
Molecular Chaperones and Thyroid Cancer

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
1.1. Thyroid Tumors
1.2. The Chaperone System
2. Quantitative Changes of Molecular Chaperones and Associated Effects in TC
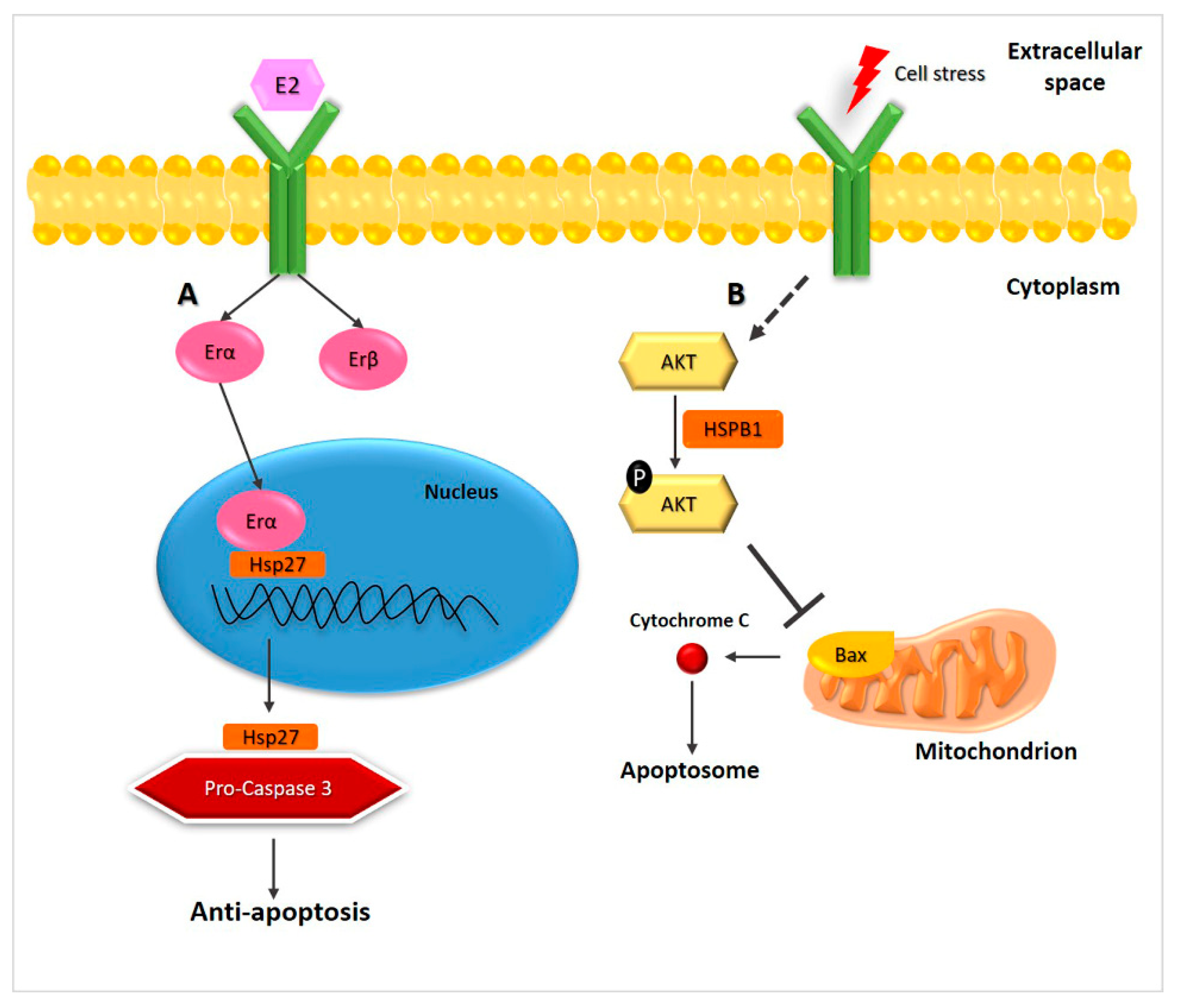
2.1. Hsp27
2.2. Hsp60
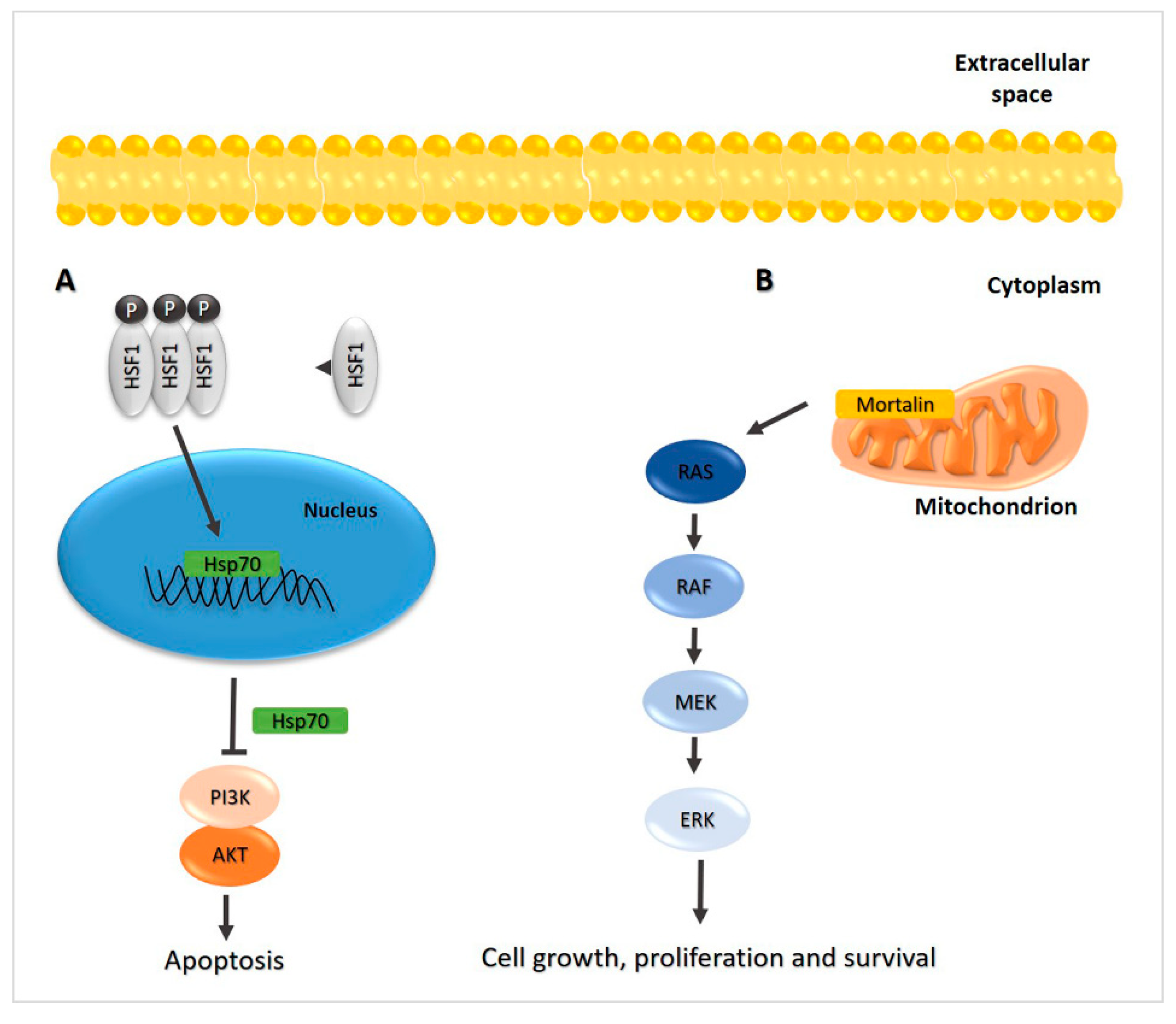
2.3. Hsp70
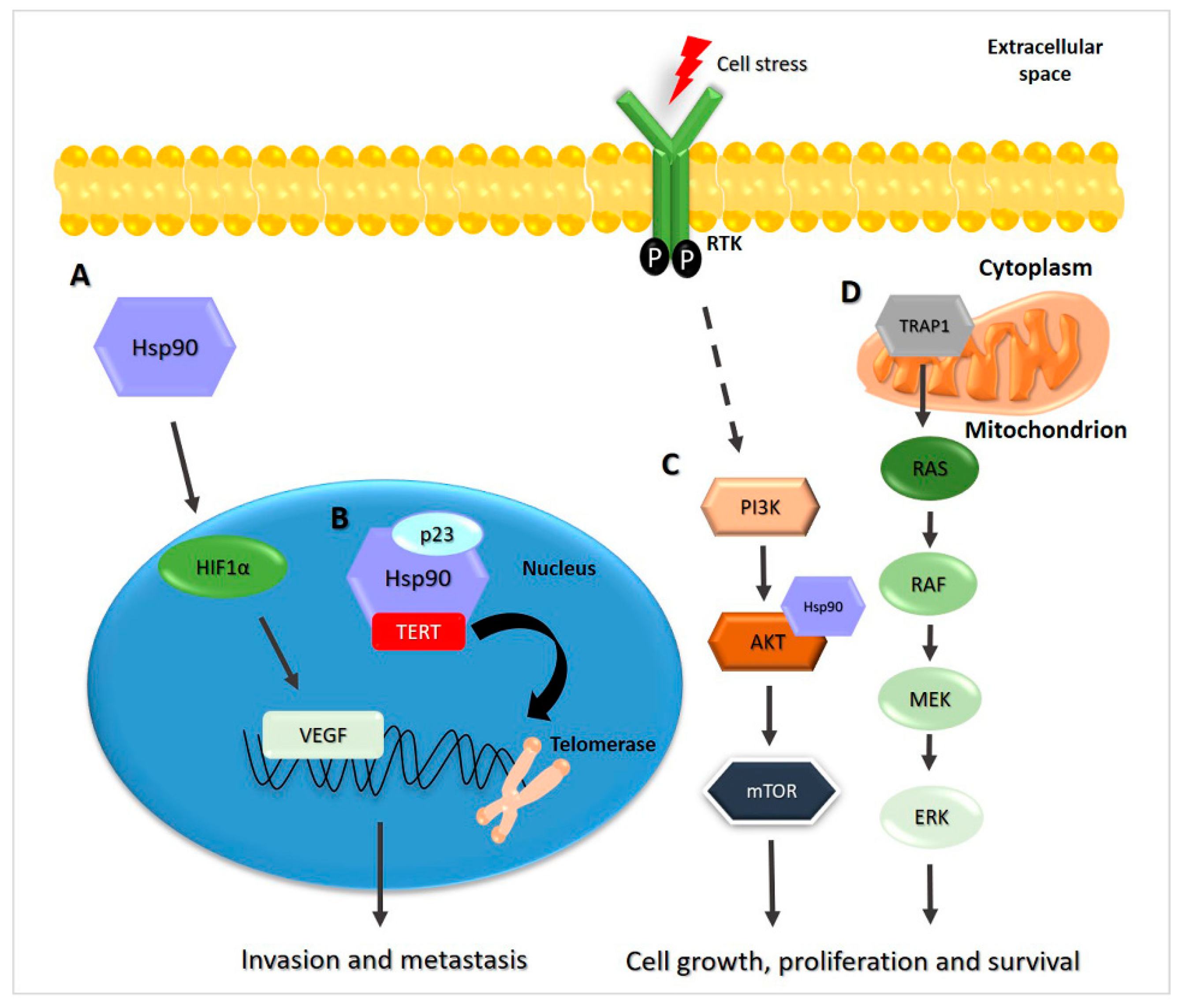
2.4. Hsp90

3. Various Chaperones Assessed Simultaneously
4. Exosomes and TC
5. Conclusions and Perspectives for the Future
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AKT | serine/threonine protein kinase B |
| ATC | anaplastic/undifferentiated thyroid carcinoma |
| Bcl-2 | B-cell lymphoma 2 |
| DM | distant metastasis |
| ERK | extracellular signal-regulated kinases |
| Erα/SP1 | estrogen receptor/specificity protein 1 |
| ERα36 | estrogen receptor alpha36 |
| ETE | extra-thyroid extension |
| EVs | extracellular vesicles |
| FA | follicular adenoma |
| FNA | fine-needle aspiration |
| FTC | follicular thyroid carcinoma |
| (HIF)-1α | hypoxia-inducible factor-1α |
| hTERT | human telomerase reverse transcriptase |
| LNM | lymph node metastasis |
| MEK | mitogen-activated protein kinase kinase |
| MTC | medullary thyroid carcinoma |
| mTOR | mammalian target of rapamycin |
| PDTC | poorly differentiated thyroid carcinoma |
| PI3K | phosphatidylinositol 3-kinase |
| PTC | papillary thyroid carcinoma |
| RAF | proto-oncogene serine/threonine protein kinase |
| ROS | reactive oxygen species |
| TC | thyroid cancer (or thyroid carcinoma) |
| TNM | tumor node metastasis stage |
| TRAP1 | tumor necrosis factor receptor-associated protein 1 |
References
- Abooshahab, R.; Gholami, M.; Sanoie, M.; Azizi, F.; Hedayati, M. Advances in metabolomics of thyroid cancer diagnosis and metabolic regulation. Endocrine 2019, 65, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lebastchi, A.H.; Callender, G.G. Thyroid cancer. Curr. Probl. Cancer 2014, 38, 48–74. [Google Scholar] [CrossRef]
- Asa, S.L. The Current Histologic Classification of Thyroid Cancer. Endocrinol. Metab. Clin. N. Am. 2019, 48, 1–22. [Google Scholar] [CrossRef]
- Grogan, R.H.; Mitmaker, E.J.; Clark, O.H. The evolution of biomarkers in thyroid cancer-from mass screening to a personalized biosignature. Cancers 2010, 2, 885–912. [Google Scholar] [CrossRef] [Green Version]
- Feldkamp, J.; Führer, D.; Luster, M.; Musholt, T.J.; Spitzweg, C.; Schott, M. Fine Needle Aspiration in the Investigation of Thyroid Nodules. Dtsch. Aerzteblatt Online 2016, 113, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Moses, W.; Weng, J.; Sansano, I.; Peng, M.; Khanafshar, E.; Ljung, B.-M.; Duh, Q.-Y.; Clark, O.H.; Kebebew, E. Molecular Testing for Somatic Mutations Improves the Accuracy of Thyroid Fine-needle Aspiration Biopsy. World J. Surg. 2010, 34, 2589–2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macario, A.J.L.; Conway de Macario, E. Sick chaperones, cellular stress, and disease. N. Engl. J. Med. 2005, 353, 1489–1501. [Google Scholar] [CrossRef]
