
The Effect of Space Travel on Bone Metabolism: Considerations on Today’s Major Challenges and Advances in Pharmacology



Abstract
:1. Introduction
2. Bone Loss and Osteoporosis
3. Countermeasures
3.1. Diet Supplements
3.2. Physical Exercise
3.3. Pharmacological Countermeasures
3.3.1. Antiresorptive Agents
3.3.2. Anabolic Agents
3.3.3. Combination and Sequential Therapies
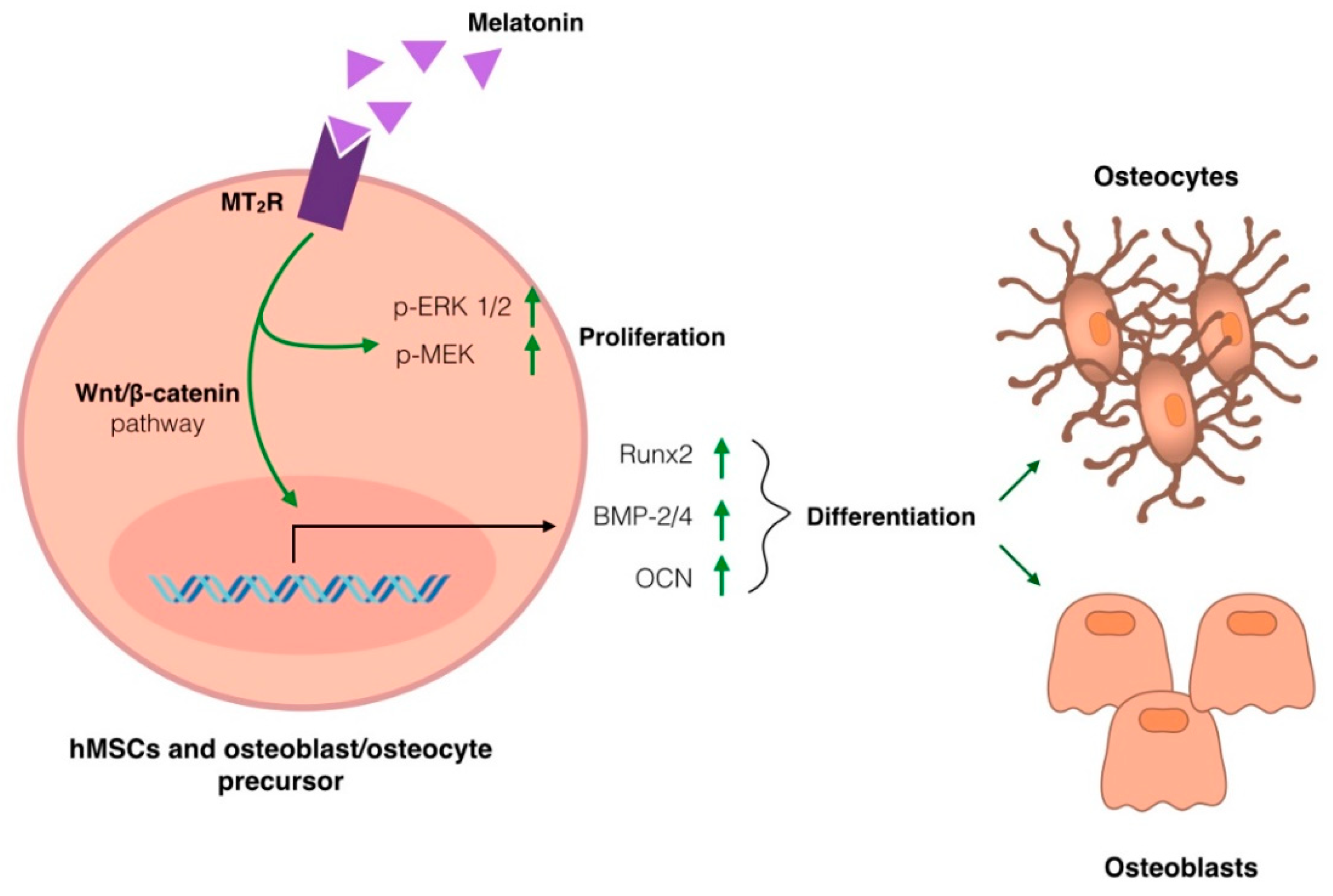
3.4. Melatonin
4. Fractures and Bone Healing
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Grimm, D.; Grosse, J.; Wehland, M.; Mann, V.; Reseland, J.E.; Sundaresan, A.; Corydon, T.J. The impact of microgravity on bone in humans. Bone 2016, 87, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.K. Cosmic rays: Are air crew at risk? Occup. Environ. Med. 2002, 59, 428–432. [Google Scholar] [CrossRef]
- Anken, R.H.; Rahmann, H. Gravitational Zoology: How Animals Use and Cope with Gravity. In Astrobiology; Horneck, G., Baumstark-Khan, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 315–333. [Google Scholar] [CrossRef]
- Tanaka, K.; Nishimura, N.; Kawai, Y. Adaptation to microgravity, deconditioning, and countermeasures. J. Physiol. Sci. 2016, 67, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Hinghofer-Szalkay, H. Gravity, the Hydrostatic Indifference Concept and the Cardiovascular System. Eur. J. Appl. Physiol. 2011, 111, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.; Wu, H.; Choi, J.U.; Rowan, A.E.; Zhang, H.; Poole, K.; Lauko, J.; Chou, J. Modeling the Impact of Microgravity at the Cellular Level: Implications for Human Disease. Front. Cell Dev. Biol. 2020, 8, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijk, D.-J.; Neri, D.F.; Wyatt, J.K.; Ronda, J.M.; Riel, E.; Cecco, A.R.-D.; Hughes, R.J.; Elliott, A.R.; Prisk, G.K.; West, J.B.; et al. Sleep, performance, circadian rhythms, and light-dark cycles during two space shuttle flights. Am. J. Physiol. Integr. Comp. Physiol. 2001, 281, R1647–R1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigoriev, A.I.; Egorov, A.D. The Effects of Prolonged Spaceflights on the Human Body. Adv. Space Biol. Med. 1991, 1, 1–35. [Google Scholar] [CrossRef]
- Carvil, P.; Baptista, R.; Russomano, T. The human body in a microgravity environment: Long term adaptations and countermeasures. Aviat. Focus 2013, 4, 10–22. [Google Scholar]
- Strollo, F.; Gentile, S.; Strollo, G.; Mambro, A.; Vernikos, J. Recent Progress in Space Physiology and Aging. Front. Physiol. 2018, 9, 1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strollo, F.; Vernikos, J. Aging-like metabolic and adrenal changes in microgravity: State of the art in preparation for Mars. Neurosci. Biobehav. Rev. 2021, 126, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, J.W.; Rummel, J.D. Long-term effects of microgravity and possible countermeasures. Adv. Space Res. 1992, 12, 281–284. [Google Scholar] [CrossRef]
- Monici, M.; Fusi, F.; Paglierani, M.; Marziliano, N.; Cogoli, A.; Pratesi, R.; Bernabei, P.A. Modeled gravitational unloading triggers differentiation and apoptosis in preosteoclastic cells. J. Cell. Biochem. 2006, 98, 65–80. [Google Scholar] [CrossRef]
