
Expression Analysis of Zinc Transporters in Nervous Tissue Cells Reveals Neuronal and Synaptic Localization of ZIP4



Abstract
:1. Introduction
2. Results
2.1. Expression of Zinc Transporters in Adult Rat Brain and Rat Neurons and Astrocytes on mRNA Level
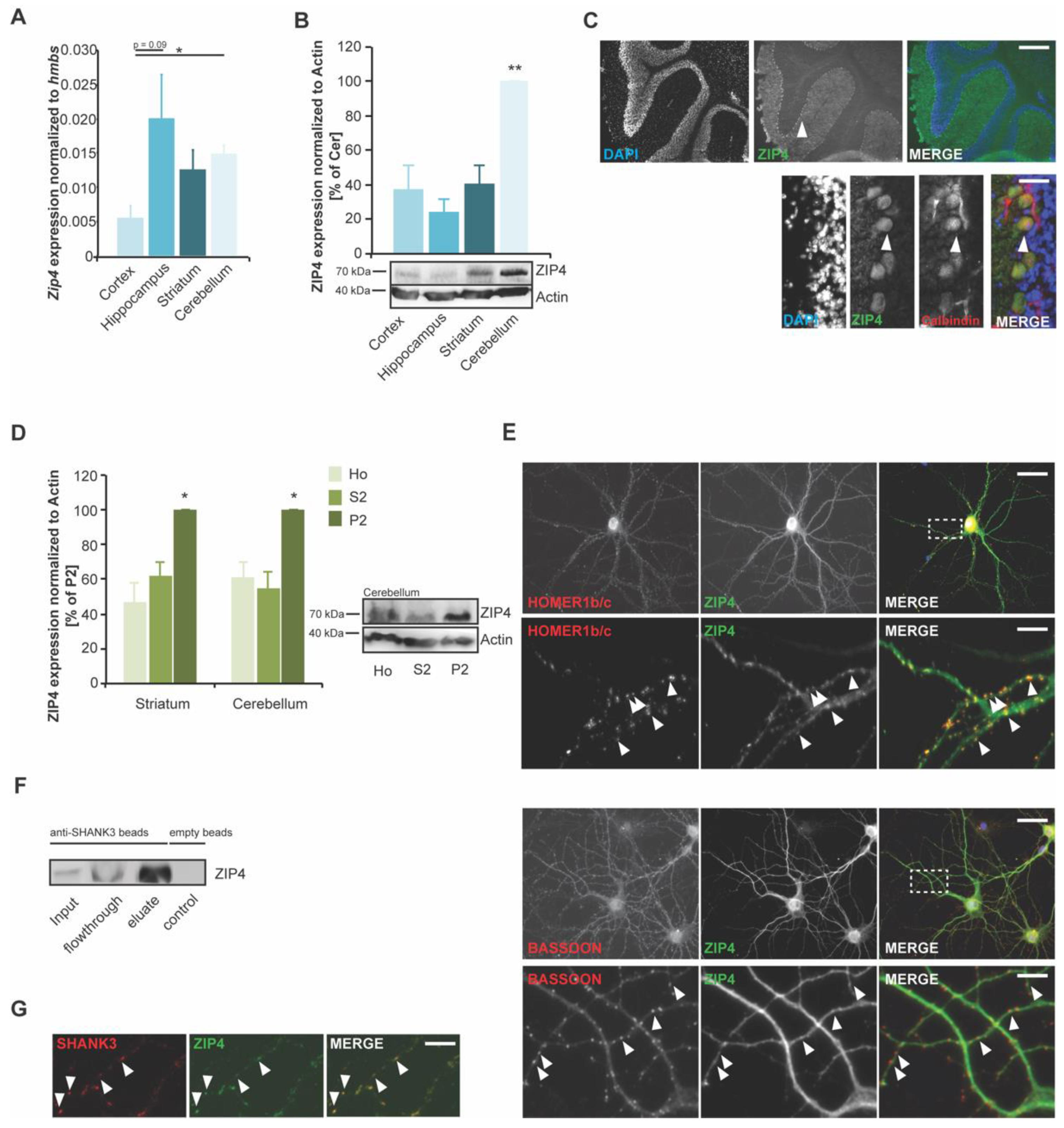
2.2. Zip4 Is Expressed and Localized at Glutamatergic Synapses in the Brain
2.3. ZIP4 Expression Is Sensitive to Local Zinc Levels
2.4. Synaptic Expression of ZIP4 Shows Dynamic Responses to Transient Local Changes in Zinc Concentrations
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Preparation and Cultivation of Primary Hippocampal Neurons
4.4. Cultivation of Cell Lines
4.5. Immunocytochemistry (ICC)
4.6. Immunohistochemistry (IHC)
4.7. Cell Lysis and Total RNA Preparation
4.8. qRT–PCR
4.9. Protein Biochemistry
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| cer | Cerebellum |
| ctx | Cortex |
| DIV | Days in vitro |
| hip | Hippocampus |
| HRP | Horseradish peroxidase |
| RT | Room temperature |
| str | Striatum |
| TPEN | N,N,N′,N′-tetrakis(2-pyridinylmethyl)-1,2-ethanediamine |
References
- Sauer, A.K.; Hagmeyer, S.; Grabrucker, A.M. Zinc Deficiency. In Nutritional Deficiency; Erkekoglu, P., Kocer-Gumusel, B., Eds.; InTech Open Science: Rijeka, Croatia, 2016; pp. 23–46. [Google Scholar] [CrossRef]
- McAllister, B.B.; Dyck, R.H. Zinc transporter 3 (ZnT3) and vesicular zinc in central nervous system function. Neurosci. Biobehav. 2017, 80, 329–350. [Google Scholar] [CrossRef]
- Mellone, M.; Pelucchi, S.; Alberti, L.; Genazzani, A.A.; Di Luca, M.; Gardoni, F. Zinc transporter-1: A novel NMDA receptor-binding protein at the postsynaptic density. J. Neurochem. 2015, 132, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowanadisai, W.; Graham, D.M.; Keen, C.L.; Rucker, R.B.; Messerli, M.A. Neurulation and neurite extension require the zinc transporter ZIP12 (slc39a12). Proc. Natl. Acad. Sci. USA 2013, 110, 9903–9908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belloni-Olivi, L.; Marshall, C.; Laal, B.; Andrews, G.K.; Bressler, J. Localization of zip1 and zip4 mRNA in the adult rat brain. J. Neurosci. Res. 2009, 87, 3221–3230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Chen, Y.; Wang, Y.; Yang, J.; Zhu, V.F.; Liu, Y.; Cui, X.; Chen, L.; Yan, W.; Jiang, T.; et al. ZIP4 is a novel molecular marker for glioma. Neuro. Oncol. 2013, 15, 1008–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, X.; Chen, R.; Zhang, J.; Li, G.; Dai, P.G.; Chen, C.; Wang, H.J. Expression Profile Analysis of Zinc Transporters (ZIP4, ZIP9, ZIP11, ZnT9) in Gliomas and their Correlation with IDH1 Mutation Status. Asian Pac. J. Cancer Prev. 2015, 16, 3355–3360. [Google Scholar] [CrossRef]
