
In Vitro and Vivo Identification, Metabolism and Action of Xenoestrogens: An Overview



Abstract
:1. Introduction
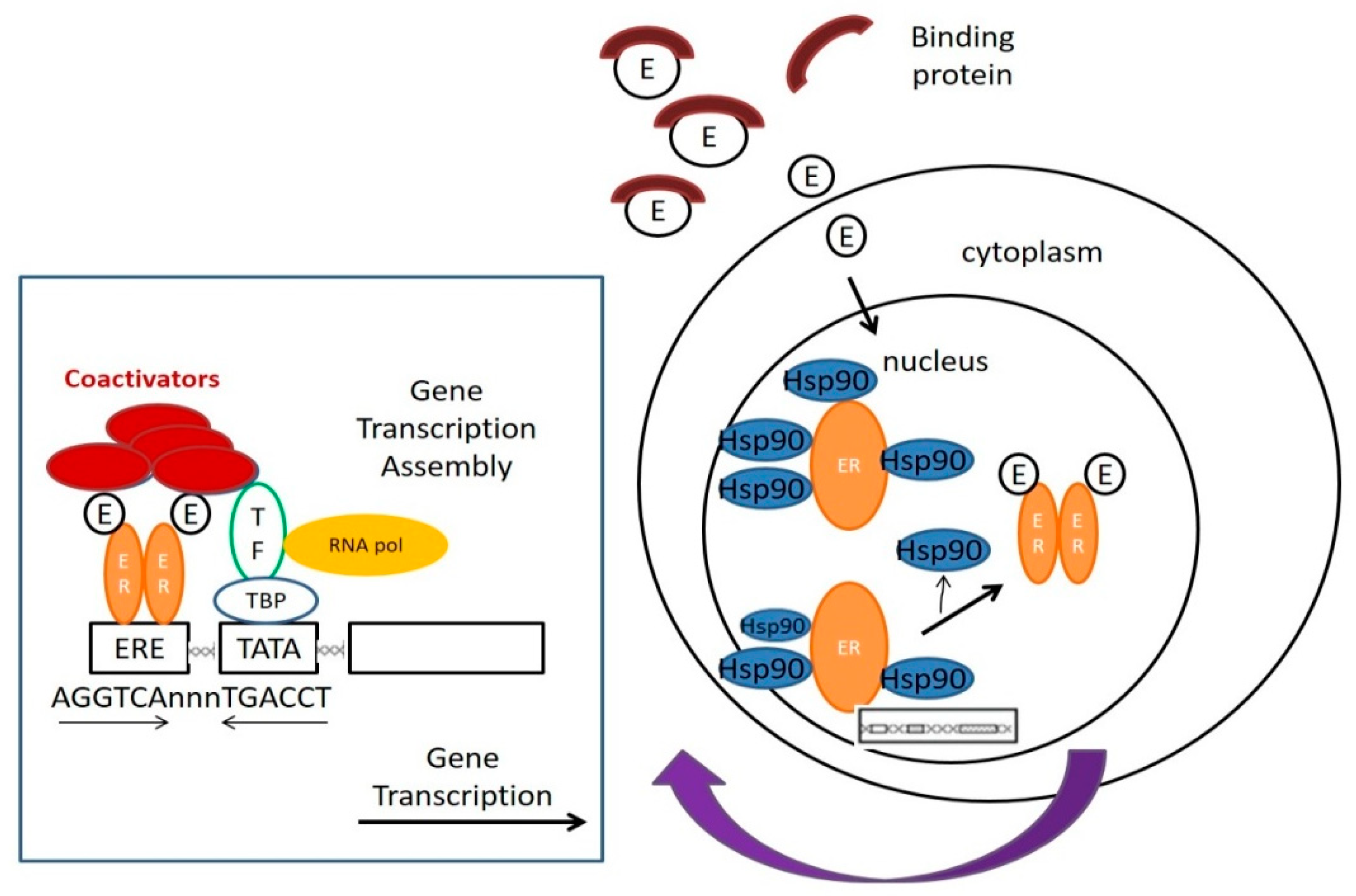
- (1.)
- Estradiol, a type of estrogen (E), is trapped over by carrier protein in the serum. After the estrogen is released from the blood by the carrier protein, it can pass through the cell membrane without disturbance and enter the cell;
- (2.)
- Located within the nucleus, before activation, the ERs will be bound to various receptor-associated proteins, such as heat-shock proteins (Hsp90);
- (3.)
- Estrogen will be bound to the ERs, replacing the receptor-associated proteins;
- (4.)
- ER dimer is bound to its corresponding DNA-binding domains, and the sequence is named estrogen-responsive element (ERE);
- (5.)
- Under the cooperation of assembled substances, including multiple transcription factors (TF), the RNA polymerase (RNA Pol) and other proteins, transcription can be started. The relevant mRNA sequence emerges as ER target genes transcription is in the process;
- (6.)
- Co-activators, including CBP/p300 and SRC-1, are all the linking brackets to ER dimer and could play an effect on DNA transcription;
- (7.)
- After DNA is transcribed into mRNA, RNA should be translated to produce protein to complete gene expression [10].
2. Identification and Detection of Xenoestrogens
2.1. E-SCREEN
2.2. Ligand-Binding
2.3. ER-Binding to ERE
2.4. Mammalian Two-Hybrid Assays
2.5. Analysis of Gene Expression
2.6. Analysis of Enzyme Activity
2.7. Analysis of Steroido-Genesis
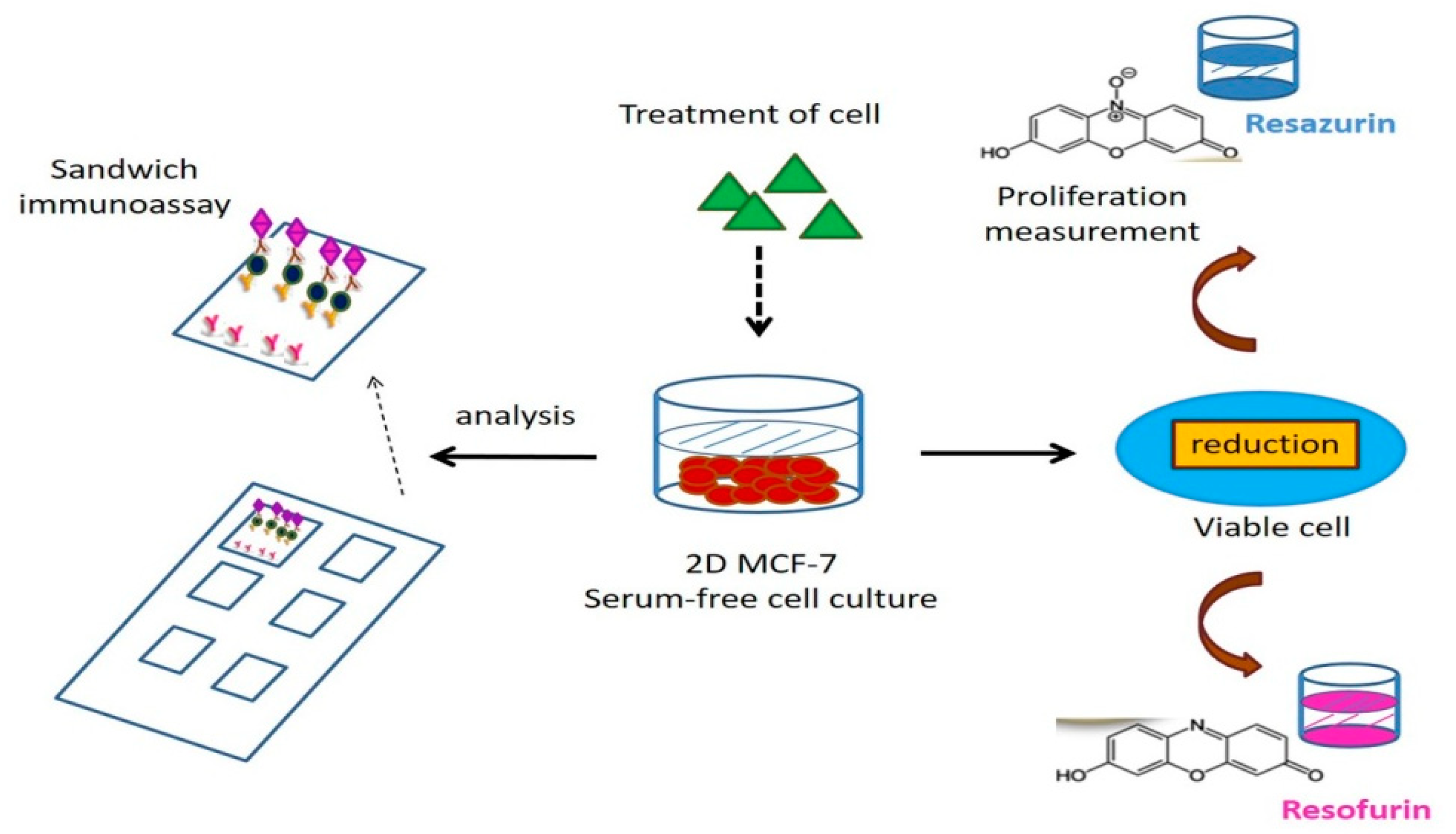
2.8. The Fluorescence-Based Multi-Analyte Chip Platform
2.9. Transcriptional Activation Assays
2.10. Triple Functional Small-Molecule–Protein Conjugate-Mediated Optical Biosensor
2.11. Estrogen Receptor Recombinant Yeast Screening (YES) Assay
3. Metabolism and Action of Xenoestrogens
3.1. The Metabolism of Xenoestrogens
3.2. The Actions of Xenoestrogens
3.2.1. Selective Estrogen Receptor Modulators (SERMs) and Aromatase Interferer
3.2.2. Polycyclic Aromatic Hydrocarbons (PAHs)
3.2.3. DDT [2,2-Bis(p-chlorophenyl)-1,1,1-trichloroethan] and Its Metabolite DDE
3.2.4. Polychlorinated Biphenyls (PCBs)
3.2.5. The Relationship between EDCs and Diseases
3.2.6. Bisphenol A (BPA)
3.2.7. Heavy Metals: Cadmium (Cd) and Arsenic (As)
3.2.8. Phytoestrogens
3.2.9. Diethylstilbestrol (DES) and 17-α-ethinylestradiol
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Olea, N.; Pazos, P.; Exposito, J. Inadvertent exposure to xenoestrogens. Eur. J. Cancer Prev. 1998, 7 (Suppl. 1), S17–S23. [Google Scholar] [CrossRef]
- Olea, N.; Pulgar, R.; Pérez, P.; Olea-Serrano, F.; Rivas, A.; Novillo-Fertrell, A.; Pedraza, V.; Soto, A.M.; Sonnenschein, C. Estrogenicity of resin-based composites and sealants used in dentistry. Environ. Health Perspect. 1996, 104, 298–305. [Google Scholar] [CrossRef]
- Viñas, R.; Jeng, Y.J.; Watson, C.S. Non-Genomic Effects of Xenoestrogen Mixtures. Int. J. Environ. Res. Public Health 2012, 9, 2694–2714. [Google Scholar] [CrossRef] [PubMed]
- Bronowicka-Kłys, D.E.; Lianeri, M.; Jagodziński, P.P. The role and impact of estrogens and xenoestrogen on the development of cervical cancer. Biomed. Pharmacother. 2016, 84, 1945–1953. [Google Scholar] [CrossRef] [PubMed]
- Singleton, D.W.; Khan, S.A. Xenoestrogen exposure and mechanisms of endocrine disruption. Front. Biosci. 2003, 8, S110–S118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndebele, K.; Graham, B.; Tchounwou, P.B. Estrogenic Activity of Coumestrol, DDT, and TCDD in Human Cervical Cancer Cells. Int. J. Environ. Res. Public Health 2010, 7, 2045–2056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wozniak, M.; Murias, M. Xenoestrogens: Endocrine disrupting compounds. Ginekol. Polska 2008, 79, 785–790. [Google Scholar]
- Ewa Forma, A.S.; Krześlak, A. Selected xenoestrogens and their impact on human health. Folia Med. Lodz. 2013, 40, 79–97. [Google Scholar]
- Mueller, S.O. Xenoestrogens: Mechanisms of action and detection methods. Anal. Bioanal. Chem. 2004, 378, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.O.; Korach, K.S. Mechanisms of Estrogen Receptor-Mediated Agonistic and Antagonistic Effects. In The Handbook of Environmental Chemistry; Springer: Berlin/Heidelberg, Germany, 2001. [Google Scholar]
- Hall, J.M.; Couse, J.F.; Korach, K.S. The Multifaceted Mechanisms of Estradiol and Estrogen Receptor Signaling. J. Biol. Chem. 2001, 276, 36869–36872. [Google Scholar] [CrossRef] [Green Version]
- Karoutsou, E.; Karoutsos, P.; Karoutsos, D. Adverse Effects on Female Human Reproductive Health from Exposure to Endocrine Disruptors: Focus on Endometrial Lesions. J. Clin. Epigenet. 2016, 2. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, S.; Russo, J. Estrogen and Xenoestrogens in Breast Cancer. Toxicol. Pathol. 2010, 38, 110–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, A.; Pirzada, M.; Jahan, S.; Ullah, H.; Turi, N.; Ullah, W.; Siddiqui, M.F.; Zakria, M.; Lodhi, K.Z.; Khan, M.M. Impact of low-dose chronic exposure to bisphenol A and its analogue bisphenol B, bisphenol F and bisphenol S on hypothalamo-pituitary-testicular activities in adult rats: A focus on the possible hormonal mode of action. Food Chem. Toxicol. 2018, 121, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Pelch, K.E.; Carleton, S.M.; Phillips, C.L.; Nagel, S.C. Developmental Exposure to Xenoestrogens at Low Doses Alters Femur Length and Tensile Strength in Adult Mice1. Biol. Reprod. 2012, 86, 69. [Google Scholar] [CrossRef]
- Korach, K.S. Surprising places of estrogenic activity. Endocrinology 1993, 132, 2277–2278. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.R. Prenatal Diethylstilbestrol Exposure and Risk of Breast Cancer. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1509–1514. [Google Scholar] [CrossRef] [Green Version]
- Toppari, J.; Larsen, J.C.; Christiansen, P.; Giwercman, A.; Grandjean, P.; Guillette, L.J., Jr.; Jégou, B.; Jensen, T.K.; Jouannet, P.; Keiding, N. Male reproductive health and environmental xenoestrogens. Environ. Health Perspect. 1996, 104 (Suppl. 4), 741–803. [Google Scholar]
