
Increased Osteocyte Lacunae Density in the Hypermineralized Bone Matrix of Children with Osteogenesis Imperfecta Type I

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
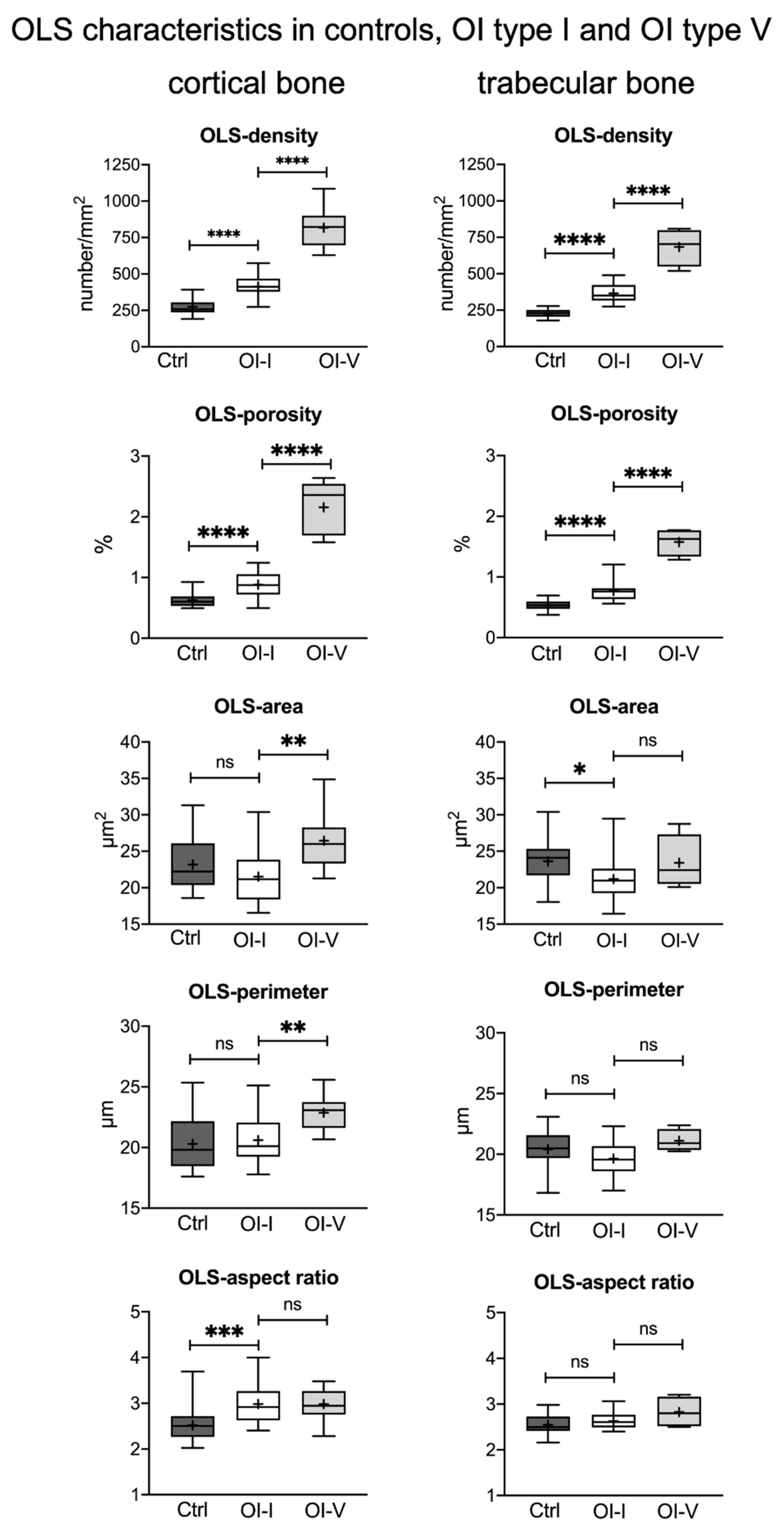
2.1. Osteocyte Lacunae Density but Not Size Is Increased in OI Type I Bone Compared to Healthy Controls
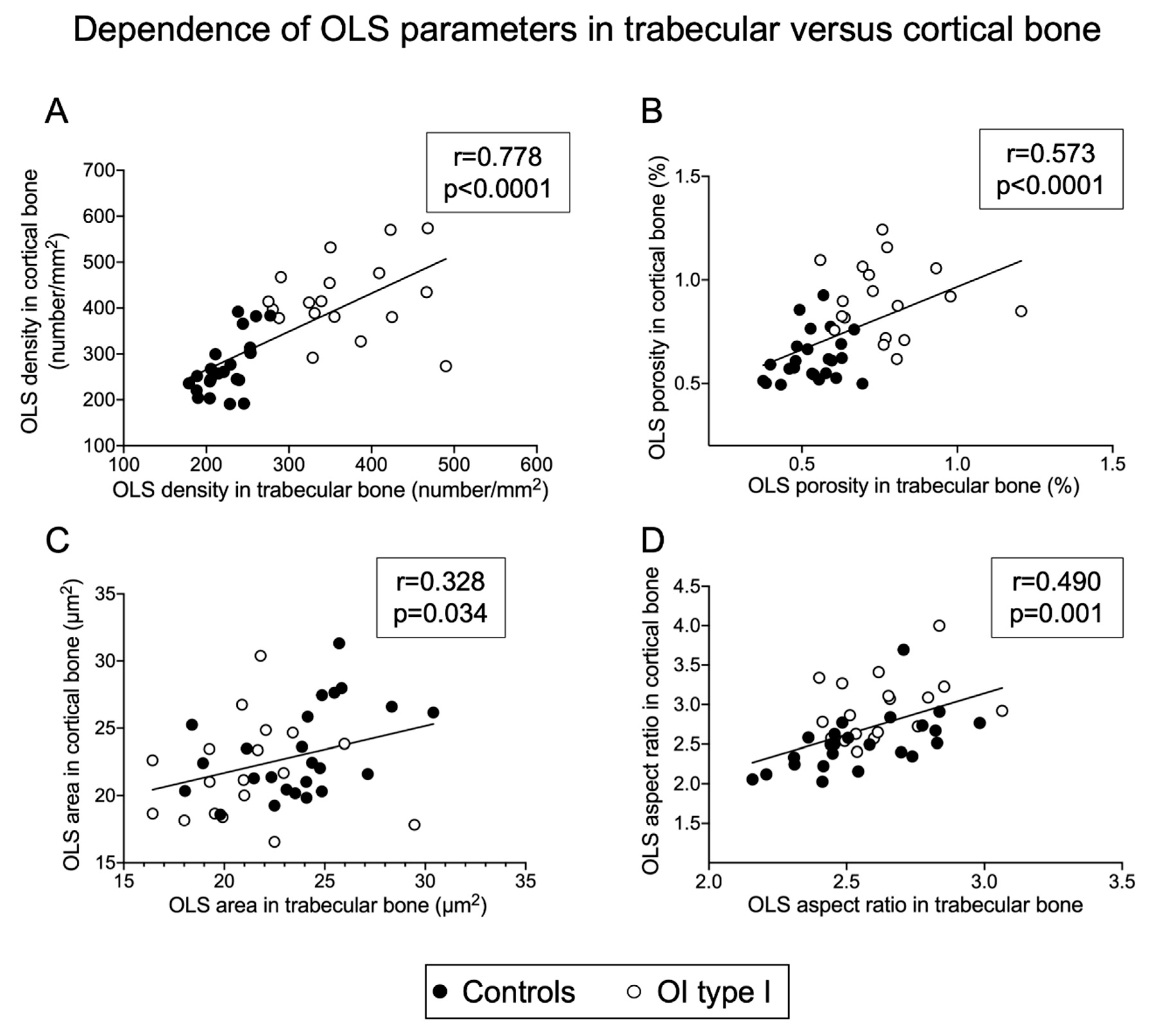
2.2. Osteocyte Lacunae Number, Size and Shape Are Correlated between Trabecular and Cortical Bone
2.3. Osteocyte Lacunar Differences between Qualitative and Quantitative Mutations
2.4. Osteocyte Lacunae Density in OI Type I Is Markedly Lower Than in OI Type V
2.5. Association of Age with Osteocyte Lacunae Characteristics
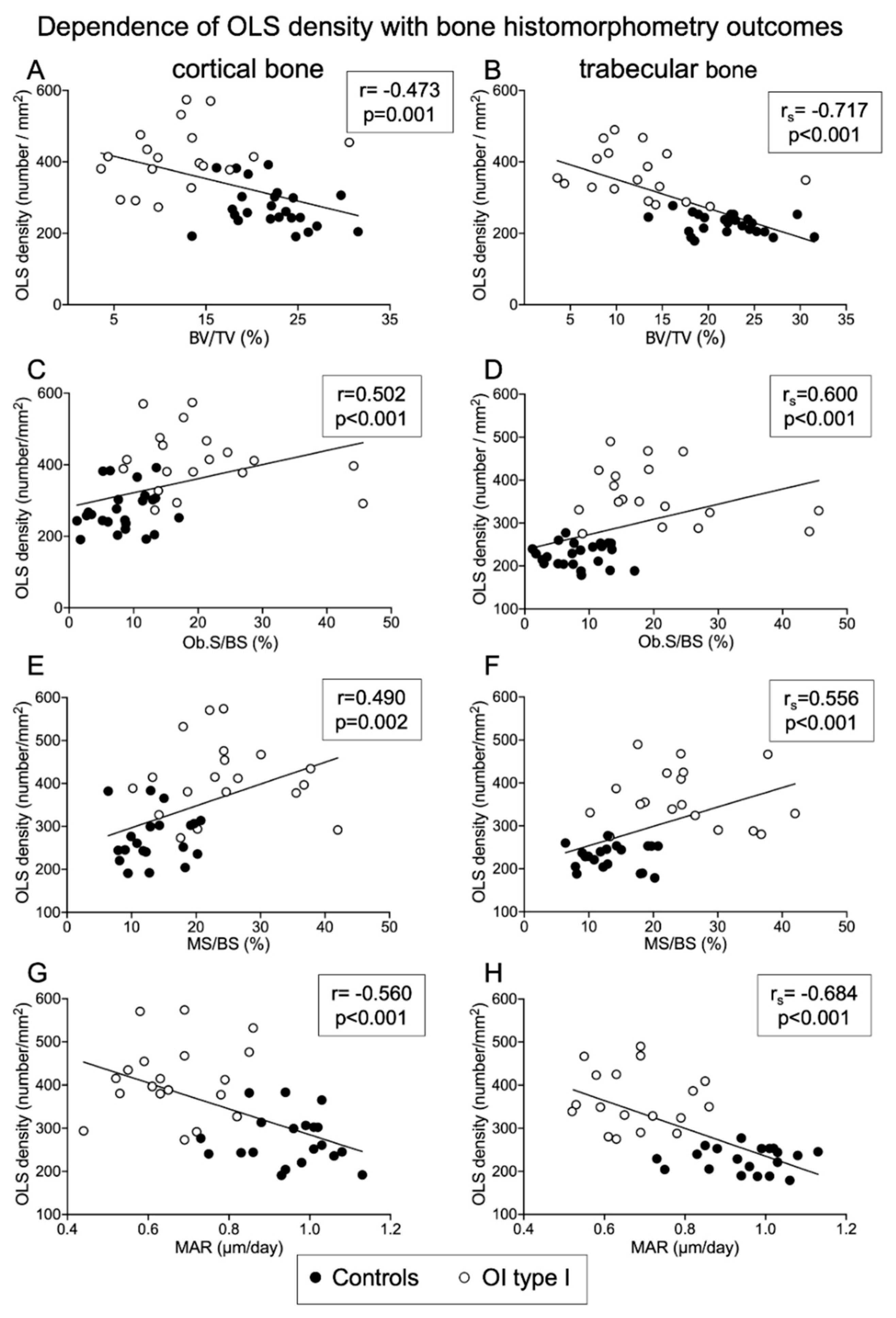
2.6. Relationship between Osteocyte Lacunae Characteristics and Bone Histomorphometry Outcomes
2.6.1. Correlations of OLS Density and Porosity with Histomorphometric Parameters
Negative Correlation of OLS Density and OLS Porosity with Structural Histomorphometric Parameters
Negative Correlation of OLS Density and OLS Porosity with Indices of Osteoblast Function
Positive Correlation of OLS Density and OLS Porosity with Surface-Based Bone Formation Indices
2.6.2. Correlation of OLS Area and OLS Perimeter with Histomorphometric Parameters
2.7. Bone Mineralization Density Distribution (BMDD) in OI Type I and Controls
3. Discussion