- Cousins, R.J. Gastrointestinal factors influencing zinc absorption and homeostasis. Int. J. Vitam. Nutr. Res. 2010, 80, 243–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kambe, T.; Andrews, G.K. Novel proteolytic processing of the ectodomain of the zinc transporter ZIP4 (SLC39A4) during zinc deficiency is inhibited by acrodermatitis enteropathica mutations. Mol. Cell Biol. 2009, 29, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Kim, B.E.; Wang, F.; Eide, D.J.; Petris, M.J. A histidine-rich cluster mediates the ubiquitination and degradation of the human zinc transporter, hZIP4, and protects against zinc cytotoxicity. J. Biol. Chem. 2007, 282, 6992–7000. [Google Scholar] [CrossRef] [Green Version]
- Dufner-Beattie, J.; Langmade, S.J.; Wang, F.; Eide, D.; Andrews, G.K. Structure, function, and regulation of a subfamily of mouse zinc transporter genes. J. Biol. Chem. 2003, 278, 50142–50150. [Google Scholar] [CrossRef] [Green Version]
- Weaver, B.P.; Dufner-Beattie, J.; Kambe, T.; Andrews, G.K. Novel zinc-responsive post-transcriptional mechanisms reciprocally regulate expression of the mouse Slc39a4 and Slc39a5 zinc transporters (Zip4 and Zip5). Biol. Chem. 2007, 388, 1301–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liuzzi, J.P.; Guo, L.; Chang, S.M.; Cousins, R.J. Krüppel-like factor 4 regulates adaptive expression of the zinc transporter Zip4 in mouse small intestine. Am. J. Physiol. Gastrointest Liver Physiol. 2009, 296, G517–G523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Küry, S.; Dréno, B.; Bézieau, S.; Giraudet, S.; Kharfi, M.; Kamoun, R.; Moisan, J.P. Identification of SLC39A4, a gene involved in acrodermatitis enteropathica. Nat. Genet. 2002, 31, 239–240. [Google Scholar] [CrossRef] [PubMed]
- Braun, O.H.; Heilmann, K.; Pauli, W.; Rossner, J.A.; Bergmann, K.E. Acrodermatitis enteropathica: Recent findings concerning clinical features, pathogenesis, diagnosis and therapy. Eur. J. Pediatr. 1976, 121, 247–261. [Google Scholar] [CrossRef]
- Dillaha, C.J.; Lorincz, A.L.; Aavik, O.R. Acrodermatitis enteropathica; review of the literature and report of a case successfully treated with diodoquin. J. Am. Med. Assoc. 1953, 152, 509–512. [Google Scholar] [CrossRef] [Green Version]
- Gözdasoğlu, S.; Taçyıldız, N.; Günlemez, A.; Bayhan, H.; Sencer, H.; Ünal, E.; İkincioğulları, A.; İnceoğlu, H.; Aksoylar, S.; Yavuz, G.; et al. Acrodermatitis enteropathica: Case report analyses of zinc metabolism electron microscopic examination and immune function. J. Trace Elem. Exp. Med. 2000, 13, 317–325. [Google Scholar] [CrossRef]