- Shanle, E.K.; Xu, W. Endocrine Disrupting Chemicals Targeting Estrogen Receptor Signaling: Identification and Mechanisms of Action. Chem. Res. Toxicol. 2011, 24, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Barnes, S.; Peterson, T.G. Biochemical Targets of the Isoflavone Genistein in Tumor Cell Lines. Exp. Biol. Med. 1995, 208, 103–108. [Google Scholar] [CrossRef]
- Soto, A.M.; Sonnenschein, C. Environmental causes of cancer: Endocrine disruptors as carcinogens. Nat. Rev. Endocrinol. 2010, 6, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.A.; Hissong, B.D.; Verthelyi, D.; Donner, K.; Becker, K.; Karpuzoglu-Sahin, E. Gender and risk of autoimmune diseases: Possible role of estrogenic compounds. Environ. Health Perspect. 1999, 107 (Suppl. 5), 681–686. [Google Scholar]
- Marselos, M.; Tomatis, L. Diethylstilboestrol: II, pharmacology, toxicology and carcinogenicity in experimental animals. Eur. J. Cancer 1992, 29A, 149–155. [Google Scholar] [CrossRef]
- Newbold, R.R.; Bullock, B.C.; A McLachlan, J. Uterine adenocarcinoma in mice following developmental treatment with estrogens: A model for hormonal carcinogenesis. Cancer Res. 1990, 50, 7677–7681. [Google Scholar]
- Zhu, L.; Xiao, L.; Xia, Y.; Zhou, K.; Wang, H.; Huang, M.; Ge, G.; Wu, Y.; Wu, G.; Yang, L. Diethylstilbestrol can effectively accelerate estradiol-17-O-glucuronidation, while potently inhibiting estradiol-3-O-glucuronidation. Toxicol. Appl. Pharmacol. 2015, 283, 109–116. [Google Scholar] [CrossRef]
- Grenader, T.; Plotkin, Y.; Gips, M.; Cherny, N.; Gabizon, A. Diethylstilbestrol for the treatment of patients with castration-resistant prostate cancer: Retrospective analysis of a single institution experience. Oncol. Rep. 2014, 31, 428–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diel, P.; Olff, S.; Schmidt, S.; Michna, H. Effects of the environmental estrogens bisphenol A, o,p′-DDT, p-tert-octylphenol and coumestrol on apoptosis induction, cell proliferation and the expression of estrogen sensitive molecular parameters in the human breast cancer cell line MCF-7. J. Steroid Biochem. Mol. Biol. 2002, 80, 61–70. [Google Scholar] [CrossRef]
- McKinlay, R.; Plant, J.; Bell, J.; Voulvoulis, N. Endocrine disrupting pesticides: Implications for risk assessment. Environ. Int. 2008, 34, 168–183. [Google Scholar] [CrossRef]
- Watson, C.S.; Bulayeva, N.N.; Wozniak, A.L.; Alyea, R.A. Xenoestrogens are potent activators of nongenomic estrogenic responses. Steroids 2007, 72, 124–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hale, G.E.; Hughes, C.L.; Cline, J.M. Endometrial Cancer: Hormonal Factors, the Perimenopausal “Window of Risk,” and Isoflavones. J. Clin. Endocrinol. Metab. 2002, 87, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Gier, K.; Preininger, C.; Sauer, U. A Chip for Estrogen Receptor Action: Detection of Biomarkers Released by MCF-7 Cells through Estrogenic and Anti-Estrogenic Effects. Sensors 2017, 17, 1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecker, M.; Giesy, J.P. Novel trends in endocrine disruptor testing: The H295R Steroidogenesis Assay for identification of inducers and inhibitors of hormone production. Anal. Bioanal. Chem. 2008, 390, 287–291. [Google Scholar] [CrossRef]
- Soto, A.M.; Sonnenschein, C.; Chung, K.L.; Fernandez, M.F.; Olea, N.; Serrano, F.O. The E-SCREEN assay as a tool to identify estrogens: An update on estrogenic environmental pollutants. Environ. Health Perspect. 1995, 103 (Suppl. 7), 113–122. [Google Scholar] [PubMed] [Green Version]
- Van der Burg, B.; Winter, R.; Weimer, M.; Berckmans, P.; Suzuki, G.; Gijsbers, L.; Jonas, A.; van der Linden, S.; Witters, H.; Aarts, J.; et al. Optimization and prevalidation of the in vitro ERalpha CALUX method to test estrogenic and antiestrogenic activity of compounds. Reprod. Toxicol. 2010, 30, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.O. Overview of in vitro tools to assess the estrogenic and antiestrogenic activity of phytoestrogens. J. Chromatogr. B 2002, 777, 155–165. [Google Scholar] [CrossRef]
- Bolger, R.; Wiese, T.E.; Ervin, K.; Nestich, S.; Checovich, W. Rapid screening of environmental chemicals for estrogen receptor binding capacity. Environ. Health Perspect. 1998, 106, 551–557. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Carlsson, B.O.; Grandien KA, J.; Enmark, E.; Häggblad, J.; Nilsson, S.; Gustafsson, J.A. Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 1997, 138, 863–870. [Google Scholar] [CrossRef]
- Nikov, G.N.; Hopkins, N.E.; Boue, S.; Alworth, W.L. Interactions of dietary estrogens with human estrogen receptors and the effect on estrogen receptor-estrogen response element complex formation. Environ. Health Perspect. 2000, 108, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Boyer, M.; Poujol, N.; Margeat, E.; Royer, C.A. Quantitative characterization of the interaction between purified human estrogen receptor and DNA using fluorescence anisotropy. Nucleic Acids Res. 2000, 28, 2494–2502. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.O.; Hall, J.M.; Swope, D.L.; Pedersen, L.C.; Korach, K.S. Molecular determinants of the stereoselectivity of agonist activity of estrogen receptors (ER) alpha and beta. J. Biol. Chem. 2003, 278, 12255–12262. [Google Scholar] [CrossRef] [Green Version]
- Routledge, E.J.; White, R.; Parker, M.G.; Sumpter, J.P. Differential effects of xenoestrogens on coactivator recruitment by estrogen receptor (ER) alpha and ERbeta. J. Biol. Chem. 2000, 275, 35986–35993. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Cummings, R.; Li, Y.; Mitra, S.; Wilkinson, H.A.; Elbrecht, A.; Hermes, J.D.; Schaeffer, J.M.; Smith, R.G.; Moller, D.E. Nuclear receptors have distinct affinities for coactivators: Characterization by fluorescence resonance energy transfer. Mol. Endocrinol. 1998, 12, 1594–1604. [Google Scholar] [CrossRef]
