
Detection of Antimicrobial Peptides in Stratum Corneum by Mass Spectrometry

 , ,
, , 

Abstract
:1. Introduction
2. Results
2.1. Method Development and Optimization for the Examination of AMPs in the Stratum Corneum
2.1.1. Optimization of Sample Collection
2.1.2. Design and Optimization of PRM-Based Mass Spectrometry Method
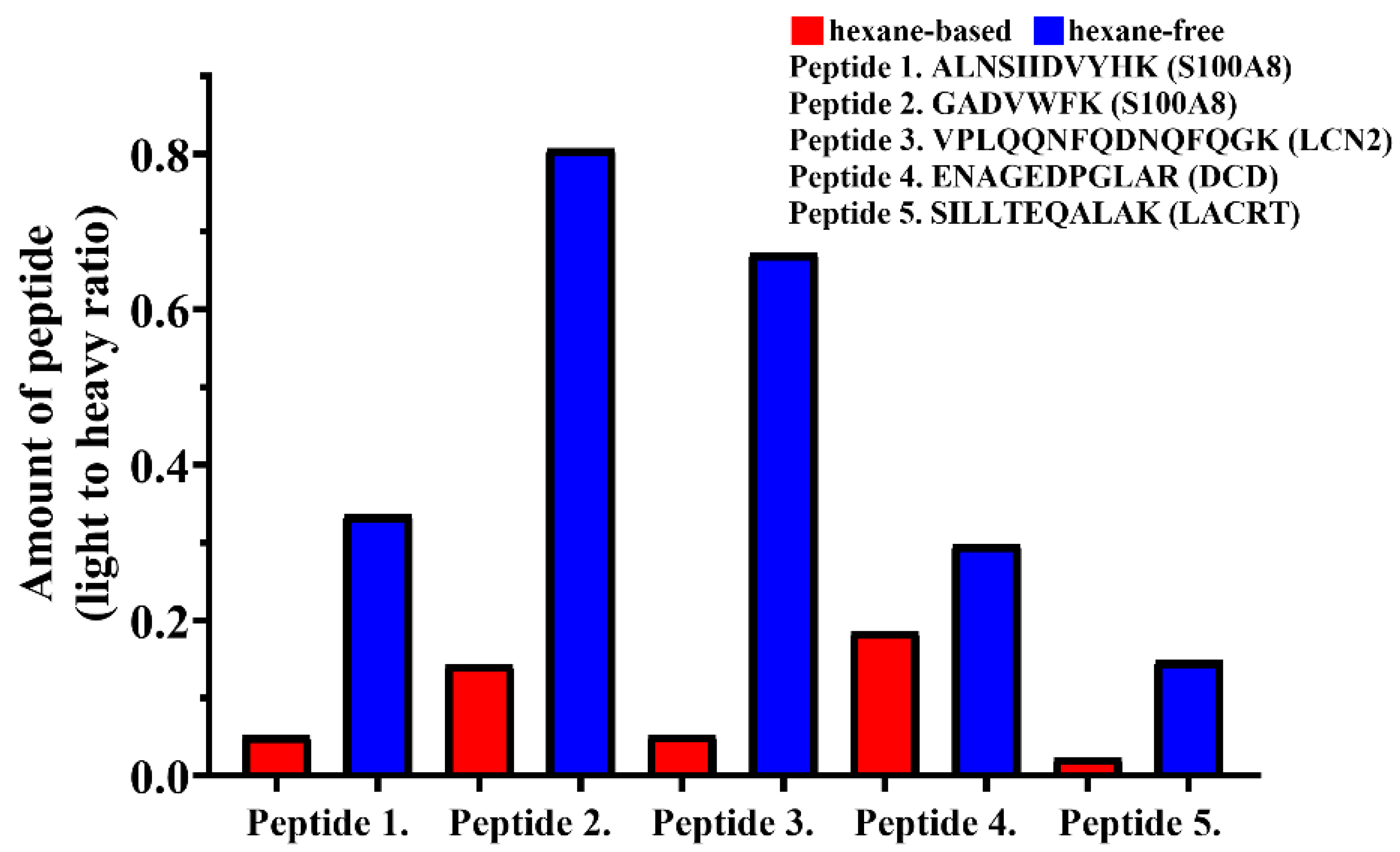
2.1.3. Optimization of Protein Elution
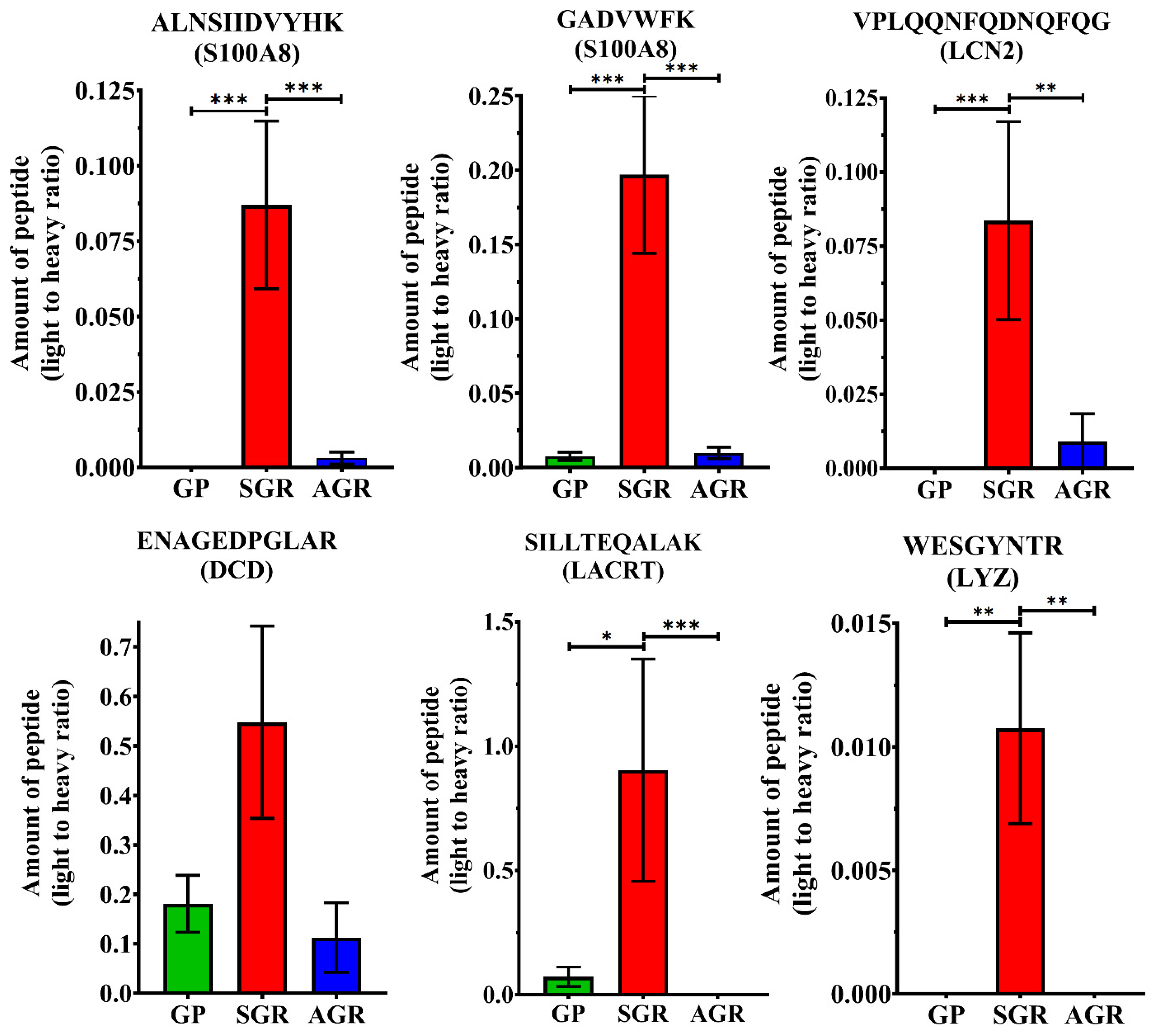
2.2. Comparative Analysis of AMP Levels of Different Healthy Skin Regions
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Sample Collection
4.3. Measurement of Protein Concentration
4.4. Protein Elution from D-Squame Discs
4.5. Protein Digestion
4.6. PRM Assay Design
4.7. Mass Spectrometry Experiment
4.8. Data Evaluation and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nguyen, H.L.T.; Trujillo-Paez, J.V.; Umehara, Y.; Yue, H.; Peng, G.; Kiatsurayanon, C.; Chieosilapatham, P.; Song, P.; Okumura, K.; Ogawa, H.; et al. Role of Antimicrobial Peptides in Skin Barrier Repair in Individuals with Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 7607. [Google Scholar] [CrossRef]
- Béke, G.; Dajnoki, Z.; Kapitány, A.; Gáspár, A.; Medgyesi, B.; Poliska, S.; Hendrik, Z.; Péter, Z.; Torocsik, D.; Bíró, T.; et al. Immunotopographical differences of human skin. Front. Immunol. 2018, 9, 424. [Google Scholar] [CrossRef] [Green Version]
- Hänel, K.H.; Cornelissen, C.; Lüscher, B.; Baron, J.M. Cytokines and the skin barrier. Int. J. Mol. Sci. 2013, 14, 6720–6745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elias, P.M.; Wakefield, J.S. Skin barrier function. Nutr. Healthy Ski. Strateg. Clin. Cosmet. Pract. 2011, 8, 35–48. [Google Scholar] [CrossRef]
- Borkowski, A.W.; Gallo, R.L. The coordinated response of the physical and antimicrobial peptide barriers of the skin. J. Investig. Dermatol. 2011, 131, 285–287. [Google Scholar] [CrossRef] [Green Version]
- Chan, A.; Godoy-Gijon, E.; Nuno-Gonzalez, A.; Crumrine, D.; Hupe, M.; Choi, E.-H.; Gruber, R.; Williams, M.L.; Choate, K.; Fleckman, P.H.; et al. Cellular basis of secondary infections and impaired desquamation in certain inherited ichthyoses. JAMA Dermatol. 2015, 151, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Niyonsaba, F.; Kiatsurayanon, C.; Chieosilapatham, P.; Ogawa, H. Friends or Foes? Host defense (antimicrobial) peptides and proteins in human skin diseases. Exp. Dermatol. 