- First, the OLS density is increased in OI both in cortical and trabecular bone. This means that the total amount of bone matrix per osteocyte is reduced in OI.
- Second, the increased OLS density correlates with increased surface-based bone formation parameters previously obtained by histomorphometry in the same samples [19,43,44]. This supports previous findings that the reduced matrix production rate of osteoblasts in OI is partially compensated by an increased number of active osteoblasts [19].
- Third, the OLS aspect ratio in cortical bone is increased in OI. Given that osteocytes typically align with the collagen direction, this surprisingly suggests the presence of more primary lamellar tissue in the cortex of OI type I bone.
- The final observation is that the OLS area (i.e., the mean lacunar size) positively correlates with osteoid thickness and mineral apposition rate, measured by histomorphometry. This correlation is not obvious to interpret, but we may speculate that the enhanced mineral deposition activity when osteoid is to be mineralized leads to an increase in OLS area, perhaps to provide additional mineral through osteocytic osteolysis.
4. Patients and Methods
4.1. Study Cohort
4.2. Sample Preparation for qBEI Measurements
4.3. Quantitative Backscattered Electron Imaging (qBEI)
4.4. Osteocyte Lacunae Sections Analysis
- OLS density (OLS number/mm2): the number of OLS/(mineralized bone matrix area + OLS total area).
- OLS porosity: (%) OLS total area/(mineralized bone matrix area + OLS total area).
- OLS area (µm2): mean value of the OLS areas per sample (total OLS area divided by OLS number).
- OLS perimeter (µm): mean value of the OLS perimeters per sample.
- OLS aspect ratio: mean value of the OLS aspect ratio per sample. The OLS aspect ratio is a measure for the shape of the OLS. It is given by the ratio of the long to the short half-axis of a fitted ellipse to the section. A value of 1 indicates a perfect circle, while increasing values indicate an increasing elongated shape. OLS with aspect ratio values >10 were excluded from the analysis.
4.5. Bone Mineralization Density Distribution (BMDD)
- CaMean: the average calcium concentration (weighted mean);
- CaPeak: the most frequently occurring calcium concentration (the position of the peak of the BMDD);
- CaWidth: the width of the BMDD distribution (full-width at half-maximum), reflecting the heterogeneity in matrix mineralization;
- CaLow: the percentage of bone material mineralized below 18.20 weight %Ca, which corresponds to the 5th percentile of the reference BMDD in adult trabecular bone;
- CaHigh: the percentage of bone matrix having a mineral content above 26.86 weight %Ca, corresponding to the 95th percentile of the reference BMDD in adult trabecular bone.