- Jorgensen, M.; Vendelbo, B.; Skakkebaek, N.E.; Leffers, H. Assaying Estrogenicity by Quantitating the Expression Levels of Endogenous Estrogen-Regulated Genes. Environ. Health Perspect. 2000, 108, 403. [Google Scholar] [CrossRef] [Green Version]
- Nuwaysir, E.F.; Bittner, M.; Trent, J.; Barrett, J.C.; Afshari, C.A. Microarrays and toxicology: The advent of toxicogenomics. Mol. Carcinog. 1999, 24, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.O.; Kling, M.; Firzani, P.A.; Mecky, A.; Duranti, E.; Shields-Botella, J.; Delansorne, R.; Broschard, T.; Kramer, P.J. Activation of estrogen receptor alpha and ERbeta by 4-methylbenzylidene-camphor in human and rat cells: Comparison with phyto- and xenoestrogens. Toxicol. Lett. 2003, 142, 89–101. [Google Scholar] [CrossRef]
- Holinka, C.F.; Hata, H.; Kuramoto, H.; Gurpide, E. Effects of steroid hormones and antisteroids on alkaline phosphatase activity in human endometrial cancer cells (Ishikawa line). Cancer Res. 1986, 46, 2771–2774. [Google Scholar]
- Sanderson, J.; Boerma, J.; Lansbergen, G.W.; Berg, M.V.D. Induction and Inhibition of Aromatase (CYP19) Activity by Various Classes of Pesticides in H295R Human Adrenocortical Carcinoma Cells. Toxicol. Appl. Pharmacol. 2002, 182, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Andò, S.; Sirianni, R.; Forastieri, P.; Casaburi, I.; Lanzino, M.; Rago, V.; Giordano, F.; Carpino, A.; Pezzi, V.; Giordano, C. Aromatase expression in prepuberal Sertoli cells: Effect of thyroid hormone. Mol. Cell. Endocrinol. 2001, 178, 11–21. [Google Scholar] [CrossRef]
- Gray, L.E., Jr.; Ostby, J.; Wilson, V.; Lambright, C.; Bobseine, K.; Hartig, P.; Hotchkiss, A.; Wolf, C.; Furr, J.; Price, M.; et al. Xenoendocrine disrupters-tiered screening and testing: Filling key data gaps. Toxicology 2002, 181, 371–382. [Google Scholar] [CrossRef]
- OECD. OECD, Appraisal of test methods for sex hormone disrupting chemicals. In OECD Environment Health and Safety Publications Series on Testing and Assessment; No 21; Environment Directorate OECD: Paris, France, 2001. [Google Scholar]
- Rasmussen, T.H.; Nielsen, F.; Andersen, H.R.; Nielsen, J.B.; Weihe, P.; Grandjean, P. Assessment of xenoestrogenic exposure by a biomarker approach: Application of the E-Screen bioassay to determine estrogenic response of serum extracts. Environ. Health 2003, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- De Paoli, M.; Gogalic, S.; Sauer, U.; Preininger, C.; Pandha, H.; Simpson, G.; Horvath, A.; Marquette, C. Multiplatform Biomarker Discovery for Bladder Cancer Recurrence Diagnosis. Dis. Markers 2016, 2016, 4591910. [Google Scholar] [CrossRef] [Green Version]
- Piraino, F.; Volpetti, F.; Watson, C.; Maerkl, S.J. A Digital–Analog Microfluidic Platform for Patient-Centric Multiplexed Biomarker Diagnostics of Ultralow Volume Samples. ACS Nano 2016, 10, 1699–1710. [Google Scholar] [CrossRef] [PubMed]
- Sanjay, S.T.; Fu, G.; Dou, M.; Xu, F.; Liu, R.; Qi, H.; Li, X. Biomarker detection for disease diagnosis using cost-effective microfluidic platforms. Analyst 2015, 140, 7062–7081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchegger, P.; Sauer, U.; Toth-Székély, H.; Preininger, C. Miniaturized Protein Microarray with Internal Calibration as Point-of-Care Device for Diagnosis of Neonatal Sepsis. Sensors 2012, 12, 1494–1508. [Google Scholar] [CrossRef] [Green Version]
- Rogers, J.M.; Denison, M.S. Recombinant cell bioassays for endocrine disruptors: Development of a stably transfected human ovarian cell line for the detection of estrogenic and anti-estrogenic chemicals. In Vitr. Mol. Toxicol. 2000, 13, 67–82. [Google Scholar]
- Liu, L.; Zhou, X.; Lu, Y.; Shi, H.; Ma, M.; Yu, T. Triple functional small-molecule-protein conjugate mediated optical biosensor for quantification of estrogenic activities in water samples. Environ. Int. 2019, 132, 105091. [Google Scholar] [CrossRef] [PubMed]
- Routledge, E.J.; Sumpter, J.P. Estrogenic activity of surfactants and some of their degradation products assessed using a recombinant yeast screen. Environ. Toxicol. Chem. 1996, 15, 241–248. [Google Scholar] [CrossRef]
- Colborn, T.; Clement, C. Chemically-Induced Alterations in Sexual and Functional Development: The Wildlife/Human Connection; Princeton Scientific Pub. Co.: Princeton, NJ, USA, 1992; p. 21. [Google Scholar]
- Gray, L.; Kelce, W.R.; Wiese, T.; Tyl, R.; Gaido, K.; Cook, J.; Klinefelter, G.; Desaulniers, D.; Wilson, E.; Zacharewski, T.; et al. Endocrine screening methods workshop report: Detection of estrogenic and androgenic hormonal and antihormonal activity for chemicals that act via receptor or steroidogenic enzyme mechanisms. Reprod. Toxicol. 1997, 11, 719–750. [Google Scholar] [CrossRef]
- Safe, S.; Wang, F.; Porter, W.; Duan, R.; McDougal, A. Ah receptor agonists as endocrine disruptors: Antiestrogenic activity and mechanisms. Toxicol. Lett. 1998, 102–103, 343–347. [Google Scholar] [CrossRef]
- Green, S.; Walter, P.; Greene, G.; Krust, A.; Goffin, C.; Jensen, E.; Scrace, G.; Waterfield, M.; Chambon, P. Cloning of the human oestrogen receptor cDNA. J. Steroid Biochem. 1986, 24, 77–83. [Google Scholar] [CrossRef]
- Green, S.; Walter, P.; Kumar, V.; Krust, A.; Bornert, J.-M.; Argos, P.; Chambon, P. Human oestrogen receptor cDNA: Sequence, expression and homology to v-erb-A. Nature 1986, 320, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Mosselman, S.; Polman, J.; Dijkema, R. ER beta: Identification and characterization of a novel human estrogen receptor. FEBS Lett. 1996, 392, 49–53. [Google Scholar] [CrossRef] [Green Version]
- Kuiper, G.G.; Enmark, E.; Pelto-Huikko, M.; Nilsson, S.; Gustafsson, J.A. Cloning of a novel receptor expressed in rat prostate and ovary. Proc. Natl. Acad. Sci. USA 1996, 93, 5925–5930. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.O.; Korach, K.S. Estrogen receptors and endocrine diseases: Lessons from estrogen receptor knockout mice. Curr. Opin. Pharmacol. 2001, 1, 613–619. [Google Scholar] [CrossRef]