- Di, S.; Tian, Z.; Qian, A.; Li, J.; Wu, J.; Wang, Z.; Zhang, D.; Yin, D.; Brandi, M.L.; Shang, P. Large gradient high magnetic field affects FLG29.1 cells differentiation to form osteoclast-like cells. Int. J. Radiat. Biol. 2012, 88, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Li, R.; Su, P.; Arfat, Y.; Zhang, G.; Shang, P.; Qian, A. Response and adaptation of bone cells to simulated microgravity. Acta Astronaut. 2014, 104, 396–408. [Google Scholar] [CrossRef]
- Nabavi, N.; Khandani, A.; Camirand, A.; Harrison, R.E. Effects of microgravity on osteoclast bone resorption and osteoblast cytoskeletal organization and adhesion. Bone 2011, 49, 965–974. [Google Scholar] [CrossRef]
- Wysolmerski, J.J. Osteocytic osteolysis: Time for a second look? BoneKEy Rep. 2012, 1, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallas, S.L.; Prideaux, M.; Bonewald, L.F. The Osteocyte: An Endocrine Cell … and More. Endocr. Rev. 2013, 34, 658–690. [Google Scholar] [CrossRef] [Green Version]
- Bloomfield, S.A.; Martinez, D.A.; Boudreaux, R.D.; Mantri, A.V. Microgravity Stress: Bone and Connective Tissue. In Comprehensive Physiology; Terjung, R., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 645–686. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Kang, H.; Liu, W.; Liu, P.; Zhang, J.; Harris, S.E.; Wu, D. Sclerostin Binds to LRP5/6 and Antagonizes Canonical Wnt Signaling. J. Biol. Chem. 2005, 280, 19883–19887. [Google Scholar] [CrossRef] [Green Version]
- Regard, J.B.; Zhong, Z.; Williams, B.O.; Yang, Y. Wnt Signaling in Bone Development and Disease: Making Stronger Bone with Wnts. Cold Spring Harb. Perspect. Biol. 2012, 4, a007997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, A.; Vayssière, B.; Garcia, T.; Newell, W.; Baron, R.; Roman-Roman, S.; Rawadi, G. Gene array analysis of Wnt-regulated genes in C3H10T1/2 cells. Bone 2005, 36, 585–598. [Google Scholar] [CrossRef]
- Macaulay, T.R.; Siamwala, J.H.; Hargens, A.R.; Macias, B.R. Thirty days of spaceflight does not alter murine calvariae structure despite increased Sost expression. Bone Rep. 2017, 7, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Bloomfield, S.A. Disuse Osteopenia. Curr. Osteoporos. Rep. 2010, 8, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Sun, L.-W.; Liang, M.; Wang, X.-N.; Fan, Y.-B. The Response of wnt/ß-Catenin Signaling Pathway in Osteocytes Under Simulated Microgravity. Microgravity Sci. Technol. 2015, 27, 473–483. [Google Scholar] [CrossRef]
- Blaber, E.; Sato, K.; Almeida, E.A.C. Stem Cell Health and Tissue Regeneration in Microgravity. Stem Cells Dev. 2014, 23 (Suppl. 1), 73–78. [Google Scholar] [CrossRef] [Green Version]
- Blaber, E.; Dvorochkin, N.; Torres, M.; Yousuf, R.; Burns, B.; Globus, R.; Almeida, E. Mechanical unloading of bone in microgravity reduces mesenchymal and hematopoietic stem cell-mediated tissue regeneration. Stem Cell Res. 2014, 13, 181–201. [Google Scholar] [CrossRef] [Green Version]
- Blaber, E.A.; Finkelstein, H.; Dvorochkin, N.; Sato, K.Y.; Yousuf, R.; Burns, B.P.; Globus, R.K.; Almeida, E.A. Microgravity Reduces the Differentiation and Regenerative Potential of Embryonic Stem Cells. Stem Cells Dev. 2015, 24, 2605–2621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibonga, J.D. Spaceflight-induced Bone Loss: Is there an Osteoporosis Risk? Curr. Osteoporos. Rep. 2013, 11, 92–98. [Google Scholar] [CrossRef]