4.6. Bone Histomorphometry
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franz-Odendaal, T.A.; Hall, B.K.; Witten, P.E. Buried alive: How osteoblasts become osteocytes. Dev. Dyn. 2006, 235, 176–190. [Google Scholar] [CrossRef]
- Schaffler, M.B.; Cheung, W.Y.; Majeska, R.; Kennedy, O. Osteocytes: Master orchestrators of bone. Calcif. Tissue Int. 2014, 94, 5–24. [Google Scholar] [CrossRef] [Green Version]
- Dallas, S.L.; Prideaux, M.; Bonewald, L.F. The osteocyte: An endocrine cell... and more. Endocr. Rev. 2013, 34, 658–690. [Google Scholar] [CrossRef] [Green Version]
- Robling, A.G.; Bonewald, L.F. The Osteocyte: New Insights. Annu. Rev. Physiol. 2020, 82, 485–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wesseling-Perry, K.; Juppner, H. The osteocyte in CKD: New concepts regarding the role of FGF23 in mineral metabolism and systemic complications. Bone 2013, 54, 222–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, C.; David, V. Inflammation regulates fibroblast growth factor 23 production. Curr. Opin. Nephrol. Hypertens. 2016, 25, 325–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erben, R.G. Pleiotropic Actions of FGF23. Toxicol. Pathol. 2017, 45, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Mullender, M.G.; van der Meer, D.D.; Huiskes, R.; Lips, P. Osteocyte density changes in aging and osteoporosis. Bone 1996, 18, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Bach-Gansmo, F.L.; Bruel, A.; Jensen, M.V.; Ebbesen, E.N.; Birkedal, H.; Thomsen, J.S. Osteocyte lacunar properties and cortical microstructure in human iliac crest as a function of age and sex. Bone 2016, 91, 11–19. [Google Scholar]
- Zimmerman, S.M.; Dimori, M.; Heard-Lipsmeyer, M.E.; Morello, R. The Osteocyte Transcriptome is Extensively Dysregulated in Mouse Models of Osteogenesis Imperfecta. JBMR Plus 2019, 3, e10171. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, F.S.; Sillence, D.O. Osteogenesis imperfecta: Clinical diagnosis, nomenclature and severity assessment. Am. J. Med. Genet. A 2014, 164A, 1470–1481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marini, J.C.; Forlino, A.; Bachinger, H.P.; Bishop, N.J.; Byers, P.H.; Paepe, A.; Fassier, F.; Fratzl-Zelman, N.; Kozloff, K.M.; Krakow, D.; et al. Osteogenesis imperfecta. Nat. Rev. Dis. Prim. 2017, 3, 17052. [Google Scholar] [CrossRef] [PubMed]
- Forlino, A.; Cabral, W.A.; Barnes, A.M.; Marini, J.C. New perspectives on osteogenesis imperfecta. Nat. Rev. Endocrinol. 2011, 7, 540–557. [Google Scholar] [CrossRef] [Green Version]
- Tauer, J.T.; Robinson, M.E.; Rauch, F. Osteogenesis Imperfecta: New Perspectives from Clinical and Translational Research. JBMR Plus 2019, 3, e10174. [Google Scholar] [CrossRef] [Green Version]
- Rauch, F.; Glorieux, F.H. Osteogenesis imperfecta. Lancet 2004, 363, 1377–1385. [Google Scholar] [CrossRef]
- Fratzl-Zelman, N.; Misof, B.M.; Roschger, P.; Klaushofer, K. Classification of osteogenesis imperfecta. Wiener Medizinische Wochenschrift 2015, 165, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Pathak, J.L.; Bravenboer, N.; Klein-Nulend, J. The Osteocyte as the New Discovery of Therapeutic Options in Rare Bone Diseases. Front. Endocrinol. 2020, 11, 405. [Google Scholar] [CrossRef]
- Jones, S.J.; Glorieux, F.H.; Travers, R.; Boyde, A. The microscopic structure of bone in normal children and patients with osteogenesis imperfecta: A survey using backscattered electron imaging. Calcif. Tissue Int. 1999, 64, 8–17. [Google Scholar] [CrossRef]
- Rauch, F.; Travers, R.; Parfitt, A.M.; Glorieux, F.H. Static and dynamic bone histomorphometry in children with osteogenesis imperfecta. Bone 2000, 26, 581–589. [Google Scholar] [CrossRef]
- Imbert, L.; Auregan, J.C.; Pernelle, K.; Hoc, T. Mechanical and mineral properties of osteogenesis imperfecta human bones at the tissue level. Bone 2014, 65, 18–24. [Google Scholar] [CrossRef]
- Albert, C.; Jameson, J.; Smith, P.; Harris, G. Reduced diaphyseal strength associated with high intracortical vascular porosity within long bones of children with osteogenesis imperfecta. Bone 2014, 66, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imbert, L.; Auregan, J.C.; Pernelle, K.; Hoc, T. Microstructure and compressive mechanical properties of cortical bone in children with osteogenesis imperfecta treated with bisphosphonates compared with healthy children. J. Mech. Behav. Biomed. Mater. 2015, 46, 261–270. [Google Scholar] [CrossRef]
- Zysset, P.K.; Guo, X.E.; Hoffler, C.E.; Moore, K.E.; Goldstein, S.A. Elastic modulus and hardness of cortical and trabecular bone lamellae measured by nanoindentation in the human femur. J. Biomech. 1999, 32, 1005–1012. [Google Scholar] [CrossRef]
