
Effect of Central Corticotropin-Releasing Factor on Hepatic Lipid Metabolism and Inflammation-Related Gene Expression in Rats

Abstract
:1. Introduction
2. Results
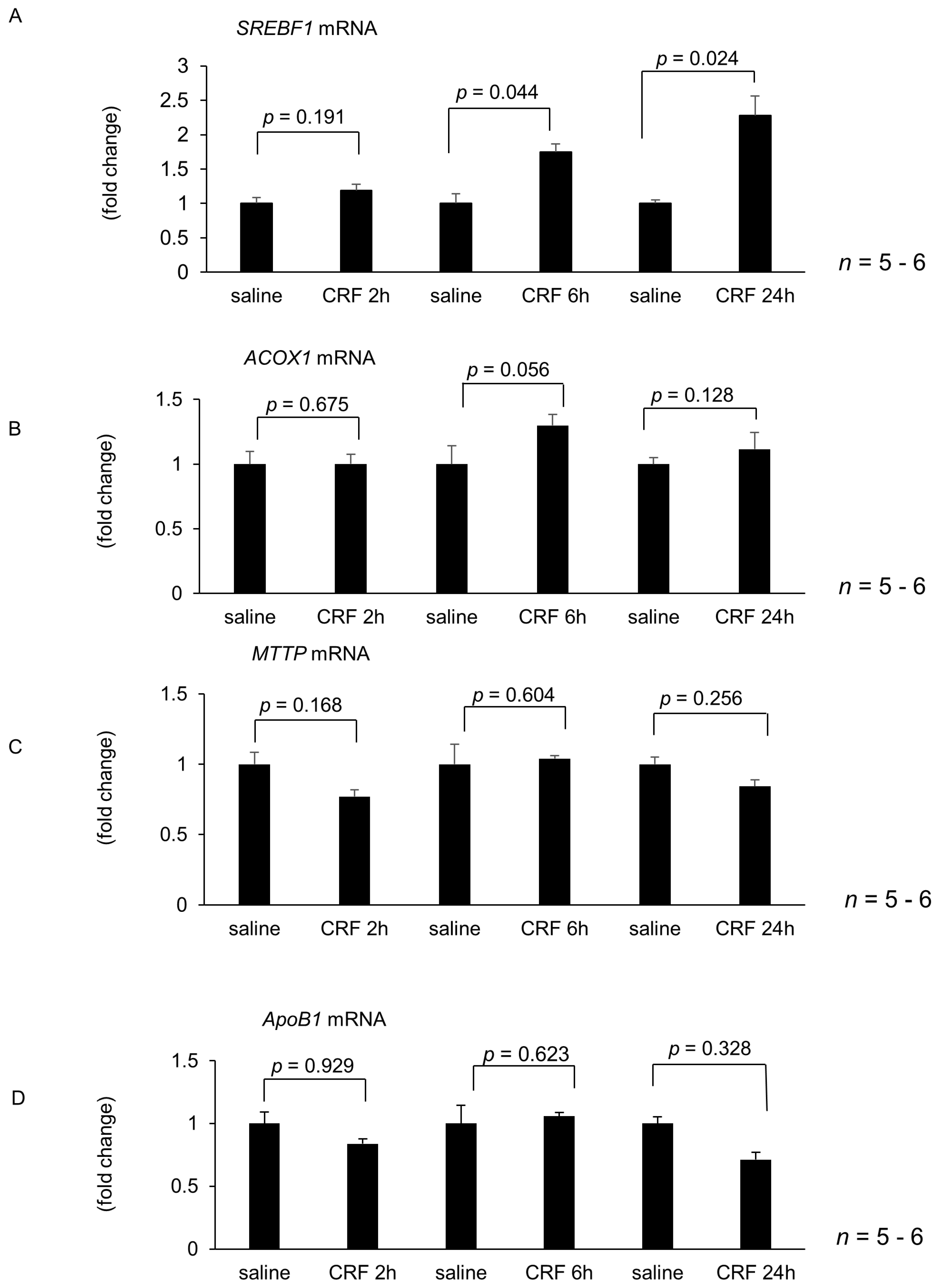
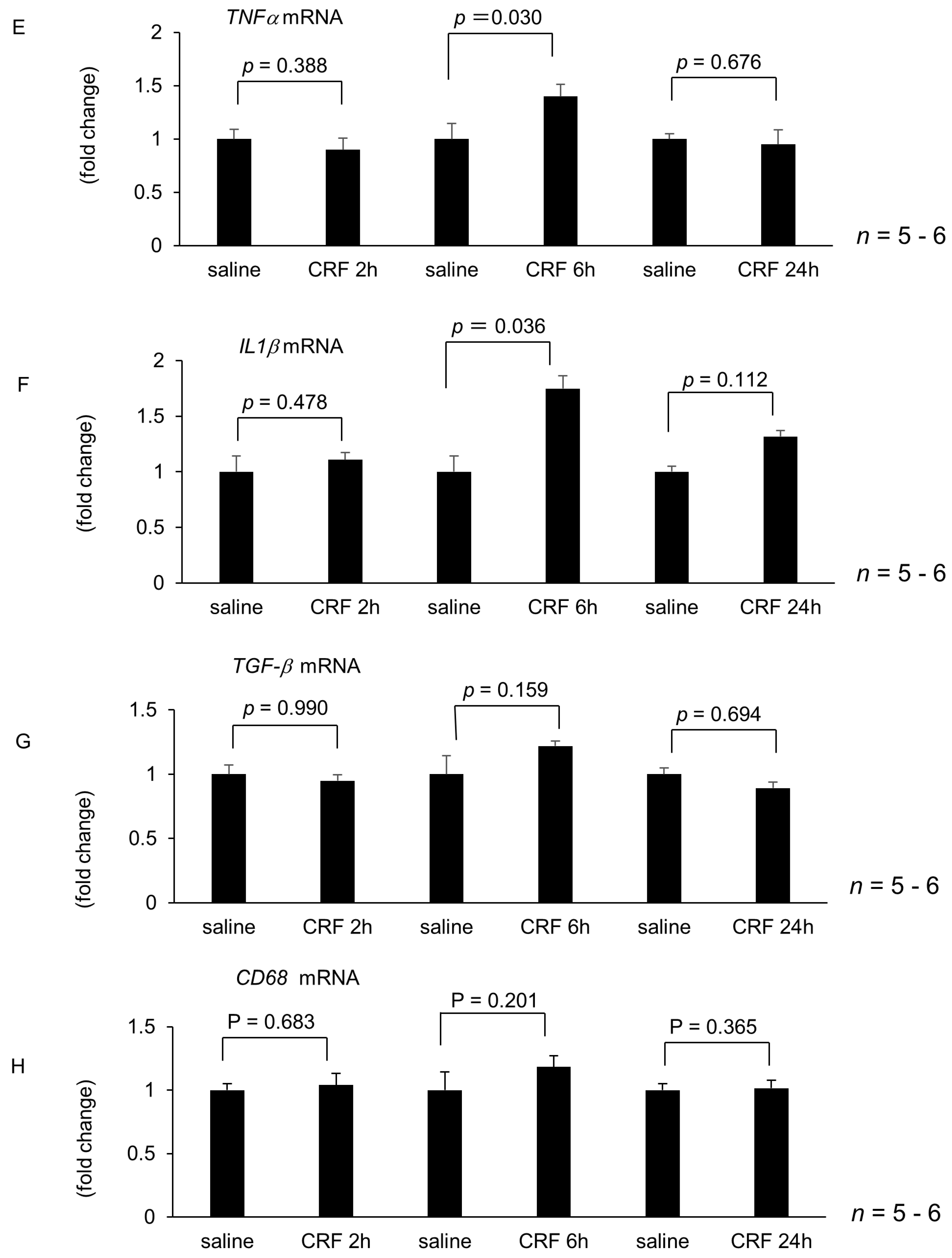
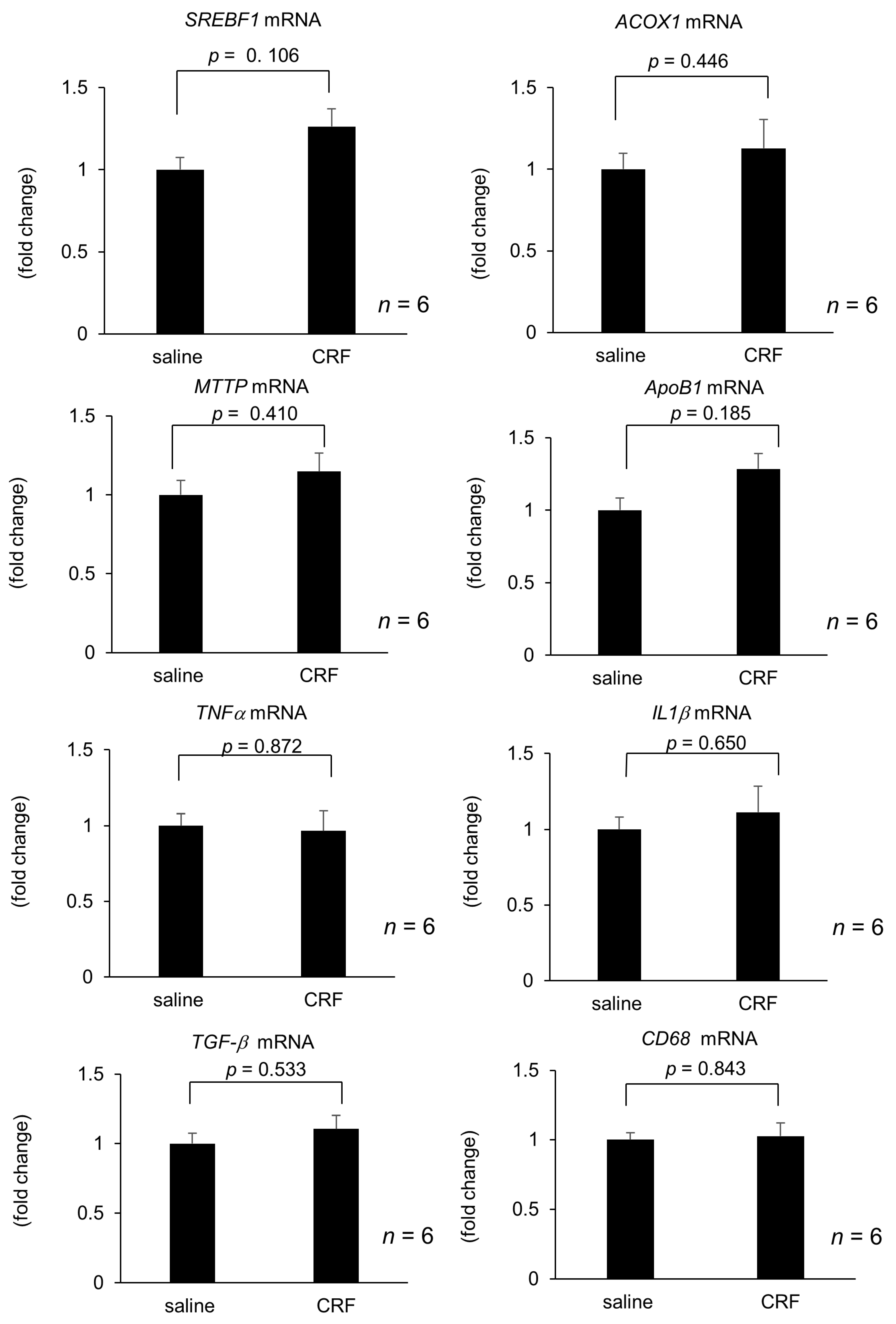
2.1. Effect of Intracisternal CRF Injection on Hepatic Lipid Metabolism and Inflammation-Related Gene Expression
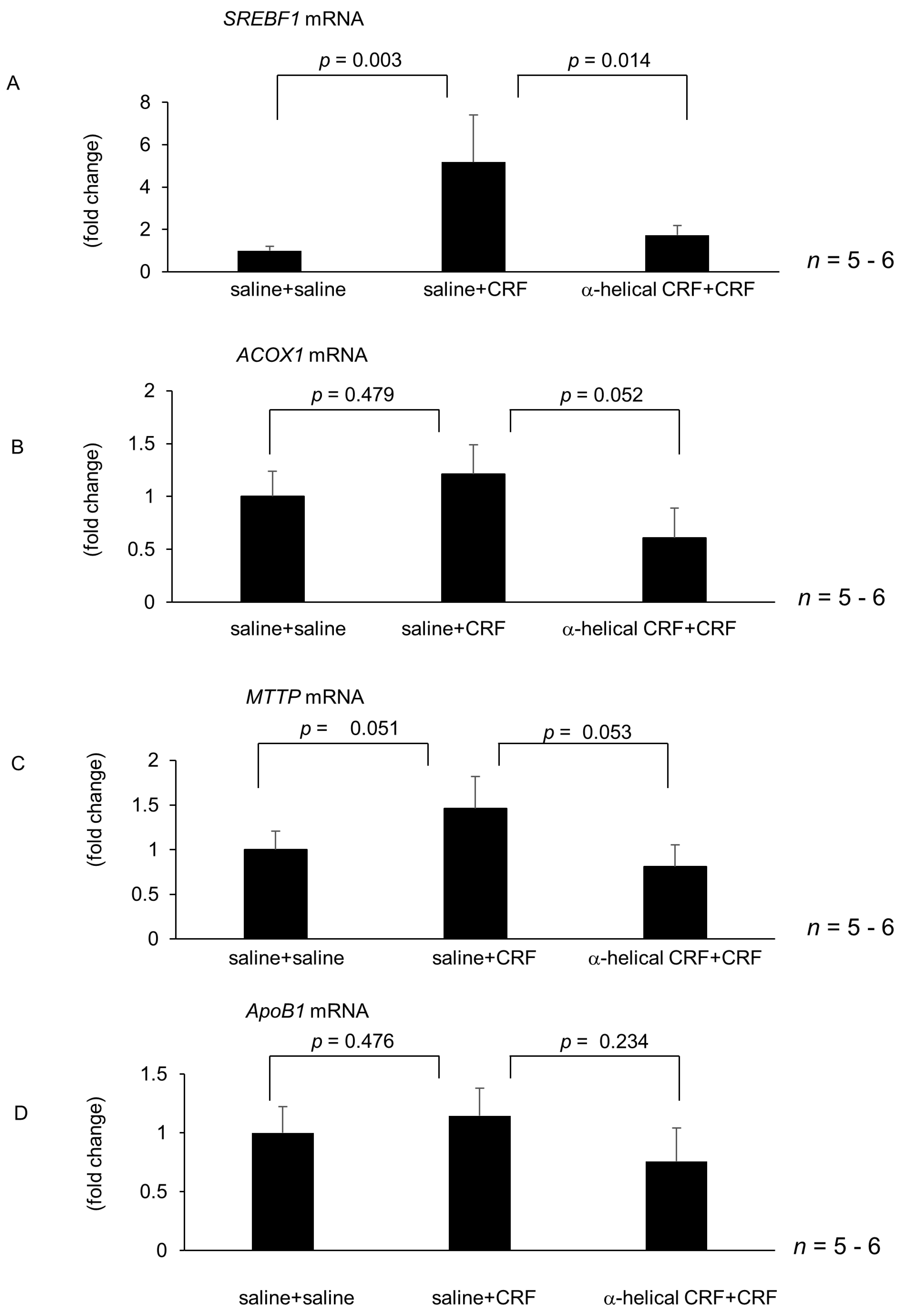
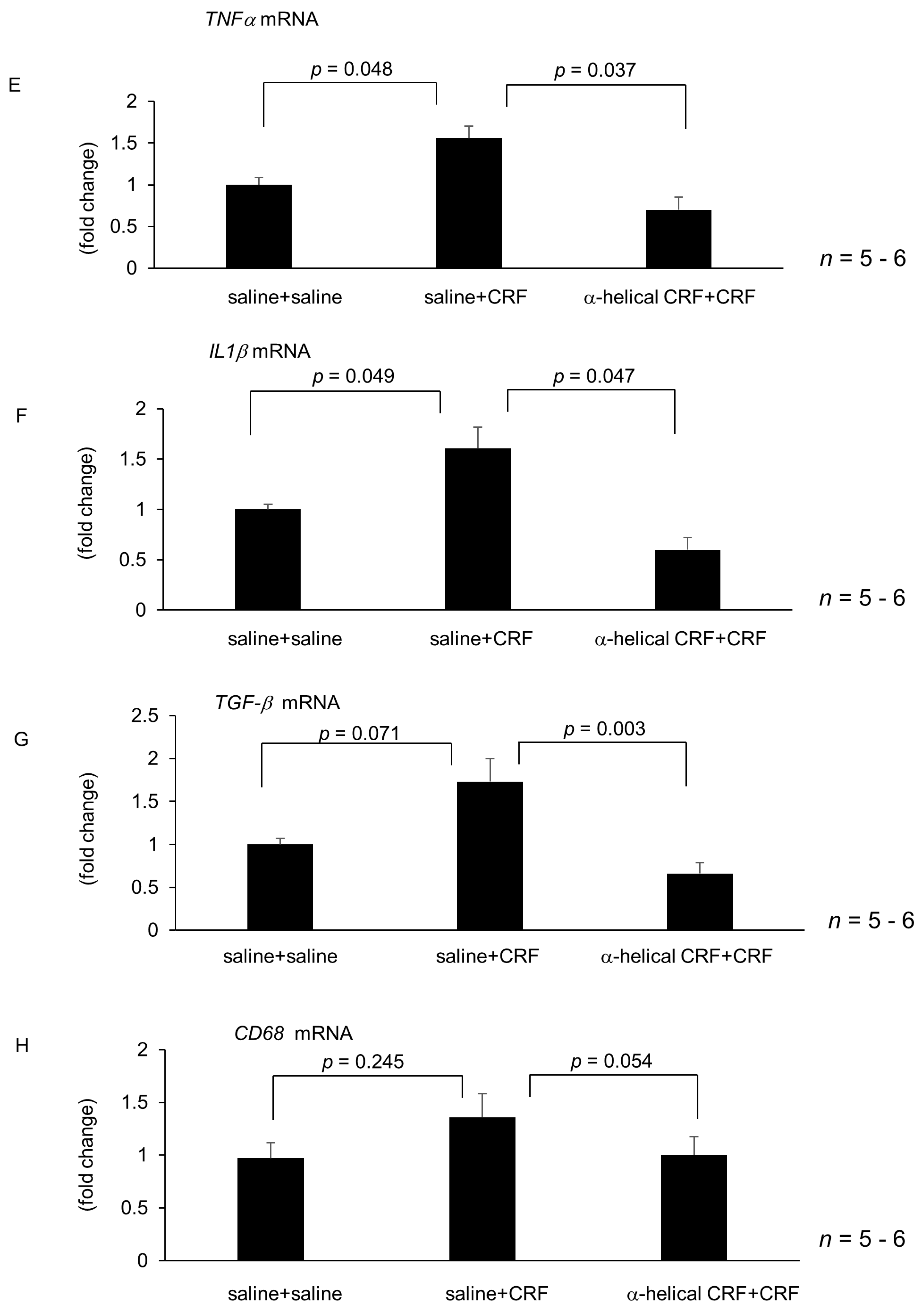
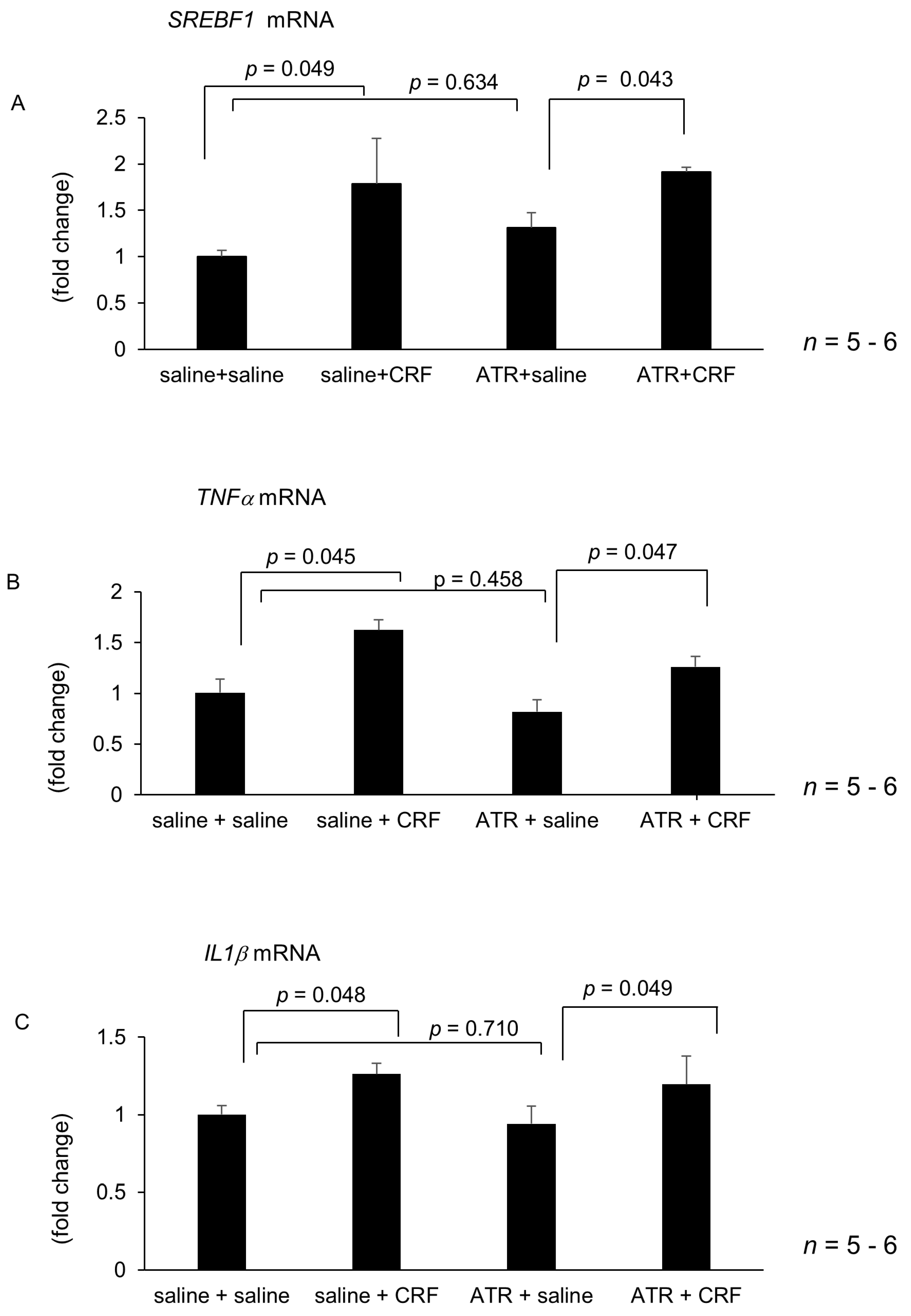
2.2. Effect of Central CRF Antagonist on CRF-Induced Modulation of Hepatic Lipid Metabolism and Inflammation-Related Gene Expression
2.3. Effect of Intravenous CRF Injection on Hepatic Lipid Metabolism and Inflammation-Related Gene Expression
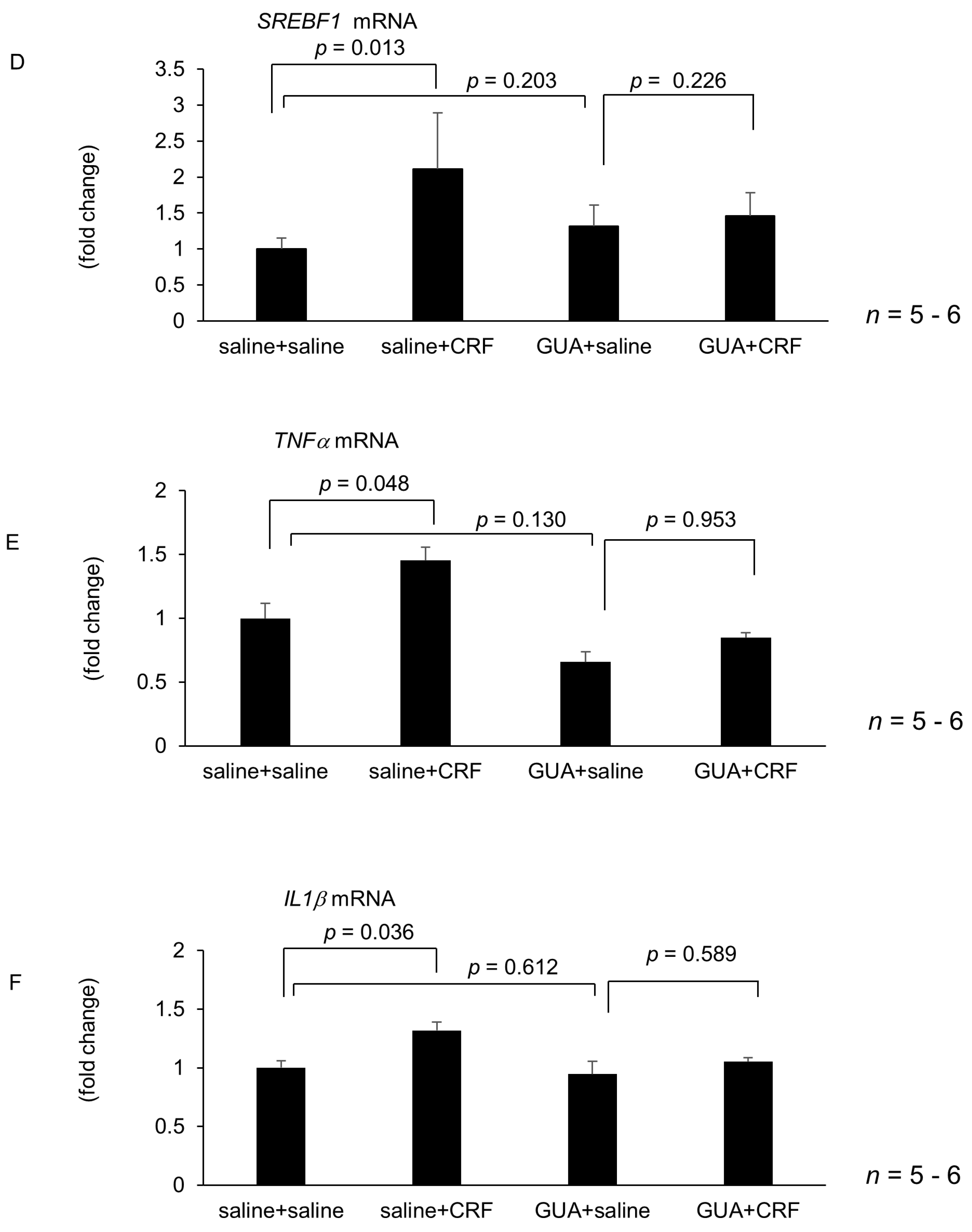
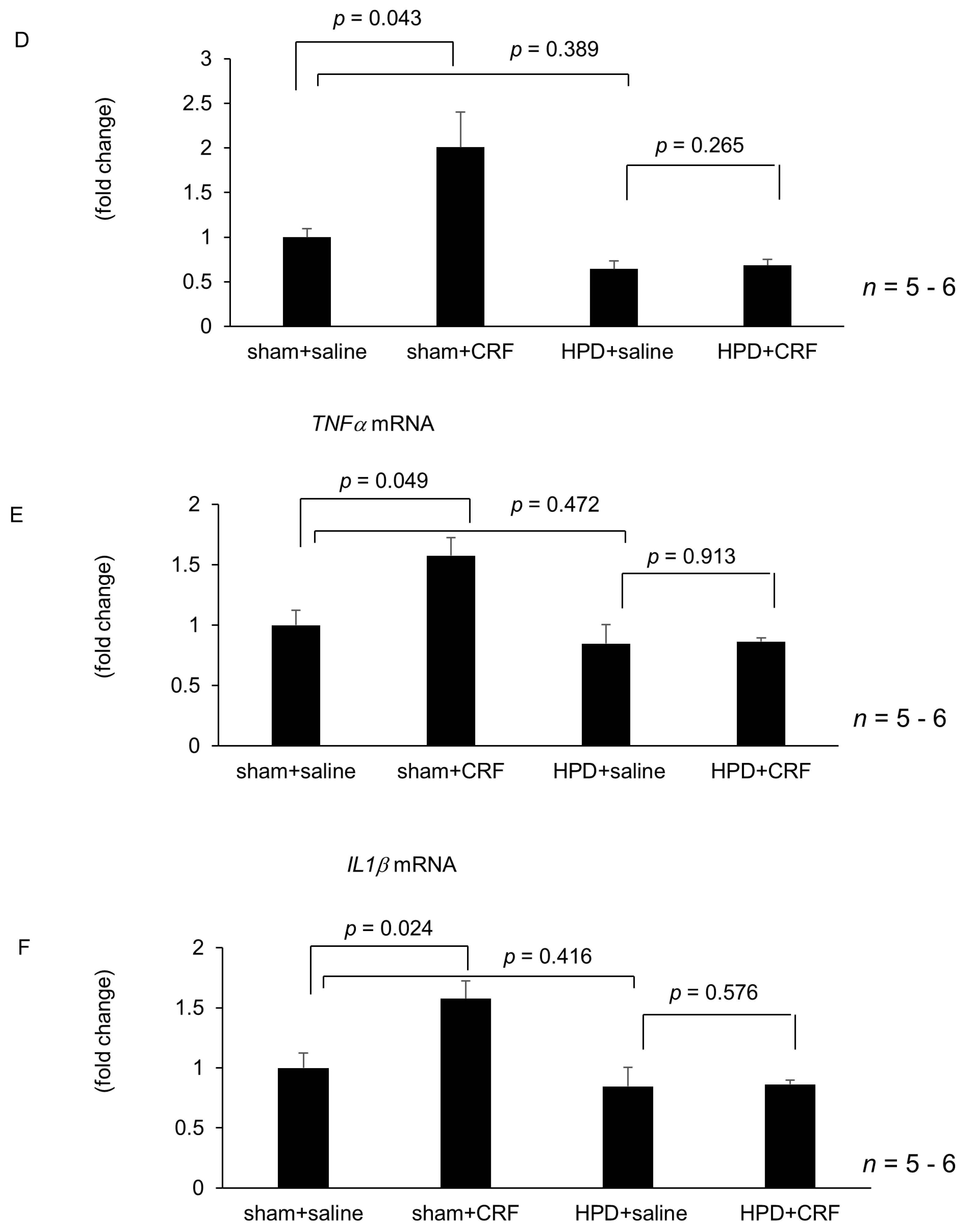
2.4. Effect of Atropine, Guanethidine, Hepatic Branch Vagotomy, and Hepatic Plexus Denervation on Intracisternal CRF-Medicated Hepatic Gene Expressions
3. Discussion
4. Materials and Methods
4.1. Substances and Treatments
4.2. Animal Model and Experimental Design
4.3. Serum Biochemical Measurements
4.4. Real-Time Polymerase Chain Reaction (PCR)
4.5. Effect of Central CRF Antagonist on CRF-Induced Modulation of Hepatic Gene Expressions
4.6. Effect of Atropine, Guanethidine, Hepatic BRANCH Vagotomy and Hepatic Plexus Denervation on CRF-Induced Modulation of Gene Expression
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Lautt, W.W. Afferent and efferent neural roles in liver function. Prog. Neurobiol. 1983, 21, 323–348. [Google Scholar] [CrossRef]
- Shimazu, T.; Matsushita, H.; Ishikawa, K. Cholinergic stimulation of the rat hypothalamus: Effects of liver glycogen synthesis. Science 1976, 194, 535–536. [Google Scholar] [CrossRef]