- Vico, L.; Collet, P.; Guignandon, A.; Lafage-Proust, M.-H.; Thomas, T.; Rehailia, M.; Alexandre, C. Effects of long-term microgravity exposure on cancellous and cortical weight-bearing bones of cosmonauts. Lancet 2000, 355, 1607–1611. [Google Scholar] [CrossRef]
- Khan, M.; Jose, A.; Sharma, S. Physiology, Parathyroid Hormone. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Smith, S.M.; Heer, M.; Shackelford, L.C.; Sibonga, J.D.; Spatz, J.; Pietrzyk, R.A.; Hudson, E.K.; Zwart, S.R. Bone metabolism and renal stone risk during International Space Station missions. Bone 2015, 81, 712–720. [Google Scholar] [CrossRef]
- Smith, J.K. Osteoclasts and Microgravity. Life 2020, 10, 207. [Google Scholar] [CrossRef]
- Iwase, S.; Nishimura, N.; Mano, T. Osteoporosis in Spaceflight. In Topics in Osteoporosis; Valds-Flores, M., Ed.; InTech: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Hodges, J.K.; Cao, S.; Cladis, D.P.; Weaver, C.M. Lactose Intolerance and Bone Health: The Challenge of Ensuring Adequate Calcium Intake. Nutrients 2019, 11, 718. [Google Scholar] [CrossRef] [Green Version]
- Sunyecz, J. The use of calcium and vitamin D in the management of osteoporosis. Ther. Clin. Risk Manag. 2008, 4, 827–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, T.; Van Loon, J.J.; Bloomfield, S.; Vico, L.; Chopard, A.; Rittweger, J.; Kyparos, A.; Blottner, D.; Vuori, I.; Gerzer, R.; et al. Towards human exploration of space: The THESEUS review series on muscle and bone research priorities. NPJ Microgravity 2017, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.M.; Heer, M.A.; Shackelford, L.C.; Sibonga, J.D.; Ploutz-Snyder, L.; Zwart, S.R. Benefits for bone from resistance exercise and nutrition in long-duration spaceflight: Evidence from biochemistry and densitometry. J. Bone Miner. Res. 2012, 27, 1896–1906. [Google Scholar] [CrossRef]
- Lambrinoudaki, I.; Christodoulakos, G.; Botsis, D. Bisphosphonates. Ann. N. Y. Acad. Sci. 2006, 1092, 397–402. [Google Scholar] [CrossRef]
- Drake, M.T.; Clarke, B.L.; Khosla, S. Bisphosphonates: Mechanism of Action and Role in Clinical Practice. Mayo Clin. Proc. 2008, 83, 1032–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leblanc, A.; Matsumoto, T.; Jones, J.; Shapiro, J.; Lang, T.; Shackelford, L.; Smith, S.M.; Evans, H.; Spector, E.; Ploutz-Snyder, R.; et al. Bisphosphonates as a supplement to exercise to protect bone during long-duration spaceflight. Osteoporos. Int. 2013, 24, 2105–2114. [Google Scholar] [CrossRef]
- Kohli, S.S.; Kohli, V.S. Role of RANKL-RANK/osteoprotegerin molecular complex in bone remodeling and its immunopathologic implications. Ind. J. Endocrinol. Metab. 2011, 15, 175–181. [Google Scholar] [CrossRef]
- Deeks, E.D. Denosumab: A Review in Postmenopausal Osteoporosis. Drugs Aging 2018, 35, 163–173. [Google Scholar] [CrossRef]