- Couse, J.F.; Korach, K.S. Estrogen receptor null mice: What have we learned and where will they lead us? Endocr. Rev. 1999, 20, 358–417. [Google Scholar] [CrossRef]
- Prins, G.S.; Korach, K.S. The role of estrogens and estrogen receptors in normal prostate growth and disease. Steroids 2008, 73, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Woodruff, T.K.; Mayo, K.E. To beta or not to beta: Estrogen receptors and ovarian function. Endocrinology 2005, 146, 3244–3246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krege, J.H.; Hodgin, J.B.; Couse, J.F.; Enmark, E.; Warner, M.; Mahler, J.F.; Sar, M.; Korach, K.S.; Gustafsson, J.Å.; Smithies, O. Generation and reproductive phenotypes of mice lacking estrogen receptor beta. Proc. Natl. Acad. Sci. USA 1998, 95, 15677–15682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuiper, G.G.; Lemmen, J.G.; Carlsson, B.O.; Corton, J.C.; Safe, S.H.; Van Der Saag, P.T.; Van Der Burg, B.; Gustafsson, J.A. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef] [PubMed]
- Chieffi, P.; Varriale, B. Estrogen receptor beta localization in the lizard (Podarcis s. sicula) testis. Zygote 2004, 12, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.-P.; Hsu, T.; Hu, C.-H.; Wang, W.-D.; Wang, K.-C.; Teng, L.-F. Expression of estrogen receptors alpha and beta in human osteoblasts: Identification of exon-2 deletion variant of estrogen receptor beta in postmenopausal women. Chang. Gung Med. J. 2004, 27, 107–115. [Google Scholar]
- Nilsson, L.O.; Boman, A.; Sävendahl, L.; Grigelioniene, G.; Ohlsson, C.; Ritzén, E.M.; Wroblewski, J. Demonstration of estrogen receptor-beta immunoreactivity in human growth plate cartilage. J. Clin. Endocrinol. Metab. 1999, 84, 370–373. [Google Scholar]
- Pedeutour, F.; Quade, B.J.; Weremowicz, S.; Cin, P.D.; Ali, S.; Morton, C.C. Localization and expression of the human estrogen receptor beta gene in uterine leiomyomata. Genes Chromosom. Cancer 1998, 23, 361–366. [Google Scholar] [CrossRef]
- Hörlein, A.J.; Näär, A.M.; Heinzel, T.; Torchia, J.; Gloss, B.; Kurokawa, R.; Ryan, A.; Kamei, Y.; Söderström, M.; Glass, C.K.; et al. Ligand-independent repression by the thyroid hormone receptor mediated by a nuclear receptor co-repressor. Nature 1995, 377, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Wolffe, A.P. Chromatin remodeling regulated by steroid and nuclear receptors. Cell Res. 1997, 7, 127–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.M.; McDonnell, D.P. The estrogen receptor beta-isoform (ERbeta) of the human estrogen receptor modulates ERalpha transcriptional activity and is a key regulator of the cellular response to estrogens and antiestrogens. Endocrinology 1999, 140, 5566–5578. [Google Scholar] [CrossRef] [PubMed]
- McKenna, N.J.; O’Malley, B.W. Minireview: Nuclear Receptor Coactivators—An Update. Endocrinology 2002, 143, 2461–2465. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.M.; Albanese, C.; Anderson, C.M.; Hilty, K.; Webb, P.; Uht, R.M.; Price, R.H., Jr.; Pestell, R.G.; Kushner, P.J. Opposing action of estrogen receptors alpha and beta on cyclin D1 gene expression. J. Biol. Chem. 2002, 277, 24353–24360. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, S. Roles of Estrogen Receptor- Gene Expression in Reproduction-Related Behaviors in Female Mice. Endocrinology 1998, 139, 5070–5081. [Google Scholar] [CrossRef] [PubMed]
- Cowley, S.M.; Hoare, S.; Mosselman, S.; Parker, M.G. Estrogen receptors alpha and beta form heterodimers on DNA. J. Biol. Chem. 1997, 272, 19858–19862. [Google Scholar] [CrossRef] [Green Version]
- McKenna, N.J.; Lanz, R.B.; O’Malley, B.W. Nuclear Receptor Coregulators: Cellular and Molecular Biology. Endocr. Rev. 1999, 20, 321–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.D.; Evans, R.M. A transcriptional co-repressor that interacts with nuclear hormone receptors. Nat. Cell Biol. 1995, 377, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Kurdoglu, M.; Yildirim, M.; Kurdoglu, Z.; Erdem, A.; Erdem, M.; Bilgihan, A.; Goktas, B. Cardiovascular risk assessment with oxidised LDL measurement in postmenopausal women receiving intranasal estrogen replacement therapy. Gynecol. Endocrinol. 2011, 27, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Iyer, S.; Martin-Millan, M.; Bartell, S.M.; Han, L.; Ambrogini, E.; Onal, M.; Xiong, J.; Weinstein, R.S.; Jilka, R.L.; et al. Estrogen receptor-α signaling in osteoblast progenitors stimulates cortical bone accrual. J. Clin. Investig. 2013, 123, 394–404. [Google Scholar] [CrossRef] [Green Version]
- Windahl, S.H.; Borjesson, A.E.; Farman, H.H.; Engdahl, C.; Moverare-Skrtic, S.; Sjogren, K.; Lagerquist, M.K.; Kindblom, J.M.; Koskela, A.; Tuukkanen, J.; et al. Estrogen receptor- in osteocytes is important for trabecular bone formation in male mice. Proc. Natl. Acad. Sci. USA 2013, 110, 2294–2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapara, A.; Brunton, V.G.; Graham, D.; Faulds, K. Characterisation of estrogen receptor alpha (ERalpha) expression in breast cancer cells and effect of drug treatment using targeted nanoparticles and SERS. Analyst 2020, 145, 7225–7233. [Google Scholar] [CrossRef]
- Topi, G.; Satapathy, S.R.; Dash, P.; Fred Mehrabi, S.; Ehrnström, R.; Olsson, R.; Lydrup, M.L.; Sjölander, A. Tumour-suppressive effect of oestrogen receptor beta in colorectal cancer patients, colon cancer cells, and a zebrafish model. J. Pathol. 2020, 251, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; Zhang, S.; Wei, Y.; Tian, S.; Lu, Z.; Jin, L.; He, Y.; Xie, W.; Li, Y. Structural Insights into the Specificity of Ligand Binding and Coactivator Assembly by Estrogen-Related Receptor beta. J. Mol. Biol. 2020, 432, 5460–5472. [Google Scholar] [CrossRef]
- Casanova, G.; Spritzer, P.M. Effects of micronized progesterone added to non-oral estradiol on lipids and cardiovascular risk factors in early postmenopause: A clinical trial. Lipids Health Dis. 2012, 11, 133. [Google Scholar] [CrossRef] [Green Version]
- Stockard, C.R.; Papanicolaou, G.N. The existence of a typical oestrous cycle in the guinea-pig—with a study of its histological and physiological changes. Am. J. Anat. 1917, 22, 225–283. [Google Scholar] [CrossRef]
