
Circadian Rhythms in Legumes: What Do We Know and What Else Should We Explore?

 , and
, and
Abstract
:
1. Introduction
2. Defining the Important Components of Clock Research
2.1. Photoperiodic Flowering
2.2. Growth and Metabolism
2.3. Hormone Signaling
2.4. Biotic and Abiotic Stress
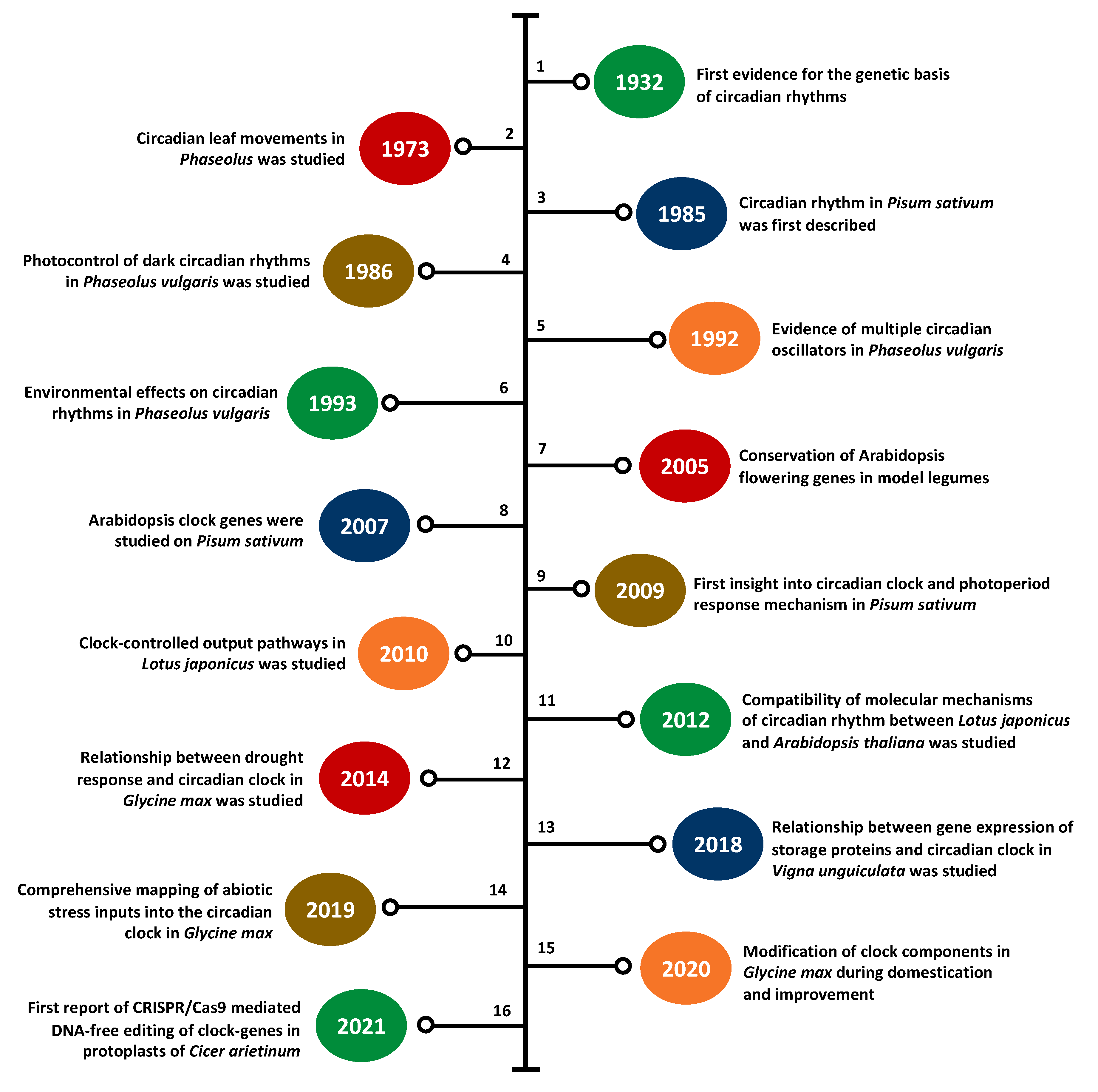
3. Legume Clock Research at a Glance
3.1. Clock Research in Model Legumes
3.1.1. Barrel Medic
3.1.2. Birds-Foot Trefoil
3.1.3. Soybean
3.1.4. Common Pea
3.2. Clock Research in Underutilized Legumes
4. Concluding Remarks and Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McClung, C.R. The plant circadian oscillator. Biology 2019, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, D.; Shamim, M.; Kumar, M.; Mishra, A.; Maurya, R.; Sharma, D.; Pandey, P.; Singh, K. Role of circadian rhythm in plant system: An update from development to stress response. Environ. Exp. Bot. 2019, 162, 256–271. [Google Scholar] [CrossRef]
- McClung, C.R. Circadian clock components offer targets for crop domestication and improvement. Genes 2021, 12, 374. [Google Scholar] [CrossRef]
- Salmela, M.J.; Weinig, C. The fitness benefits of genetic variation in circadian clock regulation. Curr. Opin. Plant Biol. 2019, 49, 86–93. [Google Scholar] [CrossRef]
- Kim, J.A.; Kim, H.S.; Choi, S.H.; Jang, J.Y.; Jeong, M.J.; Lee, S.I. The importance of the circadian clock in regulating plant metabolism. Int. J. Mol. Sci. 2017, 18, 2680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Nusinow, D.A. Into the evening: Complex interactions in the Arabidopsis circadian clock. Trends Genet. 2016, 32, 674–686. [Google Scholar] [CrossRef]
- Sahruzaini, N.A.; Rejab, N.A.; Harikrishna, J.A.; Ikram, N.K.K.; Ismail, I.; Kugan, H.M.; Cheng, A. Pulse crop genetics for a sustainable future: Where we are now and where we should be heading. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.; Raai, M.N.; Zain, N.A.M.; Massawe, F.; Singh, A.; Wan, W.A.A.Q.I. In search of alternative proteins: Unlocking the potential of underutilized tropical legumes. Food Secur. 2019, 11, 1205–1215. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, G. Biofortification of pulses and legumes to enhance nutrition. Heliyon 2020, 6, e03682. [Google Scholar] [CrossRef] [PubMed]
- Smýkal, P.; Aubert, G.; Burstin, J.; Coyne, C.J.; Ellis, N.T.; Flavell, A.J.; Ford, R.; Hýbl, M.; Macas, J.; Neumann, P. Pea (Pisum sativum L.) in the genomic era. Agronomy 2012, 2, 74–115. [Google Scholar] [CrossRef]
- Inoue, K.; Araki, T.; Endo, M. Integration of input signals into the gene network in the plant circadian clock. Plant Cell Physiol. 2017, 58, 977–982. [Google Scholar] [CrossRef]
- Gil, K.E.; Park, C.M. Thermal adaptation and plasticity of the plant circadian clock. New Phytol. 2018, 221, 1215–1229. [Google Scholar] [CrossRef] [Green Version]