- Macario, A.J.L.; Conway de Macario, E. Chaperone proteins and Chaperonopathies. In Handbook of Stress, Volume 3, Stress Physiology, Biochemistry, and Pathology; Fink, G., Ed.; Elsevier/Academic Press: Cambridge, MA, USA, 2019; Chapter 12; pp. 135–152. [Google Scholar]
- Kocaturk, N.M.; Gozuacik, D. Crosstalk Between Mammalian Autophagy and the Ubiquitin-Proteasome System. Front. Cell Dev. Biol. 2018, 6, 128. [Google Scholar] [CrossRef]
- Tekirdag, K.; Cuervo, A.M. Chaperone-mediated autophagy and endosomal microautophagy: Jointed by a chaperone. J. Biol. Chem. 2018, 293, 5414–5424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, B.; Fares, M.A.; Lund, P.A. Chaperonin 60: A paradoxical, evolutionarily conserved protein family with multiple moonlighting functions. Biol. Rev. Camb. Philos. Soc. 2013, 88, 955–987. [Google Scholar] [CrossRef]
- Macario, A.J.L.; Conway de Macario, E.; Cappello, F. The Chaperonopathies. Diseases with Defective Molecular Chaperones; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2013. [Google Scholar]
- Calderwood, S.K.; Gong, J. Heat shock proteins promote cancer: It’s a protection racket. Trends Biochem. Sci. 2016, 41, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Jeffery, C.J. Protein moonlighting: What is it, and why is it important? Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20160523. [Google Scholar] [CrossRef] [PubMed]
- Saini, J.; Sharma, P.K. Clinical, Prognostic and therapeutic significance of heat shock proteins in cancer. Curr. Drug Targets 2018, 19, 1478–1490. [Google Scholar] [CrossRef] [PubMed]
- Milani, A.; Basirnejad, M.; Bolhassani, A. Heat-shock proteins in diagnosis and treatment: An overview of different biochemical and immunological functions. Immunotherapy 2019, 11, 215–239. [Google Scholar] [CrossRef]
- Yun, C.W.; Kim, H.J.; Lim, J.H.; Lee, S.H. Heat Shock Proteins: Agents of Cancer Development and Therapeutic Targets in Anti-Cancer Therapy. Cells 2019, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Cappello, F.; Marino Gammazza, A.; Palumbo Piccionello, A.; Campanella, C.; Pace, A.; Conway de Macario, E.; Macario, A.J.L. Hsp60 chaperonopathies and chaperonotherapy: Targets and agents. Expert Opin. Ther. Targets 2013, 18, 185–208. [Google Scholar] [CrossRef]
- Meng, Q.; Li, B.X.; Xiao, X. Toward developing chemical modulators of Hsp60 as potential therapeutics. Front. Mol. Biosci. 2018, 5, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Yang, S.; Vlantis, A.C.; Liu, S.Y.; Ng, E.K.; Chan, A.B.; Wu, J.; Du, J.; Wei, W.; Liu, X.; et al. Expression of antioxidant molecules and heat shock protein 27 in thyroid tumors. J. Cell. Biochem. 2016, 117, 2473–2481. [Google Scholar] [CrossRef]