- Cheng, C.; Wentworth, K.; Shoback, D.M. New Frontiers in Osteoporosis Therapy. Annu. Rev. Med. 2020, 71, 277–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Saag, K.G.; Curtis, J.R. Long-term Safety Concerns of Antiresorptive Therapy. Rheum. Dis. Clin. N. Am. 2011, 37, 387–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kast, J.; Yu, Y.; Seubert, C.N.; Wotring, V.E.; Derendorf, H. Drugs in space: Pharmacokinetics and pharmacodynamics in astronauts. Eur. J. Pharm. Sci. 2017, 109, S2–S8. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Hodges, M.; Yurgin, N.; Boye, K.S. Impact of dose frequency on compliance and health outcomes: A literature review (1966–2006). Expert Rev. Pharm. Outcomes Res. 2007, 7, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Baykan, E.K.; Saygılı, L.F.; Erdogan, M.; Cetinkalp, S.; Ozgen, A.G.; Yilmaz, C. Efficacy of zoledronic acid treatment in Paget disease of bone. Osteoporos. Int. 2014, 25, 2221–2223. [Google Scholar] [CrossRef] [PubMed]
- Maricic, M. The role of zoledronic acid in the management of osteoporosis. Clin. Rheumatol. 2010, 29, 1079–1084. [Google Scholar] [CrossRef] [PubMed]
- Wein, M.N.; Kronenberg, H.M. Regulation of Bone Remodeling by Parathyroid Hormone. Cold Spring Harb. Perspect. Med. 2018, 8, a031237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satterwhite, J.; Heathman, M.; Miller, P.D.; Marín, F.; Glass, E.V.; Dobnig, H. Pharmacokinetics of Teriparatide (rhPTH[1–34]) and Calcium Pharmacodynamics in Postmenopausal Women with Osteoporosis. Calcif. Tissue Int. 2010, 87, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minisola, S.; Cipriani, C.; Della Grotta, G.; Colangelo, L.; Occhiuto, M.; Biondi, P.; Sonato, C.; Vigna, E.; Cilli, M.; Pepe, J. Update on the safety and efficacy of teriparatide in the treatment of osteoporosis. Ther. Adv. Musculoskelet. Dis. 2019, 11, 1759720X1987799. [Google Scholar] [CrossRef] [Green Version]
- Henriksen, K.; Andersen, J.; Riis, B.; Mehta, N.; Tavakkol, R.; Alexandersen, P.; Byrjalsen, I.; Valter, I.; Nedergaard, B.; Teglbjærg, C.; et al. Evaluation of the efficacy, safety and pharmacokinetic profile of oral recombinant human parathyroid hormone [rhPTH(1–31)NH2] in postmenopausal women with osteoporosis. Bone 2013, 53, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Hämmerle, S.P.; Mindeholm, L.; Launonen, A.; Kiese, B.; Loeffler, R.; Harfst, E.; Azria, M.; Arnold, M.; John, M.R. The single dose pharmacokinetic profile of a novel oral human parathyroid hormone formulation in healthy postmenopausal women. Bone 2012, 50, 965–973. [Google Scholar] [CrossRef]
- Altaani, B.M.; Almaaytah, A.M.; Dadou, S.; Alkhamis, K.; Daradka, M.H.; Hananeh, W. Oral Delivery of Teriparatide Using a Nanoemulsion System: Design, in Vitro and in Vivo Evaluation. Pharm. Res. 2020, 37, 80. [Google Scholar] [CrossRef] [PubMed]
- Zerwekh, J.E. Nutrition and renal stone disease in space. Nutrition 2002, 18, 857–863. [Google Scholar] [CrossRef]
- Liakopoulos, V.; Leivaditis, K.; Eleftheriadis, T.; Dombros, N. The kidney in space. Int. Urol. Nephrol. 2012, 44, 1893–1901. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.C.; Bilezikian, J.P. Parathyroid hormone: Anabolic and catabolic actions on the skeleton. Curr. Opin. Pharmacol. 2015, 22, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, J.S.; Wyland, J.J.; Leder, B.Z.; Burnett-Bowie, S.-A.M.; Lee, H.; Jüppner, H.; Neer, R.M. Effects of Teriparatide Retreatment in Osteoporotic Men and Women. J. Clin. Endocrinol. Metab. 2009, 94, 2495–2501. [Google Scholar] [CrossRef]