2017, 26, 989–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaleja, P.; Emmert, H.; Gerstel, U.; Weidinger, S.; Tholey, A. Evaluation and improvement of protein extraction methods for analysis of skin proteome by noninvasive tape stripping. J. Proteom. 2020, 217. [Google Scholar] [CrossRef] [PubMed]
- Bardan, A.; Nizet, V.; Gallo, R.L. Antimicrobial peptides and the skin. Expert Opin. Biol. Ther. 2004, 4, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Dinulos, J.G.H.; Mentele, L.; Fredericks, L.P.; Dale, B.A.; Darmstadt, G.L. Keratinocyte expression of human β defensin 2 following bacterial infection: Role in cutaneous host defense. Clin. Diagn. Lab. Immunol. 2003, 10, 161–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalló, G.; Emri, M.; Varga, Z.; Ujhelyi, B.; Tőzsér, J.; Csutak, A.; Csősz, É. Changes in the chemical barrier composition of tears in Alzheimer’s disease reveal potential tear diagnostic biomarkers. PLoS ONE 2016, 11, e0158000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, I.; Pivarcsi, A.; Kis, K.; Koreck, A.; Bodai, L.; McDowell, A.; Seltmann, H.; Patrick, S.; Zouboulis, C.C.; Kemény, L.; et al. Propionibacterium acnes and lipopolysaccharide induce the expression of antimicrobial peptides and proinflammatory cytokines/chemokines in human sebocytes. Microbes Infect. 2006, 8, 2195–2205. [Google Scholar] [CrossRef] [PubMed]
- Schauber, J.; Gallo, R.L. Antimicrobial peptides and the skin immune defense system. J. Allergy Clin. Immunol. 2009, 124, 13–18. [Google Scholar] [CrossRef]
- Yamasaki, K.; Di Nardo, A.; Bardan, A.; Murakami, M.; Ohtake, T.; Coda, A.; Dorschner, R.A.; Bonnart, C.; Descargues, P.; Hovnanian, A.; et al. Increased serine protease activity and cathelicidin promotes skin inflammation in rosacea. Nat. Med. 2007, 13, 975–980. [Google Scholar] [CrossRef]
- Kilmer, P.D. Review Article: Review Article. J. Theory Pract. Crit. 2010, 11, 369–373. [Google Scholar] [CrossRef]