- Oakenfull, R.J.; Davis, S.J. Shining a light on the Arabidopsis circadian clock. Plant Cell Environ. 2017, 40, 2571–2585. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, S.E.; Kay, S.A. The plant circadian clock: From a simple timekeeper to a complex developmental manager. Cold Spring Harb. Perspect. Biol. 2016, 8, a027748. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.A. Retrograde signalling as an informant of circadian timing. New Phytol. 2019, 221, 1749–1753. [Google Scholar] [CrossRef] [Green Version]
- Ionescu, I.A.; Møller, B.L.; Sánchez-Pérez, R. Chemical control of flowering time. J. Exp. Bot. 2016, 68, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Hecht, V.; Foucher, F.; Ferrándiz, C.; Macknight, R.; Navarro, C.; Morin, J.; Vardy, M.E.; Ellis, N.; Beltrán, J.P.; Rameau, C.; et al. Conservation of Arabidopsis flowering genes in model legumes. Plant Physiol. 2005, 137, 1420–1434. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Krom, N.; Zhang, S.; Mysore, K.S.; Udvardi, M.; Wen, J. Enabling Reverse Genetics in Medicago truncatula using high-throughput sequencing for Tnt1 flanking sequence recovery. Methods Mol. Biol. 2017, 1610, 25–37. [Google Scholar]
- Magne, K.; George, J.; Tornero, A.B.; Broquet, B.; Madueño, F.; Andersen, S.U.; Ratet, P. Lotus japonicus NOOT-BOP-COCH-LIKE1is essential for nodule, nectary, leaf and flower development. Plant J. 2018, 94, 880–894. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Gill, U.S.; Nandety, R.S.; Kwon, S.; Mehta, P.; Dickstein, R.; Udvardi, M.K.; Mysore, K.S.; Wen, J. Genome-wide analysis of flanking sequences reveals that Tnt1 insertion is positively correlated with gene methylation in Medicago truncatula. Plant J. 2019, 98, 1106–1119. [Google Scholar] [CrossRef]
- Cronk, Q.; Ojeda, I.; Pennington, R.T. Legume comparative genomics: Progress in phylogenetics and phylogenomics. Curr. Opin. Plant Biol. 2006, 9, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Hecht, V.; Knowles, C.L.; Vander Schoor, J.K.; Liew, L.C.; Jones, S.E.; Lambert, M.J.; Weller, J.L. Pea LATE BLOOMER1 is a GIGANTEA ortholog with roles in photoperiodic flowering, deetiolation, and transcriptional regulation of circadian clock gene homologs. Plant Physiol. 2007, 144, 648–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weller, J.L.; Ortega, R. Genetic control of flowering time in legumes. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.; Hecht, V.F.; Picard, K.; Diwadkar, P.; Laurie, R.E.; Wen, J.; Mysore, K.; Macknight, R.C.; Weller, J.L. Isolation and functional analysis of CONSTANS-LIKE genes suggests that a central role for CONSTANS in flowering time control is not evolutionarily conserved in Medicago truncatula. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Hecht, V.; Laurie, R.E.; Schoor, J.K.V.; Ridge, S.; Knowles, C.L.; Liew, L.C.; Sussmilch, F.C.; Murfet, I.C.; Macknight, R.C.; Weller, J.L. The pea GIGAS Gene is a FLOWERING LOCUS T homolog necessary for graft-transmissible specification of flowering but not for responsiveness to photoperiod. Plant Cell 2011, 23, 147–161. [Google Scholar] [CrossRef] [Green Version]