- Caruso Bavisotto, C.; Cipolla, C.; Graceffa, G.; Barone, R.; Bucchieri, F.; Bulone, D.; Cabibi, D.; Campanella, C.; Marino Gammazza, A.; Pitruzzella, A.; et al. Immunomorphological pattern of molecular chaperones in normal and pathological thyroid tissues and circulating exosomes: potential use in clinics. Int. J. Mol. Sci. 2019, 20, 4496. [Google Scholar] [CrossRef] [Green Version]
- Pitruzzella, A.; Paladino, L.; Vitale, A.M.; Martorana, S.; Cipolla, C.; Graceffa, G.; Cabibi, D.; David, S.; Fucarino, A.; Bucchieri, F.; et al. Quantitative immunomorphological analysis of heat shock proteins in thyroid follicular adenoma and carcinoma tissues reveals their potential for differential diagnosis and points to a role in carcinogenesis. Appl. Sci. 2019, 9, 4324. [Google Scholar] [CrossRef] [Green Version]
- Starenki, D.; Hong, S.-K.; Lloyd, R.V.; Park, J.-I. Mortalin (GRP75/HSPA9) upregulation promotes survival and proliferation of medullary thyroid carcinoma cells. Oncogene 2014, 34, 4624–4634. [Google Scholar] [CrossRef] [Green Version]
- Starenki, D.; Sosonkina, N.; Hong, S.K.; Lloyd, R.V.; Park, J.I. Mortalin (GRP75/HSPA9) promotes survival and proliferation of thyroid carcinoma cells. Int. J. Mol. Sci. 2019, 20, 2069. [Google Scholar] [CrossRef] [Green Version]
- Soudry, E.; Shavit, S.S.; Hardy, B.; Morgenstern, S.; Hadar, T.; Feinmesser, R. Heat shock proteins HSP90, HSP70 and GRP78 expression in medullary thyroid carcinoma. Ann. Diagn. Pathol. 2017, 26, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.-J.; Qiu, Y.-B.; Jiang, R.; Xu, M.; Liao, L.-Y.; Chen, G.G.; Liu, Z.-M. Concomitant high expression of ERα36, GRP78 and GRP94 is associated with aggressive papillary thyroid cancer behavior. Cell. Oncol. 2018, 41, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Zhan, S.; Xia, W.; Huang, L.; Ge, W.; Wang, T. Proteomics study of serum exosomes from papillary thyroid cancer patients. Endocr. Relat. Cancer 2018, 25, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Garrido, C.; Brunet, M.; Didelot, C.; Zermati, Y.; Schmitt, E.; Kroemer, G. Heat shock proteins 27 and 70: Anti-apoptotic proteins with tumorigenic properties. Cell Cycle 2006, 5, 2592–2601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, X.-M.; Li, L.; Zhu, P.; Dai, Y.-J.; Zhao, T.-T.; Liao, L.-Y.; Chen, G.G.; Liu, Z.-M. Up-regulation of Hsp27 by ERα/Sp1 facilitates proliferation and confers resistance to apoptosis in human papillary thyroid cancer cells. Mol. Cell. Endocrinol. 2016, 431, 71–87. [Google Scholar] [CrossRef]
- Pandey, P.M.; Farber, R.; Nakazawa, A.; Kumar, S.; Bharti, A.C.; Nalin, C.M.; Weichselbaum, R.R.; Kufe, D.; Kharbanda, S. Hsp27 functions as a negative regulator of cytochrome c-dependent activation of procaspase-3. Oncogene 2000, 19, 1975–1981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voss, O.H.; Batra, S.; Kolattukudy, S.J.; Gonzalez-Mejia, M.E.; Smith, J.B.; Doseff, A.I. Binding of Caspase-3 Prodomain to Heat Shock Protein 27 Regulates Monocyte Apoptosis by Inhibiting Caspase-3 Proteolytic Activation. J. Biol. Chem. 2007, 282, 25088–25099. [Google Scholar] [CrossRef] [Green Version]