- Csősz, É.; Deák, E.; Kalló, G.; Csutak, A.; Tőzsér, J. Diabetic retinopathy: Proteomic approaches to help the differential diagnosis and to understand the underlying molecular mechanisms. J. Proteom. 2017, 150, 351–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csosz, É.; Márton, I.J.; Horváth, J.; Lábiscsák, P.; Márkus, B.; Dezső, B.; Szabó, A.; Tar, I.; Piffkó, J.; Jakus, P.; et al. Proteomics investigation of OSCC-specific salivary biomarkers in a Hungarian population highlights the importance of identification of population-tailored biomarkers. PLoS ONE 2017, 12, e0177282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csősz, É.; Kalló, G.; Márkus, B.; Deák, E.; Csutak, A.; Tőzsér, J. Quantitative body fluid proteomics in medicine—A focus on minimal invasiveness. J. Proteom. 2017, 153, 30–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csősz, É.; Márkus, B.; Darula, Z.; Medzihradszky, K.F.; Nemes, J.; Szabó, E.; Tőzsér, J.; Kiss, C.; Márton, I.; Kalló, G.; et al. Salivary proteome profiling of oral squamous cell carcinoma in a Hungarian population. FEBS Open Bio 2018, 8, 556–569. [Google Scholar] [CrossRef] [Green Version]
- Rieg, S.; Garbe, C.; Sauer, B.; Kalbacher, H.; Schittek, B. Dermcidin is constitutively produced by eccrine sweat glands and is not induced in epidermal cells under inflammatory skin conditions. Br. J. Dermatol. 2004, 151, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Ong, P.Y.; Ohtake, T.; Brandt, C.; Strickland, I.; Boguniewicz, M.; Ganz, T.; Gallo, R.L.; Leung, D.Y.M. Endogenous antimicrobial peptides and skin infections in atopic dermatitis. N. Engl. J. Med. 2002, 347, 1151–1160. [Google Scholar] [CrossRef] [Green Version]
- Rieg, S.; Steffen, H.; Seeber, S.; Humeny, A.; Kalbacher, H.; Dietz, K.; Garbe, C.; Schittek, B. Deficiency of Dermcidin-Derived Antimicrobial Peptides in Sweat of Patients with Atopic Dermatitis Correlates with an Impaired Innate Defense of Human Skin In Vivo. J. Immunol. 2005, 174, 8003–8010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttman-Yassky, E.; Diaz, A.; Pavel, A.B.; Fernandes, M.; Lefferdink, R.; Erickson, T.; Canter, T.; Rangel, S.; Peng, X.; Estrada, Y.; et al. Use of Tape Strips to Detect Immune and Barrier Abnormalities in the Skin of Children with Early-Onset Atopic Dermatitis. JAMA Dermatol. 2019, 155, 1358–1370. [Google Scholar] [CrossRef]