- Brown, M.; Rivier, J.; Vale, W. Somatostatin: Central nervous system actions on glucoregulation. Endocrinology 1979, 104, 1709–1715. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.G.; Hacker, R.R.; Burton, J.H.; Veira, D.M. Response of bovine serum prolactin and growth hormone to duodenal, abomasal, and oral administration of thyrotropin-releasing hormone. J. Dairy Sci. 1977, 60, 1624–1628. [Google Scholar] [CrossRef]
- Im, E.; Rhee, S.H.; Park, Y.S.; Fiocchi, C.; Tache, Y.; Pothoulakis, C. Corticotropin-releasing hormone family of peptides regulates intestinal angiogenesis. Gastroenterology 2010, 138, 2457–2467.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tache, Y.; Monnikes, H.; Bonaz, B.; Rivier, J. Role of CRF in stress-related alterations of gastric and colonic motor function. Ann. N. Y. Acad. Sci. 1993, 697, 233–243. [Google Scholar] [CrossRef]
- Gardemann, A.; Puschel, G.P.; Jungermann, K. Nervous control of liver metabolism and hemodynamics. Eur. J. Biochem. 1992, 207, 399–411. [Google Scholar] [CrossRef]
- Iwai, M.; Shimazu, T. Effects of ventromedial and lateral hypothalamic stimulation on chemically-induced liver injury in rats. Life Sci. 1988, 42, 1833–1840. [Google Scholar] [CrossRef]
- Rogers, R.C.; Hermann, G.E. Central connections of the hepatic branch of the vagus nerve: A horseradish peroxidase histochemical study. J. Auton. Nerv. Syst. 1983, 7, 165–174. [Google Scholar] [CrossRef]
- Koo, A.; Liang, I.Y. Microvascular filling pattern in rat liver sinusoids during vagal stimulation. J. Physiol. 1979, 295, 191–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Yoneda, M.; Takamoto, S.; Nakade, Y.; Yokohama, S.; Tamori, K.; Aso, K.; Matui, T.; Sato, Y.; Aoshima, M.; et al. Effect of ursodeoxycholic acid on autoimmune-associated chronic hepatitis C. J. Gastroenterol. Hepatol. 