- Dodd, A.N.; Belbin, F.E.; Frank, A.; Webb, A.A.R. Interactions between circadian clocks and photosynthesis for the temporal and spatial coordination of metabolism. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foo, M.; Somers, D.E.; Kim, P.-J. Kernel architecture of the genetic circuitry of the Arabidopsis circadian system. PLoS Comput. Biol. 2016, 12, e1004748. [Google Scholar] [CrossRef] [Green Version]
- Nagel, D.H.; Kay, S.A. Complexity in the wiring and regulation of plant circadian networks. Curr. Biol. 2012, 22, R648–R657. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.-J.; Wang, X.; Deng, Y.-R.; Li, J.-H.; Chen, W.; Chiang, J.Y.; Yang, J.-B.; Zheng, L. Nondestructive and intuitive determination of circadian chlorophyll rhythms in soybean leaves using multispectral imaging. Sci. Rep. 2015, 5, 11108. [Google Scholar] [CrossRef]
- Singh, V.K.; Rajkumar, M.S.; Garg, R.; Jain, M. Genome-wide identification and co-expression network analysis provide insights into the roles of auxin response factor gene family in chickpea. Sci. Rep. 2017, 7, 10895. [Google Scholar] [CrossRef]
- Zhuo, C.; Liang, L.; Zhao, Y.; Guo, Z.; Lu, S. A cold responsive ethylene responsive factor from Medicago falcata confers cold tolerance by up-regulation of polyamine turnover, antioxidant protection, and proline accumulation. Plant Cell Environ. 2018, 41, 2021–2032. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Wang, H.; Hong, L.; Xu, Y.; Zhao, Y.; Zhou, C. MtBZR1 plays an important role in nodule development in Medicago truncatula. Int. J. Mol. Sci. 2019, 20, 2941. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Debellé, F.; Gamas, P.; Frugier, F.; Brault, M. Diversification of cytokinin phosphotransfer signaling genes in Medicago truncatula and other legume genomes. BMC Genom. 2019, 20, 373. [Google Scholar] [CrossRef]
- Kong, Y.; Meng, Z.; Wang, H.; Wang, Y.; Zhang, Y.; Hong, L.; Liu, R.; Wang, M.; Zhang, J.; Han, L.; et al. Brassinosteroid homeostasis is critical for the functionality of the Medicago truncatula pulvinus. Plant Physiol. 2021. [Google Scholar] [CrossRef]
- Lyu, X.; Cheng, Q.; Qin, C.; Li, Y.; Xu, X.; Ji, R.; Mu, R.; Li, H.; Zhao, T.; Liu, J.; et al. GmCRY1s modulate gibberellin metabolism to regulate soybean shade avoidance in response to reduced blue light. Mol. Plant 2021, 14, 298–314. [Google Scholar] [CrossRef]
- Goodspeed, D.; Chehab, E.W.; Min-Venditti, A.; Braam, J.; Covington, M.F. Arabidopsis synchronizes jasmonate-mediated defense with insect circadian behavior. Proc. Natl. Acad. Sci. USA 2012, 109, 4674–4677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Martínez, J.L.; Gil, J. Light regulation of gibberellin biosynthesis and mode of action. J. Plant Growth Regul. 2001, 20, 354–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hisamatsu, T.; King, R.W.; Helliwell, C.A.; Koshioka, M. The involvement of gibberellin 20-oxidase genes in phytochrome-regulated petiole elongation of Arabidopsis. Plant Physiol. 2005, 138, 1106–1116. [Google Scholar] [CrossRef] [Green Version]
- Arana, M.V.; Marín-de la Rosa, N.; Maloof, J.N.; Blázquez, M.A.; Alabadí, D. Circadian oscillation of gibberellin signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 9292–9297. [Google Scholar] [CrossRef] [Green Version]