- Lettini, G.; Pietrafesa, M.; Lepore, S.; Maddalena, F.; Crispo, F.; Sgambato, A.; Esposito, F.; Landriscina, M. Heat shock proteins in thyroid malignancies: Potential therapeutic targets for poorly differentiated and anaplastic tumours? Mol. Cell. Endocrinol. 2020, 502, 110676. [Google Scholar] [CrossRef]
- Derwahl, M.; Nicula, D. Estrogen and its role in thyroid cancer. Endocr. Relat. Cancer 2014, 21, T273–T283. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, Q.; Guan, H.; Shi, B.; Ji, M.; Hou, P. ZNF677 Suppresses Akt Phosphorylation and Tumorigenesis in Thyroid Cancer. Cancer Res. 2018, 78, 5216–5228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jutabha, P.; Kanai, Y.; Hosoyamada, M.; Chairoungdua, A.; Kim, D.K.; Iribe, Y.; Babu, E.; Kim, J.Y.; Anzai, N.; Chatsudthipong, V.; et al. Identification of a Novel Voltage-driven Organic Anion Transporter Present at Apical Membrane of Renal Proximal Tubule. J. Biol. Chem. 2003, 278, 27930–27938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappello, F.; Czarnecka, A.M.; La Rocca, G.; Di Stefano, A.; Zummo, G.; Macario, A.J.L. Hsp60 and Hspl0 as antitumor molecular agents. Cancer Biol. Ther. 2007, 6, 487–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappello, F.; Bellafiore, M.; Palma, A.; Marciano, V.; Martorana, G.; Belfiore, P.; Martorana, A.; Farina, F.; Zummo, G.; Bucchieri, F. Expression of 60-kD heat shock proteinin creases during carcinogenesis in the uterine exocervix. Pathobiology 2002, 70, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Cappello, F.; Bellafiore, M.; Palma, A.; David, S.; Marcianò, V.; Bartolotta, T.; Sciumè, C.; Modica, G.; Farina, F.; Zummo, G.; et al. 60kDa chaperonin (HSP60) is over-expressed during colorectal carcinogenesis. Eur. J. Histochem. 2003, 47, 105–110. [Google Scholar] [CrossRef]
- Cappello, F.; David, S.; Rappa, F.; Bucchieri, F.; Marasà, L.; Bartolotta, T.E.; Farina, F.; Zummo, G. The expression of HSP60 and HSP10 in large bowel carcinomas with lymph node metastase. BMC Cancer 2005, 5, 139. [Google Scholar] [CrossRef] [Green Version]
- Caruso Bavisotto, C.; Alberti, G.; Vitale, A.M.; Paladino, L.; Campanella, C.; Rappa, F.; Gorska, M.; Conway de Macario, E.; Cappello, F.; Macario, A.J.L.; et al. Hsp60 post-translational modifications: Functional and pathological consequences. Front. Mol. Biosci. 2020, 7, 95. [Google Scholar] [CrossRef]
- Jolly, C.; Morimoto, R.I. Role of the heat shock response and molecular chaperones in oncogenesis and cell death. J. Natl. Cancer Inst. 2000, 92, 1564–1572. [Google Scholar] [CrossRef] [Green Version]
- Yaglom, J.A.; Wang, Y.; Li, A.; Li, Z.; Monti, S.; Alexandrov, I.; Lu, X.; Sherman, M.Y. Cancer cell responses to Hsp70 inhibitor JG-98: Comparison with Hsp90 inhibitors and finding synergistic drug combinations. Sci. Rep. 2018, 8, 3010. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Kang, J.G.; Kim, C.S.; Ihm, S.-H.; Choi, M.G.; Yoo, H.J.; Lee, S.J. Hsp70 inhibition potentiates radicicol-induced cell death in anaplastic thyroid carcinoma cells. Anticancer Res. 2014, 34, 4829–4837. [Google Scholar] [PubMed]
