossRef]
- Friedman, J.R.; Fredericks, W.J.E.; Jensen, D.; Speicher, D.W.; Huang, X.P.; Neilson, E.G.; Rauscher, F.J. KAP-1, a novel corepressor for the highly conserved KRAB repression domain. Genes Dev. 1996, 10, 2067–2078. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Munoz-Antonia, T.; Cress, W.D. Trim28 Contributes to EMT via Regulation of E-Cadherin and N-Cadherin in Lung Cancer Cell Lines. PLoS ONE 2014, 9, e101040. [Google Scholar] [CrossRef]
- Pei, L.; He, X.; Li, S.; Sun, R.; Xiang, Q.; Ren, G.; Xiang, T. KRAB zinc-finger protein 382 regulates epithelial-mesenchymal transition and functions as a tumor suppressor but is silenced by CpG methylation in gastric cancer. Int. J. Oncol. 2018, 53, 961–972. [Google Scholar] [CrossRef] [Green Version]
- Mezheyeuski, A.; Bergsland, C.H.; Backman, M.; Djureinovic, D.; Sjöblom, T.; Bruun, J.; Micke, P. Multispectral imaging for quantitative and compartment-specific immune infiltrates reveals distinct immune profiles that classify lung cancer patients. J. Pathol. 2018, 244, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Broad Institute. DepMap 20Q2 Public. In DepMap; Broad Institute: Cambridge, MA, USA, 2020. [Google Scholar]
- Meyers, R.M.; Bryan, J.G.; McFarland, J.M.; Weir, B.A.; Sizemore, A.E.; Xu, H.; Dharia, N.V.; Montgomery, P.G.; Cowley, G.S.; Pantel, S.; et al. Computational correction of copy number effect improves specificity of CRISPR-Cas9 essentiality screens in cancer cells. Nat. Genet. 2017, 49, 1779–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFarland, J.M.; Ho, Z.V.; Kugener, G.; Dempster, J.M.; Montgomery, P.G.; Bryan, J.G.; Krill-Burger, J.M.; Green, T.M.; Vazquez, F.; Boehm, J.S.; et al. Improved estimation of cancer dependencies from large-scale RNAi screens using model-based normalization and data integration. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuse, K. Platinum/Oral Etoposide Therapy in Non-Small Cell Lung Cancer. Oncology 1992, 49, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Steuer, C.E.; Behera, M.; Ernani, V.; Higgins, K.A.; Saba, N.F.; Shin, D.M.; Pakkala, S.; Pillai, R.N.; Owonikoko, T.K.; Curran, W.J.; et al. Comparison of Concurrent use of Thoracic Radiation with Either Carboplatin-Paclitaxel or Cisplatin-Etoposide for Patients with Stage III Non-Small-Cell Lung Cancer: A Systematic Review. JAMA Oncol. 2017, 3, 1120–1129. [Google Scholar] [CrossRef]
- Yang, Z.; Hackshaw, A.; Feng, Q.; Fu, X.; Zhang, Y.; Mao, C.; Tang, J. Comparison of gefitinib, erlotinib and afatinib in non-small cell lung cancer: A meta-analysis. Int. J. Cancer 2017, 140, 2805–2819. [Google Scholar] [CrossRef]
- Fossella, F.V. Docetaxel for previously treated non-small-cell lung cancer. Oncology 2002, 16 (Suppl. 6), 45–51. [Google Scholar]
- Arrieta, O.; Barrón, F.; Ramírez-Tirado, L.A.; Zatarain-Barrón, Z.L.; Cardona, A.F.; Díaz-García, D.; Yamamoto Ramos, M.; Mota-Vega, B.; Carmona, A.; Peralta Álvarez, M.P.; et al. Efficacy and Safety of Pembrolizumab Plus Docetaxel vs Docetaxel Alone in Patients with Previously Treated Advanced Non-Small Cell Lung Cancer: The PROLUNG Phase 2 Randomized Clinical Trial. JAMA Oncol. 2020, 6, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Kurata, T.; Nakagawa, K. Gemcitabine: Efficacy in the Treatment of Advanced Stage Nonsquamous Non-Small Cell Lung Cancer. Clin. Med. Insights Oncol. 2011, 5. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, S.; Belani, C.P. Paclitaxel for non-small cell lung cancer. Expert Opin. Pharmacother. 2004, 5, 1771–1780. [Google Scholar] [CrossRef] [PubMed]