- Pokhilko, A.; Bou-Torrent, J.; Pulido, P.; Rodríguez-Concepción, M.; Ebenhöh, O. Mathematical modelling of the diurnal regulation of the MEP pathway in Arabidopsis. New Phytol. 2015, 206, 1075–1085. [Google Scholar] [CrossRef]
- Rawat, R.; Schwartz, J.; Jones, M.A.; Sairanen, I.; Cheng, Y.; Andersson, C.R.; Zhao, Y.; Ljung, K.; Harmer, S.L. REVEILLE1, a Myb-like transcription factor, integrates the circadian clock and auxin pathways. Proc. Natl. Acad. Sci. USA 2009, 106, 16883–16888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Mas, P. A functional connection between the circadian clock and hormonal timing in Arabidopsis. Genes 2018, 9, 567. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Cao, L.; Mwimba, M.; Zhou, Y.; Li, L.; Zhou, M.; Schnable, P.S.; O’Rourke, J.A.; Dong, X.; Wang, W. Comprehensive mapping of abiotic stress inputs into the soybean circadian clock. Proc. Natl. Acad. Sci. USA 2019, 116, 23840–23849. [Google Scholar] [CrossRef] [Green Version]
- Vezza, M.E.; Alderete, L.G.S.; Agostini, E.; Talano, M.A. Expression of circadian clock genes and diurnal oscillations of key physiological events in response to AsV and AsIII in soybean plants. Environ. Exp. Bot. 2020, 174. [Google Scholar] [CrossRef]
- Preuss, S.B.; Meister, R.; Xu, Q.; Urwin, C.P.; Tripodi, F.A.; Screen, S.E.; Anil, V.S.; Zhu, S.; Morrell, J.A.; Liu, G. Expression of the Arabidopsis thaliana BBX32 gene in soybean increases grain yield. PLoS ONE 2012, 7, e30717. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.; Köster, T. On the move through time–a historical review of plant clock research. Plant Biol. 2019, 21, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Paltiel, J.; Amin, R.; Gover, A.; Ori, N.; Samach, A. Novel roles for GIGANTEA revealed under environmental conditions that modify its expression in Arabidopsis and Medicago truncatula. Planta 2006, 224, 1255–1268. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Han, L.; Liu, X.; Wang, H.; Wen, L.; Yu, X.; Xu, X.; Kong, F.; Fu, C.; Mysore, K.S. The nodulation and nyctinastic leaf movement is orchestrated by clock gene LHY in Medicago truncatula. J. Integr. Plant Biol. 2020, 62, 1880–1895. [Google Scholar] [CrossRef]
- Ishida, K.; Niwa, Y.; Yamashino, T.; Mizuno, T. A genome-wide compilation of the two-component systems in Lotus japonicus. DNA Res. 2009, 16, 237–247. [Google Scholar] [CrossRef]
- Ueoka-Nakanishi, H.; Yamashino, T.; Ishida, K.; Kamioka, M.; Nakamichi, N.; Mizuno, T. Molecular mechanisms of circadian rhythm in Lotus japonicus and Arabidopsis thaliana are sufficiently compatible to regulate heterologous core clock genes robustly. Biosci. Biotechnol. Biochem. 2012. [Google Scholar] [CrossRef] [Green Version]
- Wakabayashi, T.; Oh, H.; Kawaguchi, M.; Harada, K.; Sato, S.; Ikeda, H.; Hiroaki, S. Polymorphisms of E1 and GIGANTEA in wild populations of Lotus japonicus. J. Plant Res. 2014, 127, 651–660. [Google Scholar] [CrossRef]
- Liu, H.; Wang, H.; Gao, P.; Xü, J.; Xü, T.; Wang, J.; Wang, B.; Lin, C.; Fu, Y.-F. Analysis of clock gene homologs using unifoliolates as target organs in soybean (Glycine max). J. Plant Physiol. 2009, 166, 278–289. [Google Scholar] [CrossRef]