- Kiang, J.G.; Gist, I.D.; Tsokos, G.C. 17 beta-estradiol-induced increases in glucose-regulated protein 78kD and 94kD protect human breast cancer T47-D cells from thermal injury. Chin. J. Physiol. 1997, 40, 213–219. [Google Scholar] [PubMed]
- Zheng, H.-C.; Takahashi, H.; Li, X.-H.; Hara, T.; Masuda, S.; Guan, Y.-F.; Takano, Y. Overexpression of GRP78 and GRP94 are markers for aggressive behavior and poor prognosis in gastric carcinomas. Hum. Pathol. 2008, 39, 1042–1049. [Google Scholar] [CrossRef]
- Scriven, P.; Coulson, S.; Haines, R.; Balasubramanian, S.; Cross, S.; Wyld, L. Activation and clinical significance of the unfolded protein response in breast cancer. Br. J. Cancer 2009, 101, 1692–1698. [Google Scholar] [CrossRef] [Green Version]
- Delie, F.; Petignat, P.; Cohen, M. GRP78 Protein expression in ovarian cancer patients and perspectives for a drug-targeting approach. J. Oncol. 2012, 2012, 1–5. [Google Scholar] [CrossRef]
- Pearl, L.H.; Prodromou, C.; Workman, P. The Hsp90 molecular chaperone: An open and shut case for treatment. Biochem. J. 2008, 410, 439–453. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sun, W.; Dong, W.; Wang, Z.; Qin, Y.; Zhang, T.; Zhang, H. HSP90 inhibitor NVP-AUY922 induces cell apoptosis by disruption of the survivin in papillary thyroid carcinoma cells. Biochem. Biophys. Res. Commun. 2017, 487, 313–319. [Google Scholar] [CrossRef]
- Mo, J.-H.; Choi, I.J.; Jeong, W.-J.; Jeon, E.-H.; Ahn, S.-H. HIF-1α and HSP90: Target molecules selected from a tumorigenic papillary thyroid carcinoma cell line. Cancer Sci. 2012, 103, 464–471. [Google Scholar] [CrossRef]
- Wang, J.; Cui, S.; Zhang, X.; Wu, Y.; Tang, H. High expression of heat shock protein 90 is associated with tumor aggressiveness and poor prognosis in patients with advanced gastric cancer. PLoS ONE 2013, 8, e62876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Guo, S.; Li, Z.; Li, D.; Zhan, Q. High expression of HSP90 is associated with poor prognosis in patients with colorectal cancer. PeerJ 2019, 7, e7946. [Google Scholar] [CrossRef] [PubMed]
- Boltze, C.; Schneider-Stock, R.; Roessner, A.; Quednow, C.; Hoang-Vu, C. Function of HSP90 and p23 in the telomerase complex of thyroid tumors. Pathol. Res. Pract. 2003, 199, 573–579. [Google Scholar] [CrossRef]
- Holt, S.E.; Aisner, D.L.; Baur, J.; Tesmer, V.M.; Dy, M.; Ouellette, M.; Trager, J.B.; Morin, G.B.; Toft, D.O.; Shay, J.W.; et al. Functional requirement of p23 and Hsp90 in telomerase complexes. Genes Dev. 1999, 13, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Hoang-Vu, C.; Boltze, C.; Gimm, O.; Poremba, C.; Dockhorn-Dworniczak, B.; Köhrle, J.; Rath, F.-W.; Dralle, H. Expression of telomerase genes in thyroid carcinoma. Int. J. Oncol. 2002, 21, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Palladino, G.; Notarangelo, T.; Pannone, G.; Piscazzi, A.; Lamacchia, O.; Sisinni, L.; Spagnoletti, G.; Toti, P.; Santoro, A.; Storto, G.; et al. TRAP1 regulates cell cycle and apoptosis in thyroid carcinoma cells. Endocr. Relat. Cancer 2016, 23, 699–709. [Google Scholar] [CrossRef] [Green Version]