1999, 14, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, M.; Nakamura, K.; Nakade, Y.; Tamano, M.; Kono, T.; Watanobe, H.; Shimada, T.; Hiraishi, H.; Terano, A. Effect of central corticotropin releasing factor on hepatic circulation in rats: The role of the CRF2 receptor in the brain. Gut 2005, 54, 282–288. [Google Scholar] [CrossRef] [Green Version]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Ekstedt, M.; Hagstrom, H.; Nasr, P.; Fredrikson, M.; Stal, P.; Kechagias, S.; Hultcrantz, R. Fibrosis stage is the strongest predictor for disease-specific mortality in NAFLD after up to 33 years of follow-up. Hepatology 2015, 61, 1547–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angulo, P.; Kleiner, D.E.; Dam-Larsen, S.; Adams, L.A.; Bjornsson, E.S.; Charatcharoenwitthaya, P.; Mills, P.R.; Keach, J.C.; Lafferty, H.D.; Stahler, A.; et al. Liver Fibrosis, but No Other Histologic Features, Is Associated with Long-term Outcomes of Patients with Nonalcoholic Fatty Liver Disease. Gastroenterology 2015, 149, 389–397.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Rinella, M.E.; Elias, M.S.; Smolak, R.R.; Fu, T.; Borensztajn, J.; Green, R.M. Mechanisms of hepatic steatosis in mice fed a lipogenic methionine choline-deficient diet. J. Lipid Res. 2008, 49, 1068–1076. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Saxena, N.K.; Lin, S.; Gupta, N.A.; Anania, F.A. Exendin-4, a glucagon-like protein-1 (GLP-1) receptor agonist, reverses hepatic steatosis in ob/ob mice. Hepatology 2006, 43, 173–181. [Google Scholar] [CrossRef]
- Charlton, M.; Sreekumar, R.; Rasmussen, D.; Lindor, K.; Nair, K.S. Apolipoprotein synthesis in nonalcoholic steatohepatitis. Hepatology 2002, 35, 898–904. [Google Scholar] [CrossRef]
- Gadd, V.L.; Skoien, R.; Powell, E.E.; Fagan, K.J.; Winterford, C.; Horsfall, L.; Irvine, K.; Clouston, A.D. The portal inflammatory infiltrate and ductular reaction in human nonalcoholic fatty liver disease. Hepatology 2014, 59, 1393–1405. [Google Scholar] [CrossRef]