- Pearce, M.C.; Gamble, J.T.; Kopparapu, P.R.; O’Donnell, E.F.; Mueller, M.J.; Jang, H.S.; Greenwood, J.A.; Satterthwait, A.C.; Tanguay, R.L.; Zhang, X.-K.; et al. Induction of apoptosis and suppression of tumor growth by Nur77-derived Bcl-2 converting peptide in chemoresistant lung cancer cells. Oncotarget 2018, 9, 26072–26085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnio, S.; Novello, S.; Papotti, M.; Iacono, M.L.; Scagliotti, G.V. Prognostic and predictive biomarkers in early stage non-small cell lung cancer: Tumor based approaches including gene signatures. Transl. Lung Cancer Res. 2013, 2, 372–381. [Google Scholar] [PubMed]
- Kosaka, T.; Yatabe, Y.; Onozato, R.; Kuwano, H.; Mitsudomi, T. Prognostic Implication of EGFR, KRAS, and TP53 Gene Mutations in a Large Cohort of Japanese Patients with Surgically Treated Lung Adenocarcinoma. J. Thorac. Oncol. 2009, 4, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pécuchet, N.; Laurent-Puig, P.; Mansuet-Lupo, A.; Legras, A.; Alifano, M.; Pallier, K.; Didelot, A.; Gibault, L.; Danel, C.; Just, P.-A.; et al. Different prognostic impact of STK11 mutations in non-squamous non-small-cell lung cancer. Oncotarget 2015, 8, 23831–23840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lord, R.V.N.; Brabender, J.; Gandara, D.; Alberola, V.; Camps, C.; Domine, M.; Cardenal, F.; Sánchez, J.M.; Gumerlock, P.H.; Tarón, M.; et al. Low ERCC1 expression correlates with prolonged survival after cisplatin plus gemcitabine chemotherapy in non-small cell lung cancer. Clin. Cancer Res. 2002, 8, 2286–2291. [Google Scholar]
- Simon, G.R.; Sharma, S.; Cantor, A.; Smith, P.; Bepler, G. ERCC1 Expression Is a Predictor of Survival in Resected Patients with Non-small Cell Lung Cancer. Chest 2005, 127, 978–983. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.-Q.; Ding, K.; Strumpf, D.; Weir, B.A.; Meyerson, M.; Pennell, N.; Thomas, R.K.; Naoki, K.; Ladd-Acosta, C.; Liu, N.; et al. Prognostic and Predictive Gene Signature for Adjuvant Chemotherapy in Resected Non-Small-Cell Lung Cancer. J. Clin. Oncol. 2010, 28, 4417–4424. [Google Scholar] [CrossRef]
- Chen, D.-T.; Hsu, Y.-L.; Fulp, W.J.; Coppola, D.; Haura, E.B.; Yeatman, T.J.; Cress, W.D. Prognostic and Predictive Value of a Malignancy-Risk Gene Signature in Early-Stage Non-Small Cell Lung Cancer. J. Natl. Cancer Inst. 2011, 103, 1859–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratz, J.R.; He, J.; Eeden, S.K.V.D.; Zhu, Z.-H.; Gao, W.; Pham, P.T.; Mulvihill, M.S.; Ziaei, F.; Zhang, H.; Su, B.; et al. A practical molecular assay to predict survival in resected non-squamous, non-small-cell lung cancer: Development and international validation studies. Lancet 2012, 379, 823–832. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Xiao, G.; Behrens, C.; Schiller, J.; Allen, J.; Chow, C.-W.; Suraokar, M.; Corvalan, A.; Mao, J.; White, M.A.; et al. A 12-Gene Set Predicts Survival Benefits from Adjuvant Chemotherapy in Non-Small Cell Lung Cancer Patients. Clin. Cancer Res. 2013, 19, 1577–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodard, G.A.; Wang, S.X.; Kratz, J.R.; Zoon-Besselink, C.T.; Chiang, C.-Y.; Gubens, M.A.; Jahan, T.M.; Blakely, C.M.; Jones, K.D.; Mann, M.J.; et al. Adjuvant Chemotherapy Guided by Molecular Profiling and Improved Outcomes in Early Stage, Non-Small-Cell Lung Cancer. Clin. Lung Cancer 2018, 19, 58–64. [Google Scholar] [CrossRef]
- Houston, K.A.; Henley, S.J.; Li, J.; White, M.C.; Richards, T.B. Patterns in lung cancer incidence rates and trends by histologic type in the United States, 2004–2009. Lung Cancer 2014, 86, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Bueno, R. Commercially available prognostic molecular models in early-stage lung cancer: A review of the Pervenio Lung RS and Myriad myPlan Lung Cancer tests. Expert Rev. Mol. Diagn. 2015, 15, 589–596. [Google Scholar] [CrossRef]
- Myriad. All Products. Available online: https://myriad.com/products-services/all-products/overview (accessed on 31 December 2020).