- Marcolino-Gomes, J.; Nakayama, T.J.; Molinari, H.B.C.; Basso, M.F.; Henning, L.M.M.; Fuganti-Pagliarini, R.; Harmon, F.G.; Nepomuceno, A.L. Functional characterization of a putative Glycine max ELF4 in transgenic Arabidopsis and its role during flowering control. Front. Plant Sci. 2017, 8, 618. [Google Scholar] [CrossRef]
- Watanabe, S.; Xia, Z.; Hideshima, R.; Tsubokura, Y.; Sato, S.; Yamanaka, N.; Takahashi, R.; Anai, T.; Tabata, S.; Kitamura, K. A map-based cloning strategy employing a residual heterozygous line reveals that the GIGANTEA gene is involved in soybean maturity and flowering. Genetics 2011, 188, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Q.; Dong, L.; Su, T.; Li, T.; Gan, Z.; Nan, H.; Lu, S.; Fang, C.; Kong, L.; Li, H. CRISPR/Cas9-mediated targeted mutagenesis of GmLHY genes alters plant height and internode length in soybean. BMC Plant Biol. 2019, 19, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, L.C.; Singh, M.B.; Bhalla, P.L. A novel role of the soybean clock gene LUX ARRHYTHMO in male reproductive development. Sci. Rep. 2017, 7, 10605. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.-G.; Zhang, X.-M.; Lei, C.-F.; Chen, X.-J.; Fu, Y.-F. Molecular cloning and functional analysis of one ZEITLUPE homolog GmZTL3 in soybean. Mol. Biol. Rep. 2012, 39, 1411–1418. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sun, S.; Wu, T.; Liu, L.; Sun, X.; Cai, Y.; Li, J.; Jia, H.; Yuan, S.; Chen, L. Natural variation and CRISPR/Cas9-mediated mutation in GmPRR37 affect photoperiodic flowering and contribute to regional adaptation of soybean. Plant Biotechnol. J. 2020, 18, 1869–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, L.; Hecht, V.; Weeden, N.; Weller, J. Isolation of pseudo response regulator genes and evaluation as candidate genes for photoperiod response loci. Pisum. Genet. 2009, 41, 21–25. [Google Scholar]
- Liew, L.C.; Hecht, V.; Laurie, R.E.; Knowles, C.L.; Vander Schoor, J.K.; Macknight, R.C.; Weller, J.L. DIE NEUTRALIS and LATE BLOOMER 1 contribute to regulation of the pea circadian clock. Plant Cell 2009, 21, 3198–3211. [Google Scholar] [CrossRef] [Green Version]
- Weller, J.L.; Liew, L.C.; Hecht, V.F.; Rajandran, V.; Laurie, R.E.; Ridge, S.; Wenden, B.; Vander Schoor, J.K.; Jaminon, O.; Blassiau, C. A conserved molecular basis for photoperiod adaptation in two temperate legumes. Proc. Natl. Acad. Sci. USA 2012, 109, 21158–21163. [Google Scholar] [CrossRef] [Green Version]
- Liew, L.C.; Hecht, V.; Sussmilch, F.C.; Weller, J.L. The pea photoperiod response gene STERILE NODES is an ortholog of LUX ARRHYTHMO. Plant. Physiol. 2014, 165, 648–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridge, S.; Deokar, A.; Lee, R.; Daba, K.; Macknight, R.C.; Weller, J.L.; Tar’an, B. The chickpea Early Flowering 1 (Efl1) locus is an ortholog of Arabidopsis ELF3. Plant Physiol. 2017, 175, 802–815. [Google Scholar] [CrossRef] [Green Version]