- Russow, G.; Jahn, D.; Appelt, J.; Märdian, S.; Tsitsilonis, S.; Keller, J. Anabolic Therapies in Osteoporosis and Bone Regeneration. Int. J. Mol. Sci. 2018, 20, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pazianas, M. Anabolic effects of PTH and the ‘Anabolic Window’. Trends Endocrinol. Metab. 2015, 26, 111–113. [Google Scholar] [CrossRef]
- Manzey, D.; Lorenz, B.; Poljakov, V. Mental performance in extreme environments: Results from a performance monitoring study during a 438-day spaceflight. Ergonomics 1998, 41, 537–559. [Google Scholar] [CrossRef]
- Leder, B.Z.; Tsai, J.N.; Uihlein, A.V.; Burnett-Bowie, S.-A.M.; Zhu, Y.; Foley, K.; Lee, H.; Neer, R.M. Two Years of Denosumab and Teriparatide Administration in Postmenopausal Women with Osteoporosis (The DATA Extension Study): A Randomized Controlled Trial. J. Clin. Endocrinol. Metab. 2014, 99, 1694–1700. [Google Scholar] [CrossRef] [PubMed]
- Leder, B.Z.; Tsai, J.N.; Uihlein, A.V.; Wallace, P.M.; Lee, H.; Neer, R.M.; Burnett-Bowie, S.-A.M. Denosumab and teriparatide transitions in postmenopausal osteoporosis (the DATA-Switch study): Extension of a randomised controlled trial. Lancet 2015, 386, 1147–1155. [Google Scholar] [CrossRef] [Green Version]
- Park, K.-H.; Kang, J.W.; Lee, E.-M.; Kim, J.S.; Rhee, Y.H.; Kim, M.; Jeong, S.J.; Park, Y.G.; Kim, S.H. Melatonin promotes osteoblastic differentiation through the BMP/ERK/Wnt signaling pathways. J. Pineal Res. 2011, 51, 187–194. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.-X.; Fuentes-Broto, L. Melatonin: A Multitasking Molecule. Prog. Brain Res. 2010, 181, 127–151. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Paredes, S.D.; Fuentes-Broto, L. Beneficial effects of melatonin in cardiovascular disease. Ann. Med. 2010, 42, 276–285. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Ladizesky, M.G.; Boggio, V.; Cutrera, R.A.; Mautalen, C. Melatonin effects on bone: Experimental facts and clinical perspectives. J. Pineal Res. 2003, 34, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Maria, S.; Samsonraj, R.M.; Munmun, F.; Glas, J.; Silvestros, M.; Kotlarczyk, M.P.; Rylands, R.; Dudakovic, A.; Van Wijnen, A.J.; Enderby, L.T.; et al. Biological effects of melatonin on osteoblast/osteoclast cocultures, bone, and quality of life: Implications of a role for MT2 melatonin receptors, MEK1/2, and MEK5 in melatonin-mediated osteoblastogenesis. J. Pineal Res. 2018, 64, e12465. [Google Scholar] [CrossRef] [PubMed]
- Amstrup, A.K.; Sikjaer, T.; Heickendorff, L.; Mosekilde, L.; Rejnmark, L. Melatonin improves bone mineral density at the femoral neck in postmenopausal women with osteopenia: A randomized controlled trial. J. Pineal Res. 2015, 59, 221–229. [Google Scholar] [CrossRef]
- Kearns, A.E.; Khosla, S.; Kostenuik, P.J. Receptor Activator of Nuclear Factor κB Ligand and Osteoprotegerin Regulation of Bone Remodeling in Health and Disease. Endocr. Rev. 2007, 29, 155–192. [Google Scholar] [CrossRef]