- Klug, A. The Discovery of Zinc Fingers and Their Applications in Gene Regulation and Genome Manipulation. Annu. Rev. Biochem. 2010, 79, 213–231. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.; Wang, Y.; Macfarlan, T.S. The Role of KRAB-ZFPs in Transposable Element Repression and Mammalian Evolution. Trends Genet. 2017, 33, 871–881. [Google Scholar] [CrossRef]
- Addison, J.B.; Koontz, C.; Fugett, J.H.; Creighton, C.J.; Chen, N.; Farrugia, M.K.; Padon, R.R.; Voronkova, M.A.; McLaughlin, S.L.; Livengood, R.H.; et al. KAP1 promotes proliferation and metastatic progression of breast cancer cells. Cancer Res. 2014, 75, 344–355. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.; Kong, J.-W.-F.; Choong, L.-Y.; Loh, M.-C.-S.; Toy, W.; Chong, P.-K.; Wong, C.-H.; Wong, C.-Y.; Shah, N.; Lim, Y.-P. Novel Breast Cancer Metastasis-Associated Proteins. J. Proteome Res. 2009, 8, 583–594. [Google Scholar] [CrossRef]
- Zhang, X.; Jiang, G.; Wu, J.; Zhou, H.; Zhang, Y.; Miao, Y.; Feng, Y.; Yu, J. Zinc finger protein 668 suppresses non-small cell lung cancer invasion and migration by downregulating Snail and upregulating E-cadherin and zonula occludens-1. Oncol. Lett. 2018, 15, 3806–3813. [Google Scholar] [CrossRef]
- Tulchinsky, E.; Demidov, O.; Kriajevska, M.; Barlev, N.A.; Imyanitov, E. EMT: A mechanism for escape from EGFR-targeted therapy in lung cancer. Biochim. Biophys. Acta (BBA) Bioenerg. 2019, 1871, 29–39. [Google Scholar] [CrossRef]
- Warzecha, C.C.; Carstens, R.P. Complex changes in alternative pre-mRNA splicing play a central role in the epithelial-to-mesenchymal transition (EMT). Semin. Cancer Biol. 2012, 22, 417–427. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Choi, P.S.; Chaffer, C.L.; Labella, K.; Hwang, J.H.O.; Giacomelli, A.; Kim, J.W.; Ilic, N.; Doench, J.G.; Ly, S.H.; et al. An alternative splicing switch in FLNB promotes the mesenchymal cell state in human breast cancer. Elife 2018, 7. [Google Scholar] [CrossRef]
- Chen, L.; Chen, D.-T.; Kurtyka, C.; Rawal, B.; Fulp, W.J.; Haura, E.B.; Cress, W. Tripartite Motif Containing 28 (Trim28) Can Regulate Cell Proliferation by Bridging HDAC1/E2F Interactions. J. Biol. Chem. 2012, 287, 40106–40118. [Google Scholar] [CrossRef] [Green Version]
- Czerwińska, P.; Mazurek, S.; Wiznerowicz, M. The complexity of TRIM28 contribution to cancer. J. Biomed. Sci. 2017, 24, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sobocińska, J.; Molenda, S.; Machnik, M.; Oleksiewicz, U. KRAB-ZFP Transcriptional Regulators Acting as Oncogenes and Tumor Suppressors: An Overview. Int. J. Mol. Sci. 2021, 22, 2212. [Google Scholar] [CrossRef]
- Huang, C.; Jia, Y.; Yang, S.; Chen, B.; Sun, H.; Shen, F.; Wang, Y. Characterization of ZNF23, a KRAB-containing protein that is downregulated in human cancers and inhibits cell cycle progression. Exp. Cell Res. 2007, 313, 254–263. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Wang, B.; Wang, H.; Zhang, C.; Wu, Y.; Fu, L.; Yu, G. RBAK is upregulated in non-small cell lung cancer and promotes cell migration and invasion. Exp. Ther. Med. 2019, 18, 2942–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Aguet, F. TOPMed RNA-Seq Pipeline Harmonization Summary. Available online: https://github.com/broadinstitute/gtex-pipeline/blob/master/TOPMed_RNAseq_pipeline.md (accessed on 24 March 2021).