- Idriss, Z.H.; Der Kaloustian, V.M. Acrodermatitis enteropathica: Report of three new cases. Clin. Pediatr. Phila. 1973, 12, 393–395. [Google Scholar] [CrossRef] [Green Version]
- Julius, R.; Schulkind, M.; Sprinkle, T.; Rennert, O. Acrodermatitis enteropathica with immune deficiency. J. Pediatr. 1973, 83, 1007–1011. [Google Scholar] [CrossRef]
- Maverakis, E.; Fung, M.A.; Lynch, P.J.; Draznin, M.; Michael, D.J.; Ruben, B.; Fazel, N. Acrodermatitis enteropathica and an overview of zinc metabolism. J. Am. Acad. Dermatol. 2007, 56, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Michaëlsson, G. Zinc therapy in acrodermatitis enteropathica. Acta Derm. Venereol. 1974, 54, 377–381. [Google Scholar]
- Perafán-Riveros, C.; França, L.F.S.; Alves, A.C.F.; Sanches, J.A. Acrodermatitis enteropathica: Case report and review of the literature. Pediatr. Dermatol. 2002, 19, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Van Wouwe, J.P. Clinical and laboratory diagnosis of acrodermatitis enteropathica. Eur. J. Pediatr. 1989, 149, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Wirsching, L. Eye symptoms in Acromdermatitis enteropathica. A description of a brother and sister, with corneal changes. Acta Ophthalmol. 1962, 40, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Beyer, N.; Coulson, D.T.; Heggarty, S.; Ravid, R.; Hellemans, J.; Irvine, G.B.; Johnston, J.A. Zinc transporter mRNA levels in Alzheimer’s disease postmortem brain. J. Alzheimers Dis. 2012, 29, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Bosomworth, H.J.; Adlard, P.A.; Ford, D.; Valentine, R.A. Altered expression of ZnT10 in Alzheimer’s disease brain. PLoS ONE 2013, 8, e65475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovell, M.A.; Smith, J.L.; Markesbery, W.R. Elevated zinc transporter-6 in mild cognitive impairment, Alzheimer disease, and pick disease. J. Neuropathol. Exp. Neurol. 2006, 65, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarr, E.; Udawela, M.; Greenough, M.A.; Neo, J.; Seo, M.S.; Money, T.T.; Upadhyay, A.; Bush, A.I.; Everall, I.P.; Thomas, E.A.; et al. Increased cortical expression of the zinc transporter SLC39A12 suggests a breakdown in zinc cellular homeostasis as part of the pathophysiology of schizophrenia. NPJ Schizophr. 2016, 2, 16002. [Google Scholar] [CrossRef] [Green Version]