- Parada, H.; Wolff, M.S.; Engel, L.S.; Eng, S.M.; Khankari, N.K.; Neugut, A.I.; Teitelbaum, S.L.; Gammon, M.D. Polychlorinated biphenyls and their association with survival following breast cancer. Eur. J. Cancer 2016, 56, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Kerdivel, G.; Habauzit, D.; Pakdel, F. Assessment and Molecular Actions of Endocrine-Disrupting Chemicals That Interfere with Estrogen Receptor Pathways. Int. J. Endocrinol. 2013, 2013, 501851. [Google Scholar] [CrossRef] [Green Version]
- Xue, N.; Wang, H.; Xu, X. Progress in study on endocrine disrupting pesticides (EDPs) in aquatic environment. Chin. Sci. Bull. 2005, 50, 2257–2266. [Google Scholar] [CrossRef]
- Vigh, E.; Garai, J. Hormonal Action of Plant Derived and Anthropogenic Non-Steroidal Estrogenic Compounds: Phytoestrogens and Xenoestrogens. Curr. Med. Chem. 2010, 17, 3542–3574. [Google Scholar] [CrossRef]
- Jagadeesh, S.; Kyo, S.; Banerjee, P.P. Genistein Represses Telomerase Activity via Both Transcriptional and Posttranslational Mechanisms in Human Prostate Cancer Cells. Cancer Res. 2006, 66, 2107–2115. [Google Scholar] [CrossRef] [Green Version]
- Watson, P.H.; Pon, R.T.; Shiu, R.P. Inhibition of c-myc expression by phosphorothioate antisense oligonucleotide identifies a critical role for c-myc in the growth of human breast cancer. Cancer Res. 1991, 51, 3996–4000. [Google Scholar]
- Hassan, S.; Siddiqui, G.; Trudgett, A.; Robert, D.; Zhao, Y.; Wang, X. Assessment of reproductive disorder (imposex) induced by tributyltins in marine gastropods. Pak. J. Pharm. Sci. 2019, 32, 1987–1993. [Google Scholar]
- Merlo, E.; Silva, I.V.; Cardoso, R.C.; Graceli, J.B. The obesogen tributyltin induces features of polycystic ovary syndrome (PCOS): A review. J. Toxicol. Environ. Health Part B 2018, 21, 181–206. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, C.S.; Miranda-Alves, L.; La Merrill, M.A.; Silva, I.V.; Graceli, J.B. The tributyltin leads to obesogenic mammary gland abnormalities in adult female rats. Toxicol. Lett. 2019, 307, 59–71. [Google Scholar] [CrossRef]
- Kanimozhi, V.; Palanivel, K.; Kadalmani, B.; Akbarsha, M. Molecular mechanisms of tributyltin-induced alterations in cholesterol homeostasis and steroidogenesis in hamster testis: In vivo and in vitro studies. J. Cell. Biochem. 2018, 119, 4021–4037. [Google Scholar] [CrossRef] [PubMed]
- Grün, F. The Obesogen Tributyltin. Vitam. Horm. 2014, 94, 277–325. [Google Scholar] [CrossRef]
- Boyer, I.J. Toxicity of dibutyltin, tributyltin and other organotin compounds to humans and to experimental animals. Toxicology 1989, 55, 253–298. [Google Scholar] [CrossRef]
- Gbeddy, G.; Goonetilleke, A.; Ayoko, G.A.; Egodawatta, P. Transformation and degradation of polycyclic aromatic hydrocarbons (PAHs) in urban road surfaces: Influential factors, implications and recommendations. Environ. Pollut. 2020, 257, 113510. [Google Scholar] [CrossRef] [PubMed]
- Singh, L.; Varshney, J.G.; Agarwal, T. Polycyclic aromatic hydrocarbons’ formation and occurrence in processed food. Food Chem. 2016, 199, 768–781. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Jahan, S.A.; Kabir, E.; Brown, R.J. A review of airborne polycyclic aromatic hydrocarbons (PAHs) and their human health effects. Environ. Int. 2013, 60, 71–80. [Google Scholar] [CrossRef]
- Guillette, L.J.; Gross, T.S.; Masson, G.R.; Matter, J.M.; Percival, H.F.; Woodward, A.R. Developmental Abnormalities of the Gonad and Abnormal Sex Hormone Concentrations in Juvenile Alligators from Contaminated and Control Lakes in Florida. Environ. Health Perspect. 1994, 102, 680. [Google Scholar] [CrossRef]
- Fujisaki, I.; Rice, K.G.; Woodward, A.R.; Percival, H.F. Possible Generational Effects of Habitat Degradation on Alligator Reproduction. J. Wildl. Manag. 2007, 71, 2284. [Google Scholar] [CrossRef]
- Wolff, M.S.; Toniolo, P.G.; Lee, E.W.; Rivera, M.; Dubin, N. Blood Levels of Organochlorine Residues and Risk of Breast Cancer. J. Natl. Cancer Inst. 1993, 85, 648–652. [Google Scholar] [CrossRef]
- Bergeron, J.M.; Crews, D.; A McLachlan, J. PCBs as Environmental Estrogens: Turtle Sex Determination as a Biomarker of Environmental Contamination. Environ. Health Perspect. 1994, 102, 780. [Google Scholar] [CrossRef]
- Sager, D.B.; Shih-Schroeder, W.; Girard, D. Effect of early postnatal exposure to polychlorinated biphenyls (PCBs) on fertility in male rats. Bull. Environ. Contam. Toxicol. 1987, 38, 946–953. [Google Scholar] [CrossRef]
- Dieringer, C.S.; Lamartiniere, C.A.; Schiller, C.M.; Lucier, G.W. Altered ontogeny of hepatic steroid-metabolizing enzymes by pure polychlorinated biphenyl congeners. Biochem. Pharmacol. 1979, 28, 2511–2514. [Google Scholar] [CrossRef]
- Parada, H., Jr.; Wolff, M.S.; Engel, L.S.; White, A.J.; Eng, S.M.; Cleveland, R.J.; Khankari, N.K.; Teitelbaum, S.L.; Neugut, A.I.; Gammon, M.D. Organochlorine insecticides DDT and chlordane in relation to survival following breast cancer. Int. J. Cancer 2016, 138, 565–575. [Google Scholar] [CrossRef] [Green Version]
- Newcomb, P.A.; Lantz, P.M. Recent trends in breast cancer incidence, mortality, and mammography. Breast Cancer Res. Treat. 1993, 28, 97–106. [Google Scholar] [CrossRef]
- E Henderson, B.; Ross, R.K.; Pike, M.C. Hormonal chemoprevention of cancer in women. Science 1993, 259, 633–638. [Google Scholar] [CrossRef]
- Feuer, E.J.; Wun, L.M. How much of the recent rise in breast cancer incidence can be explained by increases in mammography utilization? A dynamic population model approach. Am. J. Epidemiol. 1992, 136, 1423–1436. [Google Scholar] [CrossRef]