- Kondo, Y.; Irie, K.; Ikegame, M.; Ejiri, S.; Hanada, K.; Ozawa, H. Role of stromal cells in osteoclast differentiation in bone marrow. J. Bone Miner. Metab. 2001, 19, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Kotlarczyk, M.P.; Lassila, H.C.; O’Neil, C.K.; D’Amico, F.; Enderby, L.T.; Witt-Enderby, P.A.; Balk, J.L. Melatonin osteoporosis prevention study (MOPS): A randomized, double-blind, placebo-controlled study examining the effects of melatonin on bone health and quality of life in perimenopausal women. J. Pineal Res. 2012, 52, 414–426. [Google Scholar] [CrossRef] [PubMed]
- Ikegame, M.; Hattori, A.; Tabata, M.J.; Kitamura, K.; Tabuchi, Y.; Furusawa, Y.; Maruyama, Y.; Yamamoto, T.; Sekiguchi, T.; Matsuoka, R.; et al. Melatonin is a potential drug for the prevention of bone loss during space flight. J. Pineal Res. 2019, 67, e12594. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Ikegame, M.; Igarashi-Migitaka, J.; Maruyama, Y.; Suzuki, N.; Hattori, A. Suppressive effect of melatonin on osteoclast function via osteocyte calcitonin. J. Endocrinol. 2019, 242, 13–23. [Google Scholar] [CrossRef]
- Naot, D.; Musson, D.S.; Cornish, J. The Activity of Peptides of the Calcitonin Family in Bone. Physiol. Rev. 2019, 99, 781–805. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Guo, J.; Kanwal, Z.; Wu, M.; Lv, X.; Ibrahim, N.A.; Li, P.; Buabeid, M.A.; Arafa, E.-S.A.; Sun, Q. Calcitonin and Bone Physiology: In Vitro, In Vivo, and Clinical Investigations. Int. J. Endocrinol. 2020, 2020, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Wimalawansa, S.J. Calcitonin. In Osteoporosis in Men; Elsevier: Amsterdam, The Netherlands, 2010; pp. 653–666. [Google Scholar] [CrossRef]
- Masi, L.; Brandi, M.L. Calcitonin and calcitonin receptors. Clin. Cases Miner. Bone Metab. 2007, 4, 117–122. [Google Scholar] [PubMed]
- McLaughlin, M.B.; Jialal, I. Calcitonin. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Lehenkari, P.; Hentunen, T.A.; Laitala-Leinonen, T.; Tuukkanen, J.; Väänänen, H. Carbonic Anhydrase II Plays a Major Role in Osteoclast Differentiation and Bone Resorption by Effecting the Steady State Intracellular pH and Ca2+. Exp. Cell Res. 1998, 242, 128–137. [Google Scholar] [CrossRef]
- Guo, J.-H.; Qu, W.-M.; Chen, S.-G.; Chen, X.-P.; Lv, K.; Huang, Z.-L.; Wu, Y.-L. Keeping the right time in space: Importance of circadian clock and sleep for physiology and performance of astronauts. Mil. Med Res. 2014, 1, 23. [Google Scholar] [CrossRef] [Green Version]
- Wotring, V. Space Pharmacology: How Space Affects Pharmacology. In Drug Discovery and Evaluation: Methods in Clinical Pharmacology; Hock, F.J., Gralinski, M.R., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–13. [Google Scholar] [CrossRef]
- Nelson, E.S.; Lewandowski, B.; Licata, A.; Myers, J.G. Development and Validation of a Predictive Bone Fracture Risk Model for Astronauts. Ann. Biomed. Eng. 2009, 37, 2337–2359. [Google Scholar] [CrossRef]
- Dadwal, U.C.; Maupin, K.A.; Zamarioli, A.; Tucker, A.; Harris, J.S.; Fischer, J.P.; Rytlewski, J.D.; Scofield, D.C.; Wininger, A.E.; Bhatti, F.U.R.; et al. The effects of spaceflight and fracture healing on distant skeletal sites. Sci. Rep. 2019, 9, 11419. [Google Scholar] [CrossRef] [Green Version]