- Dajnoki, Z.; Béke, G.; Kapitány, A.; Mócsai, G.; Gáspár, K.; Rühl, R.; Hendrik, Z.; Juhász, I.; Zouboulis, C.C.; Bácsi, A.; et al. Sebaceous Gland-Rich Skin Is Characterized by TSLP Expression and Distinct Immune Surveillance Which Is Disturbed in Rosacea. J. Investig. Dermatol. 2017, 137, 1114–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenei, A.; Dajnoki, Z.; Medgyesi, B.; Gáspár, K.; Béke, G.; Kinyó, Á.; Méhes, G.; Hendrik, Z.; Dinya, T.; Törőcsik, D.; et al. Apocrine Gland–Rich Skin Has a Non-Inflammatory IL-17–Related Immune Milieu, that Turns to Inflammatory IL-17–Mediated Disease in Hidradenitis Suppurativa. J. Investig. Dermatol. 2019, 139, 964–968. [Google Scholar] [CrossRef] [Green Version]
- De Jongh, G.J.; Zeeuwen, P.L.; Kucharekova, M.; Pfundt, R.; van der Valk, P.G.; Blokx, W.; Dogan, A.; Hiemstra, P.S.; van de Kerkhof, P.C.; Schalkwijk, J. High expression levels of keratinocyte antimicrobial proteins in psoriasis compared with atopic dermatitis. J. Investig. Dermatol. 2005, 125, 1163–1173. [Google Scholar] [CrossRef] [PubMed]
- Clausen, M.L.; Slotved, H.C.; Krogfelt, K.A.; Agner, T. Measurements of AMPs in stratum corneum of atopic dermatitis and healthy skin-tape stripping technique. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sano, Y.; Masuda, K.; Tamagawa-Mineoka, R.; Matsunaka, H.; Murakami, Y.; Yamashita, R.; Morita, E.; Katoh, N. Thymic stromal lymphopoietin expression is increased in the horny layer of patients with atopic dermatitis. Clin. Exp. Immunol. 2013, 171, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Matsushima, K.; Miyamoto, K.; Oe, T. Mass spectrometry data from proteomic analysis of human skin keratins after exposure to UV radiation. Data Br. 2016, 7, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Clausen, M.L.; Slotved, H.C.; Krogfelt, K.A.; Agner, T. Tape Stripping Technique for Stratum Corneum Protein Analysis. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lademann, J.; Jacobi, U.; Surber, C.; Weigmann, H.J.; Fluhr, J.W. The tape stripping procedure-evaluation of some critical parameters. Eur. J. Pharm. Biopharm. 2009, 72, 317–323. [Google Scholar] [CrossRef]
- Escobar-Chávez, J.J.; Merino-Sanjuán, V.; López-Cervantes, M.; Urban-Morlan, Z.; Piñón-Segundo, E.; Quintanar-Guerrero, D.; Ganem-Quintanar, A. The tape-stripping technique as a method for drug quantification in skin. J. Pharm. Pharm. Sci. 2008, 11, 104–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiba, T.; Nakahara, T.; Kohda, F.; Ichiki, T.; Manabe, M.; Furue, M. Measurement of trihydroxy-linoleic acids in stratum corneum by tape-stripping: Possible biomarker of barrier function in atopic dermatitis. PLoS ONE 2019, 14, e0210013. [Google Scholar] [CrossRef] [PubMed]