- Rychel, S.; Książkiewicz, M.; Tomaszewska, M.; Bielski, W.; Wolko, B. FLOWERING LOCUS T, GIGANTEA, SEPALLATA, and FRIGIDA homologs are candidate genes involved in white lupin (Lupinus albus L.) early flowering. Mol. Breed. 2019, 39, 43. [Google Scholar] [CrossRef] [Green Version]
- Kaldis, A.-D.; Kousidis, P.; Kesanopoulos, K.; Prombona, A. Light and circadian regulation in the expression of LHY and Lhcb genes in Phaseolus vulgaris. Plant Mol. Biol. 2003, 52, 981–997. [Google Scholar] [CrossRef]
- Kaldis, A.-D.; Prombona, A. Synergy between the light-induced acute response and the circadian cycle: A new mechanism for the synchronization of the Phaseolus vulgaris clock to light. Plant Mol. Biol. 2006, 61, 883–895. [Google Scholar] [CrossRef]
- Galeou, A.; Prombona, A. Light at night resynchronizes the evening-phased rhythms of TOC1 and ELF4 in Phaseolus vulgaris. Plant Sci. 2012, 184, 141–147. [Google Scholar] [CrossRef]
- Galeou, A.; Roussis, A.; Prombona, A. Investigation of the Phaseolus vulgaris circadian clock and the repressive role of the PvTOC1 factor by a newly established in vitro system. J. Plant Physiol. 2018, 222, 79–85. [Google Scholar] [CrossRef]
- Kwak, M.; Velasco, D.; Gepts, P. Mapping homologous sequences for determinacy and photoperiod sensitivity in common bean (Phaseolus vulgaris). J. Hered. 2008, 99, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.; Terry, M.I.; Martos-Fuentes, M.; Letourneux, L.; Ruiz-Hernández, V.; Fernández, J.A.; Egea-Cortines, M. Diel pattern of circadian clock and storage protein gene expression in leaves and during seed filling in cowpea (Vigna unguiculata). BMC Plant Biol. 2018, 18, 33. [Google Scholar] [CrossRef] [Green Version]
- Bünning, E. Über die Erblichkeit der Tagesperiodizität bei den Phaseolus Blättern. Jahrb Bot. 1932, 81, 411–418. [Google Scholar]
- Bünning, E.; Moser, I. Light-induced phase shifts of circadian leaf movements of Phaseolus: Comparison with the effects of potassium and of ethyl alcohol. Proc. Natl. Acad. Sci. USA 1973, 70, 3387–3389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloppstech, K. Diurnal and circadian rhythmicity in the expression of light-induced plant nuclear messenger RNAs. Planta 1985, 165, 502–506. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.G.; Klein, W.H. Photocontrol of dark circadian rhythms in stomata of Phaseolus vulgaris L. Plant Physiol. 1986, 82, 28–33. [Google Scholar] [CrossRef] [Green Version]
- Hennessey, T.L.; Field, C.B. Evidence of multiple circadian oscillators in bean plants. J. Biol. Rhythms 1992, 7, 105–113. [Google Scholar] [CrossRef]
- Hennessey, T.L.; Freeden, A.L.; Field, C.B. Environmental effects on circadian rhythms in photosynthesis and stomatal opening. Planta 1993, 189, 369–376. [Google Scholar] [CrossRef]
- Weller, J.L.; Hecht, V.; Liew, L.C.; Sussmilch, F.C.; Wenden, B.; Knowles, C.L.; Vander Schoor, J.K. Update on the genetic control of flowering in garden pea. J. Exp. Bot. 2009, 60, 2493–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, N.; Ishida, K.; Yamashino, T.; Nakanishi, H.; Sato, S.; Tabata, S.; Mizuno, T. Genomewide characterization