- Yamada, T.; Oka, Y.; Katagiri, H. Inter-organ metabolic communication involved in energy homeostasis: Potential therapeutic targets for obesity and metabolic syndrome. Pharmacol. Ther. 2008, 117, 188–198. [Google Scholar] [CrossRef]
- Clark, J.T.; Kalra, P.S.; Kalra, S.P. Neuropeptide Y stimulates feeding but inhibits sexual behavior in rats. Endocrinology 1985, 117, 2435–2442. [Google Scholar] [CrossRef]
- Billington, C.J.; Briggs, J.E.; Harker, S.; Grace, M.; Levine, A.S. Neuropeptide Y in hypothalamic paraventricular nucleus: A center coordinating energy metabolism. Am. J. Physiol. 1994, 266, R1765–R1770. [Google Scholar] [CrossRef] [PubMed]
- Bruinstroop, E.; Pei, L.; Ackermans, M.T.; Foppen, E.; Borgers, A.J.; Kwakkel, J.; Alkemade, A.; Fliers, E.; Kalsbeek, A. Hypothalamic neuropeptide Y (NPY) controls hepatic VLDL-triglyceride secretion in rats via the sympathetic nervous system. Diabetes 2012, 61, 1043–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krahn, D.D.; Gosnell, B.A.; Grace, M.; Levine, A.S. CRF antagonist partially reverses CRF- and stress-induced effects on feeding. Brain Res. Bull. 1986, 17, 285–289. [Google Scholar] [CrossRef]
- Barquist, E.; Zinner, M.; Rivier, J.; Tache, Y. Abdominal surgery-induced delayed gastric emptying in rats: Role of CRF and sensory neurons. Am. J. Physiol. 1992, 262, G616–G620. [Google Scholar] [CrossRef] [PubMed]
- Lenz, H.J.; Messmer, B.; Zimmerman, F.G. Noradrenergic inhibition of canine gallbladder contraction and murine pancreatic secretion during stress by corticotropin-releasing factor. J. Clin. Investig. 1992, 89, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Gerber, J.G.; Freed, C.R.; Nies, A.S. Antihypertensive pharmacology. West. J. Med. 1980, 132, 430–439. [Google Scholar] [PubMed]