- Ajjimaporn, A.; Botsford, T.; Garrett, S.H.; Sens, M.A.; Zhou, X.D.; Dunlevy, J.R.; Sens, D.A.; Somji, S. ZIP8 expression in human proximal tubule cells, human urothelial cells transformed by Cd+2 and As+3 and in specimens of normal human urothelium and urothelial cancer. Cancer Cell Int. 2012, 12, 16. [Google Scholar] [CrossRef] [Green Version]
- Mei, Z.; Yan, P.; Wang, Y.; Liu, S.; He, F. Knockdown of zinc transporter ZIP8 expression inhibits neuroblastoma progression and metastasis in vitro. Mol. Med. Rep. 2018, 18, 477–485. [Google Scholar] [CrossRef]
- Grabrucker, S.; Boeckers, T.M.; Grabrucker, A.M. Gender Dependent Evaluation of Autism like Behavior in Mice Exposed to Prenatal Zinc Deficiency. Front. Behav. Neurosci. 2016, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Grabrucker, S.; Jannetti, L.; Eckert, M.; Gaub, S.; Chhabra, R.; Pfaender, S.; Mangus, K.; Reddy, P.P.; Rankovic, V.; Schmeisser, M.J.; et al. Zinc deficiency dysregulates the synaptic ProSAP/Shank scaffold and might contribute to autism spectrum disorders. Brain 2014, 137, 137–152. [Google Scholar] [CrossRef] [Green Version]
- Pfaender, S.; Föhr, K.; Lutz, A.K.; Putz, S.; Achberger, K.; Linta, L.; Liebau, S.; Boeckers, T.M.; Grabrucker, A.M. Cellular Zinc Homeostasis Contributes to Neuronal Differentiation in Human Induced Pluripotent Stem Cells. Neural. Plast. 2016, 2016, 3760702. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Xu, J.; Bezanilla, M.; van Huizen, R.; Derin, R.; Li, M. Differential stimulation of PKC phosphorylation of potassium channels by ZIP1 and ZIP2. Science 1999, 285, 1565–1569. [Google Scholar] [CrossRef]
- Colvin, R.A.; Lai, B.; Holmes, W.R.; Lee, D. Understanding metal homeostasis in primary cultured neurons. Studies using single neuron subcellular and quantitative metallomics. Metallomics 2015, 7, 1111–1123. [Google Scholar] [CrossRef]
- Pérez-Clausell, J.; Danscher, G. Intravesicular localization of zinc in rat telencephalic boutons. A histochemical study. Brain Res. 1985, 337, 91–98. [Google Scholar] [CrossRef]
- Frederickson, C.J.; Howell, G.A.; Haigh, M.D.; Danscher, G. Zinc-containing fiber systems in the cochlear nuclei of the rat and mouse. Hear. Res. 1988, 36, 203–211. [Google Scholar] [CrossRef]
- Sharonova, I.N.; Vorobjev, V.S.; Haas, H.L. Interaction between copper and zinc at GABA(A) receptors in acutely isolated cerebellar Purkinje cells of the rat. Br. J. Pharmacol. 2000, 130, 851–856. [Google Scholar] [CrossRef] [Green Version]
- Wall, M.J. A role for zinc in cerebellar synaptic transmission? Cerebellum 2005, 4, 224–229. [Google Scholar] [CrossRef]
- Frederickson, C.J.; Cuajungco, M.P.; LaBuda, C.J.; Suh, S.W. Nitric oxide causes apparent release of zinc from presynaptic boutons. Neuroscience 2002, 115, 471–474. [Google Scholar] [CrossRef]