- Birnbaum, L.S.; E Fenton, S. Cancer and developmental exposure to endocrine disruptors. Environ. Health Perspect. 2003, 111, 389–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Hond, E.; Roels, H.A.; Hoppenbrouwers, K.; Nawrot, T.; Thijs, L.; Vandermeulen, C.; Winneke, G.; Vanderschueren, D.; Staessen, J.A. Sexual maturation in relation to polychlorinated aromatic hydrocarbons: Sharpe and Skakkebaek’s hypothesis revisited. Environ. Health Perspect. 2002, 110, 771–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, A.M.; Brisken, C.; Schaeberle, C.; Sonnenschein, C. Does cancer start in the womb? Altered mammary gland development and predisposition to breast cancer due to in utero exposure to endocrine disruptors. J. Mammary Gland Biol. Neoplasia 2013, 18, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Villeneuve, S.; Févotte, J.; Anger, A.; Truong, T.; Lamkarkach, F.; Gaye, O.; Kerbrat, P.; Arveux, P.; Miglianico, L.; Imbernon, E.; et al. Breast cancer risk by occupation and industry: Analysis of the CECILE study, a population-based case-control study in France. Am. J. Ind. Med. 2011, 54, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Labrèche, F.; Goldberg, M.S.; Valois, M.-F.; Nadon, L. Postmenopausal breast cancer and occupational exposures. Occup. Environ. Med. 2010, 67, 263–269. [Google Scholar] [CrossRef]
- Hoover, R.N.; Hyer, M.; Pfeiffer, R.M.; Adam, E.; Bond, B.; Cheville, A.L.; Colton, T.; Hartge, P.; Hatch, E.E.; Herbst, A.L.; et al. Adverse Health Outcomes in Women Exposed In Utero to Diethylstilbestrol. N. Engl. J. Med. 2011, 365, 1304–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.Y.; Izumi, S.; Suzuki, T.; Goya, K.; Nakamura, E.; Sugiyama, T.; Kobayashi, H. Dioxins in ascites and serum of women with endometriosis: A pilot study. Hum. Reprod. 2011, 26, 117–126. [Google Scholar] [CrossRef]
- Signorile, P.G.; Spugnini, E.P.; Mita, L.; Mellone, P.; D’Avino, A.; Bianco, M.; Diano, N.; Caputo, L.; Rea, F.; Viceconte, R.; et al. Pre-natal exposure of mice to bisphenol A elicits an endometriosis-like phenotype in female offspring. Gen. Comp. Endocrinol. 2010, 168, 318–325. [Google Scholar] [CrossRef]
- Nayyar, T.; Bruner-Tran, K.L.; Piestrzeniewicz-Ulanska, D.; Osteen, K.G. Developmental exposure of mice to TCDD elicits a similar uterine phenotype in adult animals as observed in women with endometriosis. Reprod. Toxicol. 2007, 23, 326–336. [Google Scholar] [CrossRef] [Green Version]
- Missmer, S.A.; Hankinson, S.E.; Spiegelman, D.; Barbieri, R.L.; Michels, K.B.; Hunter, D.J. In utero exposures and the incidence of endometriosis. Fertil. Steril. 2004, 82, 1501–1508. [Google Scholar] [CrossRef] [PubMed]
- Bois, F.Y.; Eskenazi, B. Possible Risk of Endometriosis for Seveso, Italy, Residents: An Assessment of Exposure to Dioxin. Environ. Health Perspect. 1994, 102, 476. [Google Scholar] [CrossRef]
- Bulun, S.E.; Cheng, Y.-H.; Pavone, M.E.; Xue, Q.; Attar, E.; Trukhacheva, E.; Tokunaga, H.; Utsunomiya, H.; Yin, P.; Luo, X.; et al. Estrogen Receptor-β, Estrogen Receptor-α, and Progesterone Resistance in Endometriosis. Semin. Reprod. Med. 2010, 28, 036–043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rier, S.E.; Martin, D.C.; Bowman, R.E.; Dmowski, W.P.; Becker, J.L. Endometriosis in rhesus monkeys (Macaca mulatta) following chronic exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Fundam. Appl. Toxicol. 1993, 21, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wang, Y.; Zhou, W.-H.; Wang, L.; He, Y.-Y.; Li, D.-J. Combination of estrogen and dioxin is involved in the pathogenesis of endometriosis by promoting chemokine secretion and invasion of endometrial stromal cells. Hum. Reprod. 2008, 23, 1614–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bencko, V. Human exposure to endocrine disrupters: Carcinogenic risk assessment. Folia Histochem. Cytobiol. 2001, 39 (Suppl. 2), 24–25. [Google Scholar] [PubMed]
- Soto, A.M.; Fernandez, M.F.; Luizzi, M.F.; Oles Karasko, A.S.; Sonnenschein, C. Developing a marker of exposure to xenoestrogen mixtures in human serum. Environ. Health Perspect. 1997, 105 (Suppl. 3), 647–654. [Google Scholar]
- Kim, J.-Y.; Yi, B.-R.; Go, R.-E.; Hwang, K.-A.; Nam, K.-H.; Choi, K.-C. Methoxychlor and triclosan stimulates ovarian cancer growth by regulating cell cycle- and apoptosis-related genes via an estrogen receptor-dependent pathway. Environ. Toxicol. Pharmacol. 2014, 37, 1264–1274. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.M.; Korach, K.S. Endocrine disrupting chemicals promote the growth of ovarian cancer cells via the ER-CXCL12-CXCR4 signaling axis. Mol. Carcinog. 2013, 52, 715–725. [Google Scholar] [CrossRef] [Green Version]
- Dey, S.; Hablas, A.; A Seifeldin, I.; Ismail, K.; Ramadan, M.; El-Hamzawy, H.; Wilson, M.L.; Banerjee, M.; Boffetta, P.; Harford, J.; et al. Urban-rural differences of gynaecological malignancies in Egypt (1999–2002). BJOG Int. J. Obstet. Gynaecol. 2010, 117, 348–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calafat, A.M.; Ye, X.; Wong, L.-Y.; Reidy, J.A.; Needham, L.L. Exposure of the U.S. Population to Bisphenol A and 4-tertiary-Octylphenol: 2003–2004. Environ. Health Perspect. 2008, 116, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivacqua, A.; Recchia, A.G.; Fasanella, G.; Gabriele, S.; Carpino, A.; Rago, V.; Di Gioia, M.L.; Leggio, A.; Bonofiglio, D.; Liguori, A.; et al. The Food Contaminants Bisphenol A and 4-Nonylphenol Act as Agonists for Estrogen Receptor α in MCF7 Breast Cancer Cells. Endocrine 2003, 22, 275–284. [Google Scholar] [CrossRef]
- Durando, M.; Kass, L.; Piva, J.; Sonnenschein, C.; Soto, A.M.; Luque, E.H.; Muñoz-De-Toro, M. Prenatal Bisphenol A Exposure Induces Preneoplastic Lesions in the Mammary Gland in Wistar Rats. Environ. Health Perspect. 2007, 115, 80–86. [Google Scholar] [CrossRef]
- Murray, T.J.; Maffini, M.V.; Ucci, A.A.; Sonnenschein, C.; Soto, A.M. Induction of mammary gland ductal hyperplasias and carcinoma in situ following fetal bisphenol A exposure. Reprod. Toxicol. 2007, 23, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Caporossi, L.; Papaleo, B. Bisphenol A and Metabolic Diseases: Challenges for Occupational Medicine. Int. J. Environ. Res. Public Health 2017, 14, 959. [Google Scholar] [CrossRef] [Green Version]