- Koppes, S.A.; Brans, R.; Hadzavdic, S.L.; Frings-Dresen, M.H.W.; Rustemeyer, T.; Kezic, S. Stratum corneum tape stripping: Monitoring of inflammatory mediators in atopic dermatitis patients using topical therapy. Int. Arch. Allergy Immunol. 2016, 170, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Olesen, C.M.; Fuchs, C.S.K.; Philipsen, P.A.; Hædersdal, M.; Agner, T.; Clausen, M.L. Advancement through epidermis using tape stripping technique and Reflectance Confocal Microscopy. Sci. Rep. 2019, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bourmaud, A.; Gallien, S.; Domon, B. Parallel reaction monitoring using quadrupole-Orbitrap mass spectrometer: Principle and applications. Proteomics 2016, 16, 2146–2159. [Google Scholar] [CrossRef]
- Peterson, A.C.; Russell, J.D.; Bailey, D.J.; Westphall, M.S.; Coon, J.J. Parallel reaction monitoring for high resolution and high mass accuracy quantitative, targeted proteomics. Mol. Cell. Proteom. 2012, 11, 1475–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mócsai, G.; Gáspár, K.; Dajnoki, Z.; Tóth, B.; Gyimesi, E.; Bíró, T.; Maródi, L.; Szegedi, A. Investigation of Skin Barrier Functions and Allergic Sensitization in Patients with Hyper-IgE Syndrome. J. Clin. Immunol. 2015, 35, 681–688. [Google Scholar] [CrossRef] [Green Version]
- Lange, V.; Picotti, P.; Domon, B.; Aebersold, R. Selected reaction monitoring for quantitative proteomics: A tutorial. Mol. Syst. Biol. 2008, 4. [Google Scholar] [CrossRef] [PubMed]
- Hulshof, L.; Hack, D.P.; Hasnoe, Q.C.J.; Dontje, B.; Jakasa, I.; Riethmüller, C.; McLean, W.H.I.; van Aalderen, W.M.C.; Van’t Land, B.; Kezic, S.; et al. A minimally invasive tool to study immune response and skin barrier in children with atopic dermatitis. Br. J. Dermatol. 2019, 180, 621–630. [Google Scholar] [CrossRef]
- Murakami, M.; Ohtake, T.; Dorschner, R.A.; Schittek, B.; Garbe, C.; Gallo, R.L. Cathelicidin anti-microbial peptide expression in sweat, an innate defense system for the skin. J. Investig. Dermatol. 2002, 119, 1090–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, K.; Gallo, R.L. Antimicrobial peptides in human skin disease. Eur. J. Dermatol. 2008, 18, 11–21. [Google Scholar] [CrossRef]