- Dempster, J.M.; Rossen, J.; Kazachkova, M.; Pan, J.; Kugener, G.; Root, D.E.; Tsherniak, A. Extracting Biological Insights from the Project Achilles Genome-Scale CRISPR Screens in Cancer Cell Lines. BioRxiv 2019. [Google Scholar] [CrossRef]
- Corsello, S.M.; Nagari, R.T.; Spangler, R.D.; Rossen, J.; Kocak, M.; Bryan, J.G.; Humeidi, R.; Peck, D.; Wu, X.; Tang, A.A.; et al. Non-oncology drugs are a source of previously unappreciated anti-cancer activity. BioRxiv 2019. [Google Scholar] [CrossRef]
- Ma, Y.; Ding, Z.; Qian, Y.; Shi, X.; Castranova, V.; Harner, E.J.; Guo, L. Predicting Cancer Drug Response by Proteomic Profiling. Clin. Cancer Res. 2006, 12, 4583–4589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, N.I.; Ma, Y.; Ding, Z.; Qian, Y.; Wan, Y.-W.; Tosun, K.; Shi, X.; Castranova, V.; Harner, E.J. An integrative genomic and proteomic approach to chemosensitivity prediction. Int. J. Oncol. 1992, 34, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, L.H.; Litière, S.; De Vries, E.; Ford, R.; Gwyther, S.; Mandrekar, S.; Shankar, L.; Bogaerts, J.; Chen, A.; Dancey, J.; et al. RECIST 1.1—Update and clarification: From the RECIST committee. Eur. J. Cancer 2016, 62, 132–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addison, J.B.; Voronkova, M.A.; Fugett, J.H.; Lin, C.-C.; Linville, N.C.; Trinh, B.; Livengood, R.H.; Smolkin, M.B.; Schaller, M.D.; Ruppert, J.M.; et al. Functional Hierarchy and Cooperation of EMT Master Transcription Factors in Breast Cancer Metastasis. Mol. Cancer Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- RS Team. RStudio: Integrated Development for R; RStudio Inc.: Boston, MA, USA, 2019; Available online: http://www.rstudio.com (accessed on 1 April 2021).










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Q.; Mohamed, R.; Dakhlallah, D.; Gencheva, M.; Hu, G.; Pearce, M.C.; Kolluri, S.K.; Marsh, C.B.; Eubank, T.D.; Ivanov, A.V.; et al. Molecular Analysis of ZNF71 KRAB in Non-Small-Cell Lung Cancer. Int. J. Mol. Sci. 2021, 22, 3752. https://doi.org/10.3390/ijms22073752
Ye Q, Mohamed R, Dakhlallah D, Gencheva M, Hu G, Pearce MC, Kolluri SK, Marsh CB, Eubank TD, Ivanov AV, et al. Molecular Analysis of ZNF71 KRAB in Non-Small-Cell Lung Cancer. International Journal of Molecular Sciences. 2021; 22(7):3752. https://doi.org/10.3390/ijms22073752
Chicago/Turabian StyleYe, Qing, Rehab Mohamed, Duaa Dakhlallah, Marieta Gencheva, Gangqing Hu, Martin C. Pearce, Siva Kumar Kolluri, Clay B. Marsh, Timothy D. Eubank, Alexey V. Ivanov, and et al. 2021. "Molecular Analysis of ZNF71 KRAB in Non-Small-Cell Lung Cancer" International Journal of Molecular Sciences 22, no. 7: 3752. https://doi.org/10.3390/ijms22073752
APA StyleYe, Q., Mohamed, R., Dakhlallah, D., Gencheva, M., Hu, G., Pearce, M. C., Kolluri, S. K., Marsh, C. B., Eubank, T. D., Ivanov, A. V., & Guo, N. L. (2021). Molecular Analysis of ZNF71 KRAB in Non-Small-Cell Lung Cancer. International Journal of Molecular Sciences, 22(7), 3752. https://doi.org/10.3390/ijms22073752