- Somm, E.P.; Schwitzgebel, V.M.; Toulotte, A.; Cederroth, C.R.; Combescure, C.; Nef, S.; Aubert, M.L.; Hüppi, P.S. Perinatal Exposure to Bisphenol A Alters Early Adipogenesis in the Rat. Environ. Health Perspect. 2009, 117, 1549–1555. [Google Scholar] [CrossRef] [Green Version]
- Nadal, A.; Alonso-Magdalena, P.; Soriano, S.; Quesada, I.; Ropero, A.B. The pancreatic beta-cell as a target of estrogens and xenoestrogens: Implications for blood glucose homeostasis and diabetes. Mol. Cell. Endocrinol. 2009, 304, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Zhang, L.; Zhang, H.; Wei, W.; Jia, L. Perinatal BPA Exposure Induces Hyperglycemia, Oxidative Stress and Decreased Adiponectin Production in Later Life of Male Rat Offspring. Int. J. Environ. Res. Public Health 2014, 11, 3728–3742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Magdalena, P.; Ropero, A.B.; Carrera, M.P.; Cederroth, C.R.; Baquie, M.; Gauthier, B.R.; Nef, S.; Stefani, E.; Nadal, A. Pancreatic insulin content regulation by the estrogen receptor ER alpha. PLoS ONE 2008, 3, e2069. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Magdalena, P.; Morimoto, S.; Ripoll, C.; Fuentes, E.; Nadal, A. The estrogenic effect of bisphenol A disrupts pancreatic beta-cell function in vivo and induces insulin resistance. Environ. Health Perspect. 2006, 114, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menale, C.; Mita, D.G.; Diano, N.; Diano, S. Adverse Effects of Bisphenol A Exposure on Glucose Metabolism Regulation. Open Biotechnol. J. 2016, 10 (Suppl. 1), 122–130. [Google Scholar] [CrossRef] [Green Version]
- Tremblay-Franco, M.; Cabaton, N.J.; Canlet, C.; Gautier, R.; Schaeberle, C.M.; Jourdan, F.; Sonnenschein, C.; Vinson, F.; Soto, A.M.; Zalko, D. Dynamic Metabolic Disruption in Rats Perinatally Exposed to Low Doses of Bisphenol-A. PLoS ONE 2015, 10, e0141698. [Google Scholar] [CrossRef] [PubMed]
- Menale, C.; Grandone, A.; Nicolucci, C.; Cirillo, G.; Crispi, S.; Di Sessa, A.; Marzuillo, P.; Rossi, S.; Mita, D.G.; Perrone, L.; et al. Bisphenol A is associated with insulin resistance and modulates adiponectin and resistin gene expression in obese children. Pediatr. Obes. 2017, 12, 380–387. [Google Scholar] [CrossRef]
- Fénichel, P.; Chevalier, N. Environmental endocrine disruptors: New diabetogens? Comptes Rendus Biol. 2017, 340, 446–452. [Google Scholar] [CrossRef]
- Chin, K.-Y.; Pang, K.-L.; Mark-Lee, W.F. A Review on the Effects of Bisphenol A and Its Derivatives on Skeletal Health. Int. J. Med Sci. 2018, 15, 1043–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Oh, C.H.; Hwang, Y.-C.; Jeong, I.-K.; Ahn, K.J.; Chung, H.-Y.; Chang, J.-S. Serum Bisphenol A Concentration in Postmenopausal Women with Osteoporosis. J. Bone Metab. 2012, 19, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Upson, K.; Sathyanarayana, S.; De Roos, A.J.; Koch, H.M.; Scholes, D.; Holt, V.L. A population-based case–control study of urinary bisphenol A concentrations and risk of endometriosis. Hum. Reprod. 2014, 29, 2457–2464. [Google Scholar] [CrossRef] [Green Version]
- Sidorkiewicz, I.; Zaręba, K.; Wołczyński, S.; Czerniecki, J. Endocrine-disrupting chemicals—Mechanisms of action on male reproductive system. Toxicol. Ind. Health 2017, 33, 601–609. [Google Scholar] [CrossRef]
- Marques-Pinto, A.; Carvalho, D. Human infertility: Are endocrine disruptors to blame? Endocr. Connect. 2013, 2, R15–R29. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Duan, W.; Li, R.; Xu, S.; Zhang, L.; Chen, C.; He, M.; Lu, Y.; Wu, H.; Pi, H.; et al. Exposure to bisphenol A disrupts meiotic progression during spermatogenesis in adult rats through estrogen-like activity. Cell Death Dis. 2013, 4, e676. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Bu, P.; Li, F.; Lan, S.; Wu, H.; Yuan, L.; Wang, Y. Neonatal bisphenol A exposure induces meiotic arrest and apoptosis of spermatogenic cells. Oncotarget 2016, 7, 10606–10615. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.-L.; Zhang, X.-F.; Feng, Y.-M.; Li, L.; Huynh, E.; Sun, X.-F.; Sun, Z.-Y.; Shen, W. Exposure to bisphenol A results in a decline in mouse spermatogenesis. Reprod. Fertil. Dev. 2013, 25, 847–859. [Google Scholar] [CrossRef]
- Aly, H.A.; Hassan, M.H.; A El-Beshbishy, H.; Alahdal, A.M.; Osman, A.-M.M. Dibutyl phthalate induces oxidative stress and impairs spermatogenesis in adult rats. Toxicol. Ind. Health 2016, 32, 1467–1477. [Google Scholar] [CrossRef] [PubMed]
- Aly, H.A.; Azhar, A.S. Methoxychlor induced biochemical alterations and disruption of spermatogenesis in adult rats. Reprod. Toxicol. 2013, 40, 8–15. [Google Scholar] [CrossRef] [PubMed]
- A Ronchetti, S.; Novack, G.V.; Bianchi, M.S.; Crocco, M.C.; Duvilanski, B.H.; Cabilla, J.P. In vivo xenoestrogenic actions of cadmium and arsenic in anterior pituitary and uterus. Reproduction 2016, 152, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tapiero, H.; Ba, G.N.; Tew, K. Estrogens and environmental estrogens. Biomed. Pharm. 2002, 56, 36–44. [Google Scholar] [CrossRef]
- Rosenfeld, C.S. Effects of Phytoestrogens on the Developing Brain, Gut Microbiota, and Risk for Neurobehavioral Disorders. Front. Nutr. 2019, 6, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeComte, S.; DeMay, F.; Ferrière, F.; Pakdel, F. Phytochemicals Targeting Estrogen Receptors: Beneficial Rather Than Adverse Effects? Int. J. Mol. Sci. 2017, 18, 1381. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-R.; Ko, N.-Y.; Chen, K.-H. Isoflavone Supplements for Menopausal Women: A Systematic Review. Nutrients 2019, 11, 2649. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-R.; Chen, K.-H. Utilization of Isoflavones in Soybeans for Women with Menopausal Syndrome: An Overview. Int. J. Mol. Sci. 2021, 22, 3212. [Google Scholar] [CrossRef] [PubMed]