- Pfaender, S.; Sauer, A.K.; Hagmeyer, S.; Mangus, K.; Linta, L.; Liebau, S.; Bockmann, J.; Huguet, G.; Bourgeron, T.; Boeckers, T.M.; et al. Zinc deficiency and low enterocyte zinc transporter expression in human patients with autism related mutations in SHANK3. Sci. Rep. 2017, 7, 45190. [Google Scholar] [CrossRef] [Green Version]
- Clifton, N.E.; Trent, S.; Thomas, K.L.; Hall, J. Regulation and Function of Activity-Dependent Homer in Synaptic Plasticity. Mol. Neuropsychiatry 2019, 5, 147–161. [Google Scholar] [CrossRef]
- Zhang, C.; Sui, D.; Zhang, T.; Hu, J. Molecular Basis of Zinc-Dependent Endocytosis of Human ZIP4 Transceptor. Cell Rep. 2020, 31, 107582. [Google Scholar] [CrossRef]
- Frederickson, C.J.; Suh, S.W.; Silva, D.; Frederickson, C.J.; Thompson, R.B. Importance of zinc in the central nervous system: The zinc-containing neuron. J. Nutr. 2000, 130, 1471–1483. [Google Scholar] [CrossRef]
- Frederickson, C.J.; Giblin, L.J., III; Balaji, R.V.; Masalha, R.; Frederickson, C.J.; Zeng, Y.; Lopez, E.V.; Koh, J.Y.; Chorin, U.; Besser, L.; et al. Synaptic release of zinc from brain slices: Factors governing release, imaging, and accurate calculation of concentration. J. Neurosci. Methods 2006, 154, 19–29. [Google Scholar] [CrossRef]
- Vogt, K.; Mellor, J.; Tong, G.; Nicoll, R. The actions of synaptically released zinc at hippocampal mossy fiber synapses. Neuron 2000, 26, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Vergnano, A.M.; Rebola, N.; Savtchenko, L.P.; Pinheiro, P.S.; Casado, M.; Kieffer, B.L.; Rusakov, D.A.; Mulle, C.; Paoletti, P. Zinc dynamics and action at excitatory synapses. Neuron 2014, 82, 1101–1114. [Google Scholar] [CrossRef] [Green Version]
- Grabrucker, A.M.; Knight, M.J.; Proepper, C.; Bockmann, J.; Joubert, M.; Rowan, M.; Nienhaus, G.U.; Garner, C.C.; Bowie, J.U.; Kreutz, M.R.; et al. Concerted action of zinc and ProSAP/Shank in synaptogenesis and synapse maturation. EMBO J. 2011, 30, 569–581. [Google Scholar] [CrossRef] [Green Version]
- Jan, H.H.; Chen, I.T.; Tsai, Y.Y.; Chang, Y.C. Structural role of zinc ions bound to postsynaptic densities. J. Neurochem. 2002, 83, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Gundelfinger, E.D.; Boeckers, T.M.; Baron, M.K.; Bowie, J.U. A role for zinc in postsynaptic density asSAMbly and plasticity? Trends Biochem. Sci. 2006, 31, 366–373. [Google Scholar] [CrossRef]
- Tao-Cheng, J.H.; Toy, D.; Winters, C.A.; Reese, T.S.; Dosemeci, A. Zinc Stabilizes Shank3 at the Postsynaptic Density of Hippocampal Synapses. PLoS ONE 2016, 11, e0153979. [Google Scholar] [CrossRef] [Green Version]