- Fukudo, S.; Suzuki, J.; Tanaka, Y.; Iwahashi, S.; Nomura, T. Impact of stress on alcoholic liver injury; a histopathological study. J. Psychosom. Res. 1989, 33, 515–521. [Google Scholar] [CrossRef]
- Hsu, C.T. The role of the sympathetic nervous system in promoting liver cirrhosis induced by carbon tetrachloride, using the essential hypertensive animal (SHR). J. Auton. Nerv. Syst. 1992, 37, 163–173. [Google Scholar] [CrossRef]
- Hsu, C.T.; Schichijo, K.; Ito, M.; Sekine, I. The effect of chemical sympathectomy on acute liver injury induced by carbon tetrachloride in spontaneously hypertensive rats. J. Auton. Nerv. Syst. 1993, 43, 91–96. [Google Scholar] [CrossRef]
- Haas, D.A.; George, S.R. Single or repeated mild stress increases synthesis and release of hypothalamic corticotropin-releasing factor. Brain Res. 1988, 461, 230–237. [Google Scholar] [CrossRef]
- Kalin, N.H.; Takahashi, L.K.; Chen, F.L. Restraint stress increases corticotropin-releasing hormone mRNA content in the amygdala and paraventricular nucleus. Brain Res. 1994, 656, 182–186. [Google Scholar] [CrossRef]
- Lenz, H.J.; Raedler, A.; Greten, H.; Vale, W.W.; Rivier, J.E. Stress-induced gastrointestinal secretory and motor responses in rats are mediated by endogenous corticotropin-releasing factor. Gastroenterology 1988, 95, 1510–1517. [Google Scholar] [CrossRef]
- Nakade, Y.; Yoneda, M.; Nakamura, K.; Makino, I.; Terano, A. Involvement of endogenous CRF in carbon tetrachloride-induced acute liver injury in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, R1782–R1788. [Google Scholar] [CrossRef] [Green Version]
- Hurr, C.; Simonyan, H.; Morgan, D.A.; Rahmouni, K.; Young, C.N. Liver sympathetic denervation reverses obesity-induced hepatic steatosis. J. Physiol. 2019, 597, 4565–4580. [Google Scholar] [CrossRef] [PubMed]