- Van Tol, A.F.; Schemenz, V.; Wagermaier, W.; Roschger, A.; Razi, H.; Vitienes, I.; Fratzl, P.; Willie, B.M.; Weinkamer, R. The mechanoresponse of bone is closely related to the osteocyte lacunocanalicular network architecture. Proc. Natl. Acad. Sci. USA 2020, 117, 32251–32259. [Google Scholar] [CrossRef]
- Carriero, A.; Doube, M.; Vogt, M.; Busse, B.; Zustin, J.; Levchuk, A.; Schneider, P.; Muller, R.; Shefelbine, S.J. Altered lacunar and vascular porosity in osteogenesis imperfecta mouse bone as revealed by synchrotron tomography contributes to bone fragility. Bone 2014, 61, 116–124. [Google Scholar] [CrossRef]
- Jin, J.; Bakker, A.D.; Wu, G.; Klein-Nulend, J.; Jaspers, R.T. Physicochemical Niche Conditions and Mechanosensing by Osteocytes and Myocytes. Curr. Osteoporos. Rep. 2019, 17, 235–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedarko, N.S.; Robey, P.G.; Vetter, U.K. Extracellular matrix stoichiometry in osteoblasts from patients with osteogenesis imperfecta. J. Bone Miner. Res. 1995, 10, 1122–1129. [Google Scholar] [CrossRef]
- Paschalis, E.P.; Gamsjaeger, S.; Fratzl-Zelman, N.; Roschger, P.; Masic, A.; Brozek, W.; Hassler, N.; Glorieux, F.H.; Rauch, F.; Klaushofer, K.; et al. Evidence for a Role for Nanoporosity and Pyridinoline Content in Human Mild Osteogenesis Imperfecta. J. Bone Miner. Res. 2016, 31, 1050–1059. [Google Scholar] [CrossRef]
- El-Gazzar, A.; Hogler, W. Mechanisms of Bone Fragility: From Osteogenesis Imperfecta to Secondary Osteoporosis. Int. J. Mol. Sci. 2021, 22, 625. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, S.M.; Heard-Lipsmeyer, M.E.; Dimori, M.; Thostenson, J.D.; Mannen, E.M.; O’Brien, C.A.; Morello, R. Loss of RANKL in osteocytes dramatically increases cancellous bone mass in the osteogenesis imperfecta mouse (oim). Bone Rep. 2018, 9, 61–73. [Google Scholar] [CrossRef]
- Uveges, T.E.; Collin-Osdoby, P.; Cabral, W.A.; Ledgard, F.; Goldberg, L.; Bergwitz, C.; Forlino, A.; Osdoby, P.; Gronowicz, G.A.; Marini, J.C. Cellular mechanism of decreased bone in Brtl mouse model of OI: Imbalance of decreased osteoblast function and increased osteoclasts and their precursors. J. Bone Miner. Res. 2008, 23, 1983–1994. [Google Scholar] [CrossRef]
- Brunetti, G.; Papadia, F.; Tummolo, A.; Fischetto, R.; Nicastro, F.; Piacente, L.; Ventura, A.; Mori, G.; Oranger, A.; Gigante, I.; et al. Impaired bone remodeling in children with osteogenesis imperfecta treated and untreated with bisphosphonates: The role of DKK1, RANKL, and TNF-alpha. Osteoporos. Int. 2016, 27, 2355–2365. [Google Scholar] [CrossRef]
- Palomo, T.; Glorieux, F.H.; Rauch, F. Circulating sclerostin in children and young adults with heritable bone disorders. J. Clin. Endocrinol. Metab. 2014, 99, E920–E925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocijan, R.; Muschitz, C.; Fahrleitner-Pammer, A.; Amrein, K.; Pietschmann, P.; Haschka, J.; Dinu, S.; Kapiotis, S.; Resch, H. Serum sclerostin levels are decreased in adult patients with different types of osteogenesis imperfecta. J. Clin. Endocrinol. Metab. 2014, 99, E311–E319. [Google Scholar] [CrossRef]
- Nicol, L.; Wang, Y.; Smith, R.; Sloan, J.; Nagamani, S.C.; Shapiro, J.; Lee, B.; Orwoll, E. Serum Sclerostin Levels in Adults with Osteogenesis Imperfecta: Comparison with Normal Individuals and Response to Teriparatide Therapy. J. Bone Miner. Res. 2018, 33, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Sinder, B.P.; Eddy, M.M.; Ominsky, M.S.; Caird, M.S.; Marini, J.C.; Kozloff, K.M. Sclerostin antibody improves skeletal parameters in a Brtl/+ mouse model of osteogenesis imperfecta. J. Bone Miner. Res. 2013, 28, 73–80. [Google Scholar] [CrossRef]
- Cardinal, M.; Dessain, A.; Roels, T.; Lafont, S.; Ominsky, M.S.; Devogelaer, J.P.; Chappard, D.; Mabilleau, G.; Ammann, P.; Nyssen-Behets, C.; et al. Sclerostin-Antibody Treatment Decreases Fracture Rates in Axial Skeleton and Improves the Skeletal Phenotype in Growing oim/oim Mice. Calcif. Tissue Int. 2020, 106, 494–508. [Google Scholar] [CrossRef] [PubMed]
- Glorieux, F.H.; Devogelaer, J.P.; Durigova, M.; Goemaere, S.; Hemsley, S.; Jakob, F.; Junker, U.; Ruckle, J.; Seefried, L.; Winkle, P.J. BPS804 Anti-Sclerostin Antibody in Adults with Moderate Osteogenesis Imperfecta: Results of a Randomized Phase 2a Trial. J. Bone Miner. Res. 2017, 32, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- Bateman, J.F.; Boot-Handford, R.P.; Lamande, S.R. Genetic diseases of connective tissues: Cellular and extracellular effects of ECM mutations. Nat. Rev. Genet. 2009, 10, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Lisse, T.S.; Thiele, F.; Fuchs, H.; Hans, W.; Przemeck, G.K.; Abe, K.; Rathkolb, B.; Quintanilla-Martinez, L.; Hoelzlwimmer, G.; Helfrich, M.; et al. ER stress-mediated apoptosis in a new mouse model of osteogenesis imperfecta. PLoS Genet. 2008, 4, e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besio, R.; Iula, G.; Garibaldi, N.; Cipolla, L.; Sabbioneda, S.; Biggiogera, M.; Marini, J.C.; Rossi, A.; Forlino, A. 4-PBA ameliorates cellular homeostasis in fibroblasts from osteogenesis imperfecta patients by enhancing autophagy and stimulating protein secretion. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864 Pt A, 1642–1652. [Google Scholar] [CrossRef]