- Prens, E.; Deckers, I. Pathophysiology of hidradenitis suppurativa: An update. J. Am. Acad. Dermatol. 2015, 73, S8–S11. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, M.; Schauber, J.; Leyden, J.J. New insights into rosacea pathophysiology: A review of recent findings. J. Am. Acad. Dermatol. 2013, 69, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Schröder, J.M.; Harder, J. Antimicrobial skin peptides and proteins. Cell. Mol. Life Sci. 2006, 63, 469–486. [Google Scholar] [CrossRef] [PubMed]
- UniProt. Available online: www.uni-prot.org (accessed on 20 June 2020).
- ExPaSy. Available online: https://web.expasy.org/peptide_cutter/ (accessed on 20 June 2020).
- BLASTp. Available online: https://blast.ncbi.nlm.nih.gov (accessed on 20 June 2020).
- Sharma, V.; Eckels, J.; Schilling, B.; Ludwig, C.; Jaffe, J.D.; MacCoss, M.J.; MacLean, B. Panorama public: A public repository for quantitative data sets processed in skyline. Mol. Cell. Proteom. 2018, 17, 1239–1244. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Protein Name | UniProt ID | Protein-Specific Unique Peptide | Precursor Ion (m/z) |
|---|---|---|---|---|
| S100A8 | calcium-binding protein A8 | P05109 | ALNSIIDVYHK | 424.903 |
| ALNSIIDVYHK * | 427.574 | |||
| S100A8 | calcium-binding protein A8 | P05109 | GADVWFK | 411.71 |
| GADVWFK * | 415.718 | |||
| S100A9 | calcium-binding protein A9 | P06702 | DLYNFLK | 456.744 |
| DLYNFLK * | 460.752 | |||
| LL-37 | cathelicidin | P49913 | LLGDFFR | 434.239 |
| LLGDFFR * | 439.243 | |||
| DCD | dermcidin | P81605 | ENAGEDPGLAR | 564.767 |
| ENAGEDPGLAR * | 569.771 | |||
| hBD1 | human beta-defensin 1 | P60022 | IQGTCYR | 449.216 |
| IQGTCYR * | 454.22 | |||
| hBD2 | human beta-defensin 2 | O15263 | GIGDPVTCLK | 530.278 |
| GIGDPVTCLK * | 534.285 | |||
| hBD3 | human beta-defensin 3 | P81534 | GIINTLQK | 443.771 |
| GIINTLQK * | 447.778 | |||
| hBD4 | human beta-defensin 4 | Q8WTQ1 | ICGYGTAR | 449.216 |
| ICGYGTAR * | 454.22 | |||
| LACRT | lacritin | Q9GZZ8 | QELNPLK | 421.242 |
| QELNPLK * | 425.249 | |||
| LACRT | lacritin | Q9GZZ8 | SILLTEQALAK | 593.855 |
| SILLTEQALAK * | 597.862 | |||
| LCN2 | lipocalin-2 | P80188 | VPLQQNFQDNQFQGK | 895.944 |