- De Souza, E.B.; Insel, T.R.; Perrin, M.H.; Rivier, J.; Vale, W.W.; Kuhar, M.J. Corticotropin-releasing factor receptors are widely distributed within the rat central nervous system: An autoradiographic study. J. Neurosci. 1985, 5, 3189–3203. [Google Scholar] [CrossRef] [Green Version]
- Chalmers, D.T.; Lovenberg, T.W.; Grigoriadis, D.E.; Behan, D.P.; De Souza, E.B. Corticotrophin-releasing factor receptors: From molecular biology to drug design. Trends Pharmacol. Sci. 1996, 17, 166–172. [Google Scholar] [CrossRef]
- Lovenberg, T.W.; Liaw, C.W.; Grigoriadis, D.E.; Clevenger, W.; Chalmers, D.T.; De Souza, E.B.; Oltersdorf, T. Cloning and characterization of a functionally distinct corticotropin-releasing factor receptor subtype from rat brain. Proc. Natl. Acad. Sci. USA 1995, 92, 836–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrin, M.; Donaldson, C.; Chen, R.; Blount, A.; Berggren, T.; Bilezikjian, L.; Sawchenko, P.; Vale, W. Identification of a second corticotropin-releasing factor receptor gene and characterization of a cDNA expressed in heart. Proc. Natl. Acad. Sci. USA 1995, 92, 2969–2973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stafford, J.M.; Yu, F.; Printz, R.; Hasty, A.H.; Swift, L.L.; Niswender, K.D. Central nervous system neuropeptide Y signaling modulates VLDL triglyceride secretion. Diabetes 2008, 57, 1482–1490. [Google Scholar] [CrossRef] [Green Version]
- Nakade, Y.; Tsuchida, D.; Fukuda, H.; Iwa, M.; Pappas, T.N.; Takahashi, T. Restraint stress delays solid gastric emptying via a central CRF and peripheral sympathetic neuron in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R427–R432. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.L.; Yamano, M.; Shiotani, Y.; Emson, P.C.; Smith, A.D.; Powell, J.F.; Tohyama, M. An arcuato-paraventricular and -dorsomedial hypothalamic neuropeptide Y-containing system which lacks noradrenaline in the rat. Brain Res. 1985, 331, 172–175. [Google Scholar] [CrossRef]