- Bateman, J.F.; Sampurno, L.; Maurizi, A.; Lamande, S.R.; Sims, N.A.; Cheng, T.L.; Schindeler, A.; Little, D.G. Effect of rapamycin on bone mass and strength in the alpha2(I)-G610C mouse model of osteogenesis imperfecta. J. Cell. Mol. Med. 2019, 23, 1735–1745. [Google Scholar] [CrossRef]
- Glorieux, F.H.; Travers, R.; Taylor, A.; Bowen, J.R.; Rauch, F.; Norman, M.; Parfitt, A.M. Normative data for iliac bone histomorphometry in growing children. Bone 2000, 26, 103–109. [Google Scholar] [CrossRef]
- Roschger, P.; Fratzl-Zelman, N.; Misof, B.M.; Glorieux, F.H.; Klaushofer, K.; Rauch, F. Evidence that abnormal high bone mineralization in growing children with osteogenesis imperfecta is not associated with specific collagen mutations. Calcif. Tissue Int. 2008, 82, 263–270. [Google Scholar] [CrossRef]
- Fratzl-Zelman, N.; Roschger, P.; Misof, B.M.; Pfeffer, S.; Glorieux, F.H.; Klaushofer, K.; Rauch, F. Normative data on mineralization density distribution in iliac bone biopsies of children, adolescents and young adults. Bone 2009, 44, 1043–1048. [Google Scholar] [CrossRef]
- Glorieux, F.H.; Rauch, F.; Plotkin, H.; Ward, L.; Travers, R.; Roughley, P.; Lalic, L.; Glorieux, D.F.; Fassier, F.; Bishop, N.J. Type V osteogenesis imperfecta: A new form of brittle bone disease. J. Bone Miner. Res. 2000, 15, 1650–1658. [Google Scholar] [CrossRef] [PubMed]
- Semler, O.; Garbes, L.; Keupp, K.; Swan, D.; Zimmermann, K.; Becker, J.; Iden, S.; Wirth, B.; Eysel, P.; Koerber, F.; et al. A mutation in the 5′-UTR of IFITM5 creates an in-frame start codon and causes autosomal-dominant osteogenesis imperfecta type V with hyperplastic callus. Am. J. Hum. Genet. 2012, 91, 349–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, T.J.; Lee, K.E.; Lee, S.K.; Song, S.J.; Kim, K.J.; Jeon, D.; Lee, G.; Kim, H.N.; Lee, H.R.; Eom, H.H.; et al. A single recurrent mutation in the 5’-UTR of IFITM5 causes osteogenesis imperfecta type V. Am. J. Hum. Genet. 2012, 91, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Blouin, S.; Fratzl-Zelman, N.; Glorieux, F.H.; Roschger, P.; Klaushofer, K.; Marini, J.C.; Rauch, F. Hypermineralization and High Osteocyte Lacunar Density in Osteogenesis Imperfecta Type V Bone Indicate Exuberant Primary Bone Formation. J. Bone Miner. Res. 2017, 32, 1884–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, M.A.; Blouin, S.; Misof, B.M.; Fratzl-Zelman, N.; Roschger, P.; Berzlanovich, A.; Gruber, G.M.; Brugger, P.C.; Zwerina, J.; Fratzl, P. Quantitative Backscattered Electron Imaging of Bone Using a Thermionic or a Field Emission Electron Source. Calcif. Tissue Int. 2021, 1–13. [Google Scholar] [CrossRef]
- Fratzl-Zelman, N.; Schmidt, I.; Roschger, P.; Glorieux, F.H.; Klaushofer, K.; Fratzl, P.; Rauch, F.; Wagermaier, W. Mineral particle size in children with osteogenesis imperfecta type I is not increased independently of specific collagen mutations. Bone 2014, 60, 122–128. [Google Scholar] [CrossRef]
- Nijhuis, W.H.; Eastwood, D.M.; Allgrove, J.; Hvid, I.; Weinans, H.H.; Bank, R.A.; Sakkers, R.J. Current concepts in osteogenesis imperfecta: Bone structure, biomechanics and medical management. J. Child. Orthop. 2019, 13, 1–11. [Google Scholar] [CrossRef]
- Grabner, B.; Landis, W.J.; Roschger, P.; Rinnerthaler, S.; Peterlik, H.; Klaushofer, K.; Fratzl, P. Age- and genotype-dependence of bone material properties in the osteogenesis imperfecta murine model (oim). Bone 2001, 29, 453–457. [Google Scholar] [CrossRef]
- Zimmermann, E.A.; Riedel, C.; Schmidt, F.N.; Stockhausen, K.E.; Chushkin, Y.; Schaible, E.; Gludovatz, B.; Vettorazzi, E.; Zontone, F.; Puschel, K.; et al. Mechanical Competence and Bone Quality Develop During Skeletal Growth. J. Bone Miner. Res. 2019, 34, 1461–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarathchandra, P.; Pope, F.M.; Kayser, M.V.; Ali, S.Y. A light and electron microscopic study of osteogenesis imperfecta bone samples, with reference to collagen chemistry and clinical phenotype. J. Pathol. 2000, 192, 385–395. [Google Scholar] [CrossRef]