- Rasola, A.; Neckers, L.; Picard, D. Mitochondrial oxidative phosphorylation TRAP(1)ped in tumor cells. Trends Cell Biol. 2014, 24, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Braga-Basaria, M.; Hardy, E.; Gottfried, R.; Burman, K.D.; Saji, M.; Ringel, M.D. 17-Allylamino-17-Demethoxygeldanamycin activity against thyroid cancer cell lines correlates with heat shock protein 90 levels. J. Clin. Endocrinol. Metab. 2004, 89, 2982–2988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, P.T.; Subramanian, C.; Zhu, Q.; Zhang, H.; Zhao, H.; Gallagher, R.J.; Timmermann, B.N.; Blagg, B.S.J.; Cohen, M.S. Novel HSP90 inhibitors effectively target functions of thyroid cancer stem cell preventing migration and invasion. Surgery 2016, 159, 142–151. [Google Scholar] [CrossRef] [Green Version]
- Samadi, A.; Loo, P.; Mukerji, R.; O’Donnell, G.; Tong, X.; Timmermann, B.N.; Cohen, M.S. A novel HSP90 modulator with selective activity against thyroid cancers in vitro. Surgery 2009, 146, 1196–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campanella, C.; Bucchieri, F.; Ardizzone, N.M.; Marino Gammazza, A.; Montalbano, A.; Ribbene, A.; Di Felice, V.; Bellafiore, M.; David, S.; Rappa, F.; et al. Upon oxidative stress, the antiapoptotic Hsp60/procaspase-3 complex persists in mucoepidermoid carcinoma cells. Eur. J. Histochem. 2008, 52, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Campanella, C.; Bucchieri, F.; Merendino, A.M.; Fucarino, A.; Burgio, G.; Corona, D.F.; Barbieri, G.; David, S.; Farina, F.; Zummo, G.; et al. The odyssey of Hsp60 from tumor cells to other destinationsincludes plasma membrane-associated stages and Golgi and exosomal protein-trafficking modalities. PLoS ONE 2012, 7, e42008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campanella, C.; Rappa, F.; Sciumè, C.; Marino Gammazza, A.; Barone, R.; Bucchieri, F.; David, S.; Curcurù, G.; Caruso Bavisotto, C.; Pitruzzella, A.; et al. Heat shock protein 60 levels in tissue and circulating exosomes in human large bowel cancer before and after ablative surgery. Cancer 2015, 121, 3230–3239. [Google Scholar] [CrossRef] [PubMed]
- Santos, T.G.; Martins, V.R.; Hajj, G.N.M. Unconventional secretion of heat shock proteins in cancer. Int. J. Mol. Sci. 2017, 18, 946. [Google Scholar] [CrossRef] [Green Version]
- Shevtsov, M.; Balogi, Z.; Khachatryan, W.; Gao, H.; Vígh, L.; Multhoff, G. Membrane-associated heat shock proteins in oncology: from basic research to new theranostic targets. Cells 2020, 9, 1263. [Google Scholar] [CrossRef] [PubMed]