- Schindeler, A.; McDonald, M.M.; Bokko, P.; Little, D.G. Bone remodeling during fracture repair: The cellular picture. Semin. Cell Dev. Biol. 2008, 19, 459–466. [Google Scholar] [CrossRef]
- Deschaseaux, F.; Sensébé, L.; Heymann, D. Mechanisms of bone repair and regeneration. Trends Mol. Med. 2009, 15, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Saran, U.; Piperni, S.G.; Chatterjee, S. Role of angiogenesis in bone repair. Arch. Biochem. Biophys. 2014, 561, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Usas, A.; Olshanski, A.; Ho, A.M.; Gearhart, B.; Cooper, G.M.; Huard, J. VEGF Improves, Whereas sFlt1 Inhibits, BMP2-Induced Bone Formation and Bone Healing Through Modulation of Angiogenesis. J. Bone Miner. Res. 2005, 20, 2017–2027. [Google Scholar] [CrossRef] [PubMed]
- Brandi, M.L.; Collin-Osdoby, P. Vascular Biology and the Skeleton. J. Bone Miner. Res. 2006, 21, 183–192. [Google Scholar] [CrossRef]
- Maes, C.; Kobayashi, T.; Selig, M.K.; Torrekens, S.; Roth, S.I.; Mackem, S.; Carmeliet, G.; Kronenberg, H.M. Osteoblast Precursors, but Not Mature Osteoblasts, Move into Developing and Fractured Bones along with Invading Blood Vessels. Dev. Cell 2010, 19, 329–344. [Google Scholar] [CrossRef] [Green Version]
- Morbidelli, L.; Monici, M.; Marziliano, N.; Cogoli, A.; Fusi, F.; Waltenberger, J.; Ziche, M. Simulated hypogravity impairs the angiogenic response of endothelium by up-regulating apoptotic signals. Biochem. Biophys. Res. Commun. 2005, 334, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Cialdai, F.; Vignali, L.; Morbidelli, L.; Colciago, A.; Celotti, F.; Santi, A.; Caselli, A.; Cirri, P.; Monici, M. Modeled Microgravity Affects Fibroblast Functions Related to Wound Healing. Microgravity Sci. Technol. 2017, 29, 121–132. [Google Scholar] [CrossRef]
- Maier, J.A.M.; Cialdai, F.; Monici, M.; Morbidelli, L. The Impact of Microgravity and Hypergravity on Endothelial Cells. BioMed Res. Int. 2015, 2015, 1–13. [Google Scholar] [CrossRef]
- Androjna, C.; McCabe, N.P.; Cavanagh, P.R.; Midura, R.J. Effects of Spaceflight and Skeletal Unloading on Bone Fracture Healing. Clin. Rev. Bone Miner. Metab. 2011, 10, 61–70. [Google Scholar] [CrossRef]
- Gadomski, B.C.; McGilvray, K.C.; Easley, J.T.; Palmer, R.H.; Santoni, B.G.; Puttlitz, C.M. Partial gravity unloading inhibits bone healing responses in a large animal model. J. Biomech. 2014, 47, 2836–2842. [Google Scholar] [CrossRef]
- Kirchen, M.E.; O’Connor, K.M.; Gruber, H.E.; Sweeney, J.R.; Fras, I.A.; Stover, S.J.; Sarmiento, A.; Marshall, G.J. Effects of microgravity on bone healing in a rat fibular osteotomy model. Clin. Orthop. Relat. Res. 1995, 318, 231–242. [Google Scholar]
- Ansari, M. Bone tissue regeneration: Biology, strategies and interface studies. Prog. Biomater. 2019, 8, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Mittermayr, R.; Haffner, N.; Feichtinger, X.; Schaden, W. The role of shockwaves in the enhancement of bone repair—From basic principles to clinical application. Injury 2021. [Google Scholar] [CrossRef] [PubMed]