- Grafe, I.; Yang, T.; Alexander, S.; Homan, E.P.; Lietman, C.; Jiang, M.M.; Bertin, T.; Munivez, E.; Chen, Y.; Dawson, B.; et al. Excessive transforming growth factor-beta signaling is a common mechanism in osteogenesis imperfecta. Nat. Med. 2014, 20, 670–675. [Google Scholar] [CrossRef] [Green Version]
- Jandl, N.M.; von Kroge, S.; Sturznickel, J.; Baranowsky, A.; Stockhausen, K.E.; Mushumba, H.; Beil, F.T.; Puschel, K.; Amling, M.; Rolvien, T. Large osteocyte lacunae in iliac crest infantile bone are not associated with impaired mineral distribution or signs of osteocytic osteolysis. Bone 2020, 135, 115324. [Google Scholar] [CrossRef]
- Lai, X.; Price, C.; Modla, S.; Thompson, W.R.; Caplan, J.; Kirn-Safran, C.B.; Wang, L. The dependences of osteocyte network on bone compartment, age, and disease. Bone Res. 2015, 3, 15009. [Google Scholar] [CrossRef] [Green Version]
- Rauch, F.; Travers, R.; Glorieux, F.H. Intracortical remodeling during human bone development—A histomorphometric study. Bone 2006, 40, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Parfitt, A.M.; Travers, R.; Rauch, F.; Glorieux, F.H. Structural and cellular changes during bone growth in healthy children. Bone 2000, 27, 487–494. [Google Scholar] [CrossRef]
- Rauch, F.; Travers, R.; Glorieux, F.H. Cellular activity on the seven surfaces of iliac bone: A histomorphometric study in children and adolescents. J. Bone Miner. Res. 2006, 21, 513–519. [Google Scholar] [CrossRef]
- Bisseret, D.; Kaci, R.; Lafage-Proust, M.H.; Alison, M.; Parlier-Cuau, C.; Laredo, J.D.; Bousson, V. Periosteum: Characteristic imaging findings with emphasis on radiologic-pathologic comparisons. Skelet. Radiol. 2015, 44, 321–338. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, C.J.; Majeska, R.J.; Schaffler, M.B. Osteocyte density in woven bone. Bone 2004, 35, 1095–1099. [Google Scholar] [CrossRef]
- Repp, F.; Kollmannsberger, P.; Roschger, A.; Berzlanovich, A.; Gruber, G.M.; Roschger, P.; Wagermaier, W.; Weinkamer, R. Coalignment of osteocyte canaliculi and collagen fibers in human osteonal bone. J. Struct. Biol. 2017, 199, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Currey, J.D. The many adaptations of bone. J. Biomech. 2003, 36, 1487–1495. [Google Scholar] [CrossRef]
- Chen, X.; Wang, L.; Zhao, K.; Wang, H. Osteocytogenesis: Roles of Physicochemical Factors, Collagen Cleavage, and Exogenous Molecules. Tissue Eng. Part B Rev. 2018, 24, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, L.I.; Bellido, T. Osteocytic signalling pathways as therapeutic targets for bone fragility. Nat. Rev. Endocrinol. 2016, 12, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Schnitzler, C.M.; Mesquita, J.M.; Pettifor, J.M. Cortical bone development in black and white South African children: Iliac crest histomorphometry. Bone 2009, 44, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Schnitzler, C.M.; Pettifor, J.M. Calcium Deficiency Rickets in African Adolescents: Cortical Bone Histomorphometry. JBMR Plus 2019, 3, e10169. [Google Scholar] [CrossRef]
- Vashishth, D.; Verborgt, O.; Divine, G.; Schaffler, M.B.; Fyhrie, D.P. Decline in osteocyte lacunar density in human cortical bone is associated with accumulation of microcracks with age. Bone 2000, 26, 375–380. [Google Scholar] [CrossRef]
- Milovanovic, P.; Zimmermann, E.A.; Riedel, C.; Scheidt, A.V.; Herzog, L.; Krause, M.; Djonic, D.; Djuric, M.; Puschel, K.; Amling, M.; et al. Multi-level characterization of human femoral cortices and their underlying osteocyte network reveal trends in quality of young, aged, osteoporotic and antiresorptive-treated bone. Biomaterials 2015, 45, 46–55. [Google Scholar] [CrossRef]
- Hemmatian, H.; Bakker, A.D.; Klein-Nulend, J.; van Lenthe, G.H. Aging, Osteocytes, and Mechanotransduction. Curr. Osteoporos. Rep. 2017, 15, 401–411. [Google Scholar] [CrossRef] [Green Version]
- Busse, B.; Djonic, D.; Milovanovic, P.; Hahn, M.; Puschel, K.; Ritchie, R.O.; Djuric, M.; Amling, M. Decrease in the osteocyte lacunar density accompanied by hypermineralized lacunar occlusion reveals failure and delay of remodeling in aged human bone. Aging Cell 2010, 9, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Roschger, P.; Misof, B.; Paschalis, E.; Fratzl, P.; Klaushofer, K. Changes in the degree of mineralization with osteoporosis and its treatment. Curr. Osteoporos. Rep. 2014, 12, 338–350. [Google Scholar] [CrossRef]
- Boyde, A.; Travers, R.; Glorieux, F.H.; Jones, S.J. The mineralization density of iliac crest bone from children with osteogenesis imperfecta. Calcif. Tissue Int. 1999, 64, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Roschger, P.; Fratzl, P.; Eschberger, J.; Klaushofer, K. Validation of quantitative backscattered electron imaging for the measurement of mineral density distribution in human bone biopsies. Bone 1998, 23, 319–326. [Google Scholar] [CrossRef]