- Lambert, P.D.; Wilding, J.P.; Turton, M.D.; Ghatei, M.A.; Bloom, S.R. Effect of food deprivation and streptozotocin-induced diabetes on hypothalamic neuropeptide Y release as measured by a radioimmunoassay-linked microdialysis procedure. Brain Res. 1994, 656, 135–140. [Google Scholar] [CrossRef]
- Vinuela, M.C.; Larsen, P.J. Identification of NPY-induced c-Fos expression in hypothalamic neurones projecting to the dorsal vagal complex and the lower thoracic spinal cord. J. Comp. Neurol. 2001, 438, 286–299. [Google Scholar] [CrossRef] [PubMed]
- La Fleur, S.E.; Kalsbeek, A.; Wortel, J.; Buijs, R.M. Polysynaptic neural pathways between the hypothalamus, including the suprachiasmatic nucleus, and the liver. Brain Res. 2000, 871, 50–56. [Google Scholar] [CrossRef]
- Stanley, S.; Pinto, S.; Segal, J.; Perez, C.A.; Viale, A.; DeFalco, J.; Cai, X.; Heisler, L.K.; Friedman, J.M. Identification of neuronal subpopulations that project from hypothalamus to both liver and adipose tissue polysynaptically. Proc. Natl. Acad. Sci. USA 2010, 107, 7024–7029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamaki, Y.; Nakade, Y.; Yamauchi, T.; Makino, Y.; Yokohama, S.; Okada, M.; Aso, K.; Kanamori, H.; Ohashi, T.; Sato, K.; et al. Angiotensin II type 1 receptor antagonist prevents hepatic carcinoma in rats with nonalcoholic steatohepatitis. J. Gastroenterol. 2013, 48, 491–503. [Google Scholar] [CrossRef]
- Tanaka, K.; Ohkawa, S.; Nishino, T.; Niijima, A.; Inoue, S. Role of the hepatic branch of the vagus nerve in liver regeneration in rats. Am. J. Physiol. 1987, 253, G439–G444. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | n | 0 h | 2 h | 6 h | 24 h |