- Paulose, T.; Speroni, L.; Sonnenschein, C.; Soto, A.M. Estrogens in the wrong place at the wrong time: Fetal BPA exposure and mammary cancer. Reprod. Toxicol. 2015, 54, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Jafari, A.; Moeckel, C.; Jones, K.C. Spatial biomonitoring of persistent organic pollutants in Iran: A study using locally produced butter. J. Environ. Monit. 2008, 10, 861–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitada, Y.; Kawahata, H.; Suzuki, A.; Oomori, T. Distribution of pesticides and bisphenol A in sediments collected from rivers adjacent to coral reefs. Chemosphere 2008, 71, 2082–2090. [Google Scholar] [CrossRef] [PubMed]
- Gouin, T.; Jantunen, L.; Harner, T.; Blanchard, P.; Bidleman, T. Spatial and Temporal Trends of Chiral Organochlorine Signatures in Great Lakes Air Using Passive Air Samplers. Environ. Sci. Technol. 2007, 41, 3877–3883. [Google Scholar] [CrossRef] [PubMed]
- Kumari, B.; Singh, J.; Singh, S.; Kathpal, T.S. Monitoring of Butter and Ghee (Clarified Butter Fat) for Pesticidal Contamination from Cotton Belt of Haryana, India. Environ. Monit. Assess. 2005, 105, 111–120. [Google Scholar] [CrossRef]
- Voisin, A.-S.; Kültz, D.; Silvestre, F. Early-life exposure to the endocrine disruptor 17-α-ethinylestradiol induces delayed effects in adult brain, liver and ovotestis proteomes of a self-fertilizing fish. J. Proteom. 2019, 194, 112–124. [Google Scholar] [CrossRef]
- Hill, R.L.; Janz, D.M. Developmental estrogenic exposure in zebrafish (Danio rerio): I. Effects on sex ratio and breeding success. Aquat. Toxicol. 2003, 63, 417–429. [Google Scholar] [CrossRef]
- Liao, T.; Guo, Q.L.; Jin, S.W.; Cheng, W.; Xu, Y. Comparative responses in rare minnow exposed to 17beta-estradiol during different life stages. Fish Physiol. Biochem. 2009, 35, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Nasiadek, M.; Danilewicz, M.; Sitarek, K.; Świątkowska, E.; Daragó, A.; Stragierowicz, J.; Kilanowicz, A. The effect of repeated cadmium oral exposure on the level of sex hormones, estrous cyclicity, and endometrium morphometry in female rats. Environ. Sci. Pollut. Res. 2018, 25, 28025–28038. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| In Vitro Assay | Endpoint of Measurement | Advantages | Limitations | Reference |
|---|---|---|---|---|
| E-Screen | ERα (+) cell proliferation | Measures physiological endpoint of estrogen action, measures estrogens and antiestrogens | No defined ER expression, no mechanistic data | [33] |
| Ligand-binding (EDSTAC) a | ERα- or ERβ-binding affinity | Simple, high-throughput method | Does not measure ER activation does not measure physiological response | [36,37] |
| ER-binding to ERE | ERα- or ERβ-binding affinity to ERE | High-throughput method, various EREs can be used | Does not measure ER activation, low sensitivity, does not measure physiological response | [38,39] |
| GST pull-down/FRET/ two-hybrid assay | ERα- or ERβ-ligand-dependent association with coactivators | Analysis of molecular interaction, defined ER subtype or ER domain as well as coactivators can be used, measures estrogens and antiestrogens | Does not measure direct ER activation, low throughput, does not measure physiological response | [40,41,42] |
| Analysis of gene expression | ER-regulated gene expression | Analysis of physiological response, versatile, measures estrogens and antiestrogens | Low throughput | [43,44] |
| Analysis of enzyme activity | ER-regulated enzyme activity | Analysis of physiological response measures estrogens and antiestrogens | Cell lines or primary cell cultures with active marker enzymes suitable only | [45,46] |
| Analysis of steroidogenesis (EDSTAC)a | Induction and inhibition of estrogen biosynthesis | Analysis of physiological response measures ER-independent pathways | Cells with active steroidogenesis suitable only | [47,48,49,50,51] |
| The fluorescence-based multianalyte chip platform(a fluorescence-based multiplexed protein microarray) | The proliferative effect in hormone-sensitive cancer cell line MCF-7 was measured with a resazurin assay. Quantification of 10 proteins from MCF-7 cells, representing endpoints of estrogen-and antiestrogen action | High throughput screening. Multiparameter panels, fast and highly specific diagnosis | binding affinities, different concentrations and different periods will be necessary to refine the specific secretion patterns | [52,53,54,55] |
| Transcriptional activation assays | Luciferase activity | Rapid screening, identification, and characterization of EDCs, the first human ovarian cell bioassay of this kind for detecting estrogens | Response observed in cell lines does not necessarily reflect the toxic or biological potential of a compound in vivo. | [56] |
| Triple functional small-molecule–protein conjugate mediated optical biosensor | Dye-labeled estradiol (E2) -streptavidin conjugate (estrogenic activity) | Easy-to-use and efficient with the high reusable capability | Expensive | [57] |
| Estrogen receptor recombinant yeastscreening (YES)assay | Estrogenic activity |
The simplicity The product of the reporter gene is secreted in the medium, and no cell lysis is required. | The presence of yeast cell wall and active transport mechanisms that may differ from those found in mammalian cells and may affect the activity of some test compounds | [58] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.-H.; Chen, L.-R.; Chen, K.-H. In Vitro and Vivo Identification, Metabolism and Action of Xenoestrogens: An Overview. Int. J. Mol. Sci. 2021, 22, 4013. https://doi.org/10.3390/ijms22084013
Wang L-H, Chen L-R, Chen K-H. In Vitro and Vivo Identification, Metabolism and Action of Xenoestrogens: An Overview. International Journal of Molecular Sciences. 2021; 22(8):4013. https://doi.org/10.3390/ijms22084013
Chicago/Turabian StyleWang, Li-Hsuan, Li-Ru Chen, and Kuo-Hu Chen. 2021. "In Vitro and Vivo Identification, Metabolism and Action of Xenoestrogens: An Overview" International Journal of Molecular Sciences 22, no. 8: 4013. https://doi.org/10.3390/ijms22084013
APA StyleWang, L. -H., Chen, L. -R., & Chen, K. -H. (2021). In Vitro and Vivo Identification, Metabolism and Action of Xenoestrogens: An Overview. International Journal of Molecular Sciences, 22(8), 4013. https://doi.org/10.3390/ijms22084013