- Roschger, P.; Paschalis, E.P.; Fratzl, P.; Klaushofer, K. Bone mineralization density distribution in health and disease. Bone 2008, 42, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Misof, B.M.; Blouin, S.; Hofstaetter, J.G.; Roschger, P.; Zwerina, J.; Erben, R.G. No Role of Osteocytic Osteolysis in the Development and Recovery of the Bone Phenotype Induced by Severe Secondary Hyperparathyroidism in Vitamin D Receptor Deficient Mice. Int. J. Mol. Sci. 2020, 21, 7989. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trabecular Bone | Cortical Bone | Trabecular Versus Cortical Bone | ||||||
|---|---|---|---|---|---|---|---|---|
| OLS parameters: | OI type I (n = 19) * | Controls (n = 24) | p-value | OI type I (n = 19) * | Controls (n = 24) | p-value | OI type I (p-value) | Controls (p-value) |
| Density (number/mm2) | 365.60 (67.73) | 226.00 (26.75) | <0.0001 | 413.80 (86.25) | 274.65 (60.14) | <0.0001 | 0.0334 | <0.0001 |
| porosity (%) | 0.76 [0.64; 0.81] | 0.54 [0.48; 0.60] | <0.0001 | 0.88 [0.72; 1.06] | 0.60 [0.53; 0.69] | <0.0001 | 0.0268 | 0.0014 |
| area (µm2) | 21.20 (3.16) | 23.63 (3.01) | 0.0150 | 21.53 (3.66) | 23.19 (3.34) | 0.1278 | 0.5944 | 0.5211 |
| perimeter (µm) | 19.65 (1.51) | 20.42 (1.50) | 0.1070 | 20.61 (2.21) | 20.30 (2.09) | 0.6368 | 0.1110 | 0.7512 |
| aspect-ratio | 2.63 (0.18) | 2.55 (0.22) | 0.2185 | 2.92 [2.63; 3.27] | 2.50 [2.27; 2.72] | 0.0003 | 0.0020 | 0.2522 |
| Trabecular Bone | ||||
|---|---|---|---|---|
| BMDD parameters | Controls (n = 24) | Controls (n = 50) | OI type I (n = 19) * | Diff. OI type I versus controls (n = 50) |
| CaMean (weight % calcium) | 22.45 (0.73) 22.45 [22.25; 22.80] | 22.48 (0.73) 22.60 [22.12; 22.96] | 23.30 (0.59) 23.28 [22.92; 23.64] | p < 0.0001 |
| CaPeak (weight % calcium) | 23.40 (0.75) 23.40 [23.09; 23.92] | 23.39 (0.70) 23.57 [23.01; 23.79] | 24.45 (0.61) 24.44 [24.05; 24.83] | p < 0.0001 |
| CaWidth (Δ weight % calcium) | 3.91 (0.47) 3.81 [3.64; 3.99] | 3.76 (0.50) 3.64 [3.47; 3.99] | 3.39 (0.20) 3.29 [3.29; 3.51] | p = 0.003 |
| CaLow (% bone area) | 6.46 (1.90) 5.95 [5.39; 6.95] | 6.14 (2.21) 5.57 [4.78; 6.80] | 6. 55 (1.65) 6.19 [5.31; 7.41] | p = 0.1475 |
| CaHigh (% bone area) | 2.04 (1.93) 1.53 [0.69; 2.48] | 1.82 (1.64) 1.52 [0.62; 2.22] | 5.77 (4.79) 4.29 [2.00; 8.48] | p < 0.0001 |
| Cortical Bone (calculated as arithmetic mean of both cortical plates) | ||||
| BMDD parameters | Controls (n = 24) | Controls (n = 50) ** | OI type I (n = 19) * | Diff. OI type I versus controls (n = 50) |
| CaMean (weight % calcium) | 21.84 (1.18) 22.02 [21.41; 22.84] | 21.86 (1.15) 22.17 [21.05; 22.76] | 23.42 (0.71) 23.50 [22.97; 24.07] | p < 0.0001 |
| CaPeak (weight % calcium) | 22.66 (1.29) 22.88 [22.38; 23.57] | 22.67 (1.21) 22.96 [22.10; 23.48] | 24.33 (0.70) 24.52 [23.92; 24.87] | p < 0.0001 |
| CaWidth (Δ weight % calcium) | 4.32 (0.76) 4.16 [3.75; 4.98] | 4.23 (0.67) 4.07 [3.73; 4.68] | 3.36 (0.29) 3.38 [3.21; 3.64] | p < 0.0001 |
| CaLow (% bone area) | 9.63 (6.76) 7.28 [5.07; 10.78] | 9.19 (6.18) 6.86 [5.06; 11.48] | 4.98 (0.87) 5.02 [4.38; 5.68] | p < 0.0007 |
| CaHigh (% bone area) | 1.47 (1.15) 1.01 [0.66; 2.26] | 1.37 (1.21) 1.01 [0.44; 1.89] | 4.75 (3.61) 3.52 [2.05; 6.49] | p < 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mähr, M.; Blouin, S.; Behanova, M.; Misof, B.M.; Glorieux, F.H.; Zwerina, J.; Rauch, F.; Hartmann, M.A.; Fratzl-Zelman, N. Increased Osteocyte Lacunae Density in the Hypermineralized Bone Matrix of Children with Osteogenesis Imperfecta Type I. Int. J. Mol. Sci. 2021, 22, 4508. https://doi.org/10.3390/ijms22094508
Mähr M, Blouin S, Behanova M, Misof BM, Glorieux FH, Zwerina J, Rauch F, Hartmann MA, Fratzl-Zelman N. Increased Osteocyte Lacunae Density in the Hypermineralized Bone Matrix of Children with Osteogenesis Imperfecta Type I. International Journal of Molecular Sciences. 2021; 22(9):4508. https://doi.org/10.3390/ijms22094508
Chicago/Turabian StyleMähr, Matthias, Stéphane Blouin, Martina Behanova, Barbara M. Misof, Francis H. Glorieux, Jochen Zwerina, Frank Rauch, Markus A. Hartmann, and Nadja Fratzl-Zelman. 2021. "Increased Osteocyte Lacunae Density in the Hypermineralized Bone Matrix of Children with Osteogenesis Imperfecta Type I" International Journal of Molecular Sciences 22, no. 9: 4508. https://doi.org/10.3390/ijms22094508
APA StyleMähr, M., Blouin, S., Behanova, M., Misof, B. M., Glorieux, F. H., Zwerina, J., Rauch, F., Hartmann, M. A., & Fratzl-Zelman, N. (2021). Increased Osteocyte Lacunae Density in the Hypermineralized Bone Matrix of Children with Osteogenesis Imperfecta Type I. International Journal of Molecular Sciences, 22(9), 4508. https://doi.org/10.3390/ijms22094508