| VPLQQNFQDNQFQGK * | 899.951 | |||
| LCN2 | lipocalin-2 | P80188 | SYNVTSVLFR | 593.316 |
| SYNVTSVLFR * | 598.32 | |||
| LCN2 | lipocalin-2 | P80188 | TFVPGCQPGEFTLGNIK | 932.966 |
| TFVPGCQPGEFTLGNIK * | 936.973 | |||
| LYZ | lysozyme | P61626 | GISLANWMCLAK | 682.346 |
| GISLANWMCLAK * | 686.353 | |||
| LYZ | lysozyme | P61626 | WESGYNTR | 506.727 |
| WESGYNTR * | 511.731 | |||
| S100A7 | psoriasin | P31151 | SIIGMIDMFHK | 646.33 |
| SIIGMIDMFHK * | 650.337 | |||
| TSLP | thymic stromal lymphopoietin | Q969D9 | CLEQVSQLQGLWR | 808.914 |
| CLEQVSQLQGLWR * | 813.918 |
| Protein | Peptide | Healthy Skin | Inflamed Skin |
|---|---|---|---|
| S100A9 | DLYNFLK | − | + |
| LL-37 | LLGDFFR | − | + |
| LCN2 | TFVPGCQPGEFTLGNIK | − | + |
| S100A8 | ALNSIIDVYHK | + | + |
| S100A8 | GADVWFK | + | + |
| LCN2 | VPLQQNFQDNQFQGK | + | + |
| DCD | ENAGEDPGLAR | + | + |
| LACRT | SILLTEQALAK | + | + |
| LYZ | WESGYNTR | + | + |
| S100A7 | SIIGMIDMFHK | − | − |
| hBD1 | IQGTCYR | − | − |
| hBD2 | GIGDPVTCLK | − | − |
| hBD3 | GIINTLQK | − | − |
| hBD4 | ICGYGTAR | − | − |
| LCN2 | SYNVTSVLFR | − | − |
| TSLP | CLEQVSQLQGLWR | − | − |
| LACRT | QELNPLK | − | − |
| LYZ | GISLANWMCLAK | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jenei, A.; Kalló, G.; Dajnoki, Z.; Gáspár, K.; Szegedi, A.; Kapitány, A.; Csősz, É. Detection of Antimicrobial Peptides in Stratum Corneum by Mass Spectrometry. Int. J. Mol. Sci. 2021, 22, 4233. https://doi.org/10.3390/ijms22084233
Jenei A, Kalló G, Dajnoki Z, Gáspár K, Szegedi A, Kapitány A, Csősz É. Detection of Antimicrobial Peptides in Stratum Corneum by Mass Spectrometry. International Journal of Molecular Sciences. 2021; 22(8):4233. https://doi.org/10.3390/ijms22084233
Chicago/Turabian StyleJenei, Adrienn, Gergő Kalló, Zsolt Dajnoki, Krisztián Gáspár, Andrea Szegedi, Anikó Kapitány, and Éva Csősz. 2021. "Detection of Antimicrobial Peptides in Stratum Corneum by Mass Spectrometry" International Journal of Molecular Sciences 22, no. 8: 4233. https://doi.org/10.3390/ijms22084233
APA StyleJenei, A., Kalló, G., Dajnoki, Z., Gáspár, K., Szegedi, A., Kapitány, A., & Csősz, É. (2021). Detection of Antimicrobial Peptides in Stratum Corneum by Mass Spectrometry. International Journal of Molecular Sciences, 22(8), 4233. https://doi.org/10.3390/ijms22084233