- Avdalyan, A.M.; Ivanov, A.A.; Lushnikova, E.L.; Molodykh, O.P.; Vikhlyanov, I.V. The Relationship of immunoexpression of Ki-67 and Hsp70 with clinical and morphological parameters and prognosis of papillary thyroid cancer. Bull. Exp. Biol. Med. 2020, 168, 688–693. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Zaborowski, M.P.; Balaj, L.; Breakefield, X.O.; Lai, C.P. Extracellular vesicles: Composition, biological relevance, and methods of study. Bioscience 2015, 65, 783–797. [Google Scholar] [CrossRef] [Green Version]
- Vader, P.; Breakefield, X.O.; Wood, M.J. Extracellular vesicles: Emerging targets for cancer therapy. Trends Mol. Med. 2014, 20, 385–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappa, G.; Puglisi, C.; Santos, M.F.; Forte, S.; Memeo, L.; Lorico, A. Extracellular vesicles from thyroid carcinoma: The new frontier of liquid biopsy. Int. J. Mol. Sci. 2019, 20, 1114. [Google Scholar] [CrossRef] [Green Version]
- Feng, K.; Ma, R.; Zhang, L.; Li, H.; Tang, Y.; Du, G.; Niu, D.; Yin, D. The role of exosomes in thyroid cancer and their potential clinical application. Front. Oncol. 2020, 10, 596132. [Google Scholar] [CrossRef]
- Gobbo, J.; Marcion, G.; Cordonnier, M.; Dias, A.M.M.; Pernet, N.; Hammann, A.; Richaud, S.; Mjahed, H.; Isambert, N.; Clausse, V.; et al. Restoring anticancer immune response by targeting tumor-derived exosomes with a HSP70 peptide aptamer. J. Natl. Cancer Inst. 2015, 108. [Google Scholar] [CrossRef]
- Li, X.; Wang, S.; Zhu, R.; Li, H.; Han, Q.; Zhao, R.C. Lung tumor exosomes induce a pro-inflammatory phenotype in mesenchymal stem cells via NFκB-TLR signaling pathway. J. Hematol. Oncol. 2016, 9, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stope, M.B.; Klinkmann, G.; Diesing, K.; Koensgen, D.; Burchardt, M.; Mustea, A. Heat shock protein HSP27 secretion by ovarian cancer cells is linked to intracellular expression levels, occurs independently of the endoplasmic reticulum pathway and HSP27′s phosphorylation status, and is mediated by exosome liberation. Dis. Mark. 2017, 2017, 1–12. [Google Scholar] [CrossRef]
- Caruso Bavisotto, C.; Graziano, F.; Rappa, F.; Marino Gammazza, A.; Logozzi, M.; Fais, S.; Maugeri, R.; Bucchieri, F.; Conway de Macario, E.; Macario, A.J.L.; et al. Exosomal chaperones and miRNAs in Gliomagenesis: State-of-Art and theranostics perspectives. Int. J. Mol. Sci. 2018, 19, 2626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyciszkiewicz, A.; Kalinowska-Łyszczarz, A.; Nowakowski, B.; Kaźmierczak, K.; Osztynowicz, K.; Michalak, S. Expression of small heat shock proteins in exosomes from patients with gynecologic cancers. Sci. Rep. 2019, 9, 9817. [Google Scholar] [CrossRef] [Green Version]
- Lorenc, T.; Klimczyk, K.; Michalczewska, I.; Słomka, M.; Kubiak-Tomaszewska, G.; Olejarz, W. Exosomes in prostate cancer diagnosis, prognosis and therapy. Int. J. Mol. Sci. 2020, 21, 2118. [Google Scholar] [CrossRef] [Green Version]