|---|---|---|---|---|---|
| CRF | 6 | 0.42 ± 0.02 | 0.69 ± 0.03 | 0.61 ± 0.05 | 0.60 ± 0.05 |
| saline | 6 | 0.38 ± 0.05 | 0.59 ± 0.09 | 0.71 ± 0.07 | 0.50 ± 0.06 |
| Group | n | 0 h | 2 h | 6 h | 24 h |
|---|---|---|---|---|---|
| CRF | 6 | 152 ± 14 | 181 ± 26 | 203 ± 21 | 213 ± 9 * |
| saline | 6 | 148 ± 11 | 151 ± 21 | 159 ± 16 | 157 ± 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakade, Y.; Kitano, R.; Yamauchi, T.; Kimoto, S.; Sakamoto, K.; Inoue, T.; Kobayashi, Y.; Ohashi, T.; Sumida, Y.; Ito, K.; et al. Effect of Central Corticotropin-Releasing Factor on Hepatic Lipid Metabolism and Inflammation-Related Gene Expression in Rats. Int. J. Mol. Sci. 2021, 22, 3940. https://doi.org/10.3390/ijms22083940
Nakade Y, Kitano R, Yamauchi T, Kimoto S, Sakamoto K, Inoue T, Kobayashi Y, Ohashi T, Sumida Y, Ito K, et al. Effect of Central Corticotropin-Releasing Factor on Hepatic Lipid Metabolism and Inflammation-Related Gene Expression in Rats. International Journal of Molecular Sciences. 2021; 22(8):3940. https://doi.org/10.3390/ijms22083940
Chicago/Turabian StyleNakade, Yukiomi, Rena Kitano, Taeko Yamauchi, Satoshi Kimoto, Kazumasa Sakamoto, Tadahisa Inoue, Yuji Kobayashi, Tomohiko Ohashi, Yoshio Sumida, Kiyoaki Ito, and et al. 2021. "Effect of Central Corticotropin-Releasing Factor on Hepatic Lipid Metabolism and Inflammation-Related Gene Expression in Rats" International Journal of Molecular Sciences 22, no. 8: 3940. https://doi.org/10.3390/ijms22083940
APA StyleNakade, Y., Kitano, R., Yamauchi, T., Kimoto, S., Sakamoto, K., Inoue, T., Kobayashi, Y., Ohashi, T., Sumida, Y., Ito, K., & Yoneda, M. (2021). Effect of Central Corticotropin-Releasing Factor on Hepatic Lipid Metabolism and Inflammation-Related Gene Expression in Rats. International Journal of Molecular Sciences, 22(8), 3940. https://doi.org/10.3390/ijms22083940