- De Benedictis, C.A.; Vilella, A.; Grabrucker, A.M. The Role of Trace Metals in Alzheimer’s Disease. In Alzheimer’s Disease; Wisniewski, T., Ed.; Codon Publications: Brisbane, Australia, 2019; Chapter 6. [Google Scholar] [CrossRef] [Green Version]
- Grabrucker, A.M.; Schmeisser, M.J.; Udvardi, P.T.; Arons, M.; Schoen, M.; Woodling, N.S.; Andreasson, K.I.; Hof, P.R.; Buxbaum, J.D.; Garner, C.C.; et al. Amyloid beta protein-induced zinc sequestration leads to synaptic loss via dysregulation of the ProSAP2/Shank3 scaffold. Mol. Neurodegener. 2011, 6, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilella, A.; Daini, E.; De Benedictis, C.A.; Grabrucker, A.M. Targeting Metal Homeostasis as a Therapeutic Strategy for Alzheimer’s Disease. In Alzheimer’s Disease: Drug Discovery; Huang, X., Ed.; Exon Publications: Brisbane, Australia, 2020; Chapter 5. [Google Scholar] [CrossRef]
- Grabrucker, A.; Vaida, B.; Bockmann, J.; Boeckers, T.M. Synaptogenesis of hippocampal neurons in primary cell culture. Cell Tissue Res. 2009, 338, 333–341. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | ZnT | Zip |
|---|---|---|
| Neurons | ZnT1, ZnT2, ZnT3, ZnT4, ZnT5, ZnT6, ZnT7, ZnT8, ZnT10 | Zip1, Zip2, Zip4 |
| Astrocytes | ZnT1, ZnT4, ZnT5, ZnT6, ZnT7, ZnT9 | Zip1, Zip6, Zip8 1, Zip9, Zip10, Zip11, Zip13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Benedictis, C.A.; Haffke, C.; Hagmeyer, S.; Sauer, A.K.; Grabrucker, A.M. Expression Analysis of Zinc Transporters in Nervous Tissue Cells Reveals Neuronal and Synaptic Localization of ZIP4. Int. J. Mol. Sci. 2021, 22, 4511. https://doi.org/10.3390/ijms22094511
De Benedictis CA, Haffke C, Hagmeyer S, Sauer AK, Grabrucker AM. Expression Analysis of Zinc Transporters in Nervous Tissue Cells Reveals Neuronal and Synaptic Localization of ZIP4. International Journal of Molecular Sciences. 2021; 22(9):4511. https://doi.org/10.3390/ijms22094511
Chicago/Turabian StyleDe Benedictis, Chiara A., Claudia Haffke, Simone Hagmeyer, Ann Katrin Sauer, and Andreas M. Grabrucker. 2021. "Expression Analysis of Zinc Transporters in Nervous Tissue Cells Reveals Neuronal and Synaptic Localization of ZIP4" International Journal of Molecular Sciences 22, no. 9: 4511. https://doi.org/10.3390/ijms22094511
APA StyleDe Benedictis, C. A., Haffke, C., Hagmeyer, S., Sauer, A. K., & Grabrucker, A. M. (2021). Expression Analysis of Zinc Transporters in Nervous Tissue Cells Reveals Neuronal and Synaptic Localization of ZIP4. International Journal of Molecular Sciences, 22(9), 4511. https://doi.org/10.3390/ijms22094511