- Naryzhny, S.; Volnitskiy, A.; Kopylov, A.; Zorina, E.; Kamyshinsky, R.; Bairamukov, V.; Garaeva, L.; Shlikht, A.; Shtam, T. Proteome of glioblastoma-derived exosomes as a source of biomarkers. Biomedicine 2020, 8, 216. [Google Scholar] [CrossRef]
- Sherman, M.E.; Troester, M.A.; Hoadley, K.A.; Anderson, W.F. Morphological and molecular classification of human cancer. In Cancer Epidemiology and Prevention; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Yasuda, Y.; Tokunaga, K.; Koga, T.; Sakamoto, C.; Goldberg, I.G.; Saitoh, N.; Nakao, M. Computational analysis of morphological and molecular features in gastric cancer tissues. Cancer Med. 2020, 9, 2223–2234. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Chaperone | Tumor | Quantitative Level | Reference | |
|---|---|---|---|---|
| Immunohistochemistry | Hsp27 (HSPB1) | FA 1 | Increased | [20] |
| FTC | Increased | [20] | ||
| PTC | Increased | [21] | ||
| Hsp60 | FTC | Increased | [22] | |
| PTC | Increased | [21] | ||
| Hsp70 | FTC | Increased | [22] | |
| Mortalin (HSPA9; GRP75) | ATC | Increased | [23,24] | |
| FTC | Increased | |||
| MTC | Increased | |||
| PTC | Increased | |||
| GRP78 | MTC | Increased | [25] | |
| PTC | Increased | [26] | ||
| GRP94 | PTC | Increased | [26] | |
| Hsp90 | FTC | Increased | [22] | |
| PTC | Increased | [21] | ||
| Biochemistry; immunochemistry | Hsp27 | ATC | Increased | [20] |
| FTC | Increased | |||
| PTC | Increased | |||
| Mortalin (HSPA9; GRP75) | MTC | Increased | [23] | |
| GRP78 | PTC | Increased | [26] | |
| GRP94 | PTC | Increased | [26] | |
| Exosomes | Hsp27 | PTC | Increased | [21,27] |
| Hsp60 | PTC | Increased | [21] | |
| Hsp90 | PTC | Increased | [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paladino, L.; Vitale, A.M.; Santonocito, R.; Pitruzzella, A.; Cipolla, C.; Graceffa, G.; Bucchieri, F.; Conway de Macario, E.; Macario, A.J.L.; Rappa, F. Molecular Chaperones and Thyroid Cancer. Int. J. Mol. Sci. 2021, 22, 4196. https://doi.org/10.3390/ijms22084196
Paladino L, Vitale AM, Santonocito R, Pitruzzella A, Cipolla C, Graceffa G, Bucchieri F, Conway de Macario E, Macario AJL, Rappa F. Molecular Chaperones and Thyroid Cancer. International Journal of Molecular Sciences. 2021; 22(8):4196. https://doi.org/10.3390/ijms22084196
Chicago/Turabian StylePaladino, Letizia, Alessandra Maria Vitale, Radha Santonocito, Alessandro Pitruzzella, Calogero Cipolla, Giuseppa Graceffa, Fabio Bucchieri, Everly Conway de Macario, Alberto J. L. Macario, and Francesca Rappa. 2021. "Molecular Chaperones and Thyroid Cancer" International Journal of Molecular Sciences 22, no. 8: 4196. https://doi.org/10.3390/ijms22084196
APA StylePaladino, L., Vitale, A. M., Santonocito, R., Pitruzzella, A., Cipolla, C., Graceffa, G., Bucchieri, F., Conway de Macario, E., Macario, A. J. L., & Rappa, F. (2021). Molecular Chaperones and Thyroid Cancer. International Journal of Molecular Sciences, 22(8), 4196. https://doi.org/10.3390/ijms22084196