- Buza, J.A. Bone healing in 2016. Clin. Cases Miner. Bone Metab. 2016, 13, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Sella, V.R.G.; Bomfim, F.R.C.D.; Machado, P.C.D.; Morsoleto, M.J.M.D.S.; Chohfi, M.; Plapler, H. Effect of low-level laser therapy on bone repair: A randomized controlled experimental study. Lasers Med. Sci. 2015, 30, 1061–1068. [Google Scholar] [CrossRef]
- Genah, S.; Cialdai, F.; Ciccone, V.; Sereni, E.; Morbidelli, L.; Monici, M. Effect of NIR Laser Therapy by MLS-MiS Source on Fibroblast Activation by Inflammatory Cytokines in Relation to Wound Healing. Biomedicines 2021, 9, 307. [Google Scholar] [CrossRef] [PubMed]
- Iaquinta, M.R.; Mazzoni, E.; Bononi, I.; Rotondo, J.C.; Mazziotta, C.; Montesi, M.; Sprio, S.; Tampieri, A.; Tognon, M.; Martini, F. Adult Stem Cells for Bone Regeneration and Repair. Front. Cell Dev. Biol. 2019, 7, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haffner-Luntzer, M. Experimental Agents to Improve Fracture Healing: Utilizing the WNT Signaling Pathway. Injury 2020. [Google Scholar] [CrossRef]
- Gregory, C.A.; Green, A.; Lee, N.; Rao, A.; Gunn, W. The Promise of Canonical Wnt Signaling Modulators in Enhancing Bone Repair. Drug News Perspect. 2006, 19, 445–452. [Google Scholar] [CrossRef]
- Garrison, K.R.; Shemilt, I.; Donell, S.; Ryder, J.J.; Mugford, M.; Harvey, I.; Song, F.; Alt, V. Bone morphogenetic protein (BMP) for fracture healing in adults. Cochrane Database Syst. Rev. 2010, 2010. [Google Scholar] [CrossRef]
- Dumic-Cule, I.; Peric, M.; Kucko, L.; Grgurevic, L.; Pecina, M.; Vukicevic, S. Bone morphogenetic proteins in fracture repair. Int. Orthop. 2018, 42, 2619–2626. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Activity | Class | Drug | Dosage | Administration |
|---|---|---|---|---|
| Anti-resorptive agents | Bisphosphonates | Alendronate | 70 mg/week | Oral |
| Risedronate | 35–75 mg/week | Oral | ||
| Ibandronate | 150 mg/month | Oral | ||
| Pamidronate | 60–90 mg/month | Intravenous | ||
| Zoledronic acid | 4–5 mg/year | Intravenous | ||
| Monoclonal antibodies | Denosumab | 60 mg/6 months | Subcutaneous | |
| Anabolic drugs | Teriparatide | 20 μg/day | Subcutaneous | |
| 2.5–5 mg/ day | Oral |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genah, S.; Monici, M.; Morbidelli, L. The Effect of Space Travel on Bone Metabolism: Considerations on Today’s Major Challenges and Advances in Pharmacology. Int. J. Mol. Sci. 2021, 22, 4585. https://doi.org/10.3390/ijms22094585
Genah S, Monici M, Morbidelli L. The Effect of Space Travel on Bone Metabolism: Considerations on Today’s Major Challenges and Advances in Pharmacology. International Journal of Molecular Sciences. 2021; 22(9):4585. https://doi.org/10.3390/ijms22094585
Chicago/Turabian StyleGenah, Shirley, Monica Monici, and Lucia Morbidelli. 2021. "The Effect of Space Travel on Bone Metabolism: Considerations on Today’s Major Challenges and Advances in Pharmacology" International Journal of Molecular Sciences 22, no. 9: 4585. https://doi.org/10.3390/ijms22094585
APA StyleGenah, S., Monici, M., & Morbidelli, L. (2021). The Effect of Space Travel on Bone Metabolism: Considerations on Today’s Major Challenges and Advances in Pharmacology. International Journal of Molecular Sciences, 22(9), 4585. https://doi.org/10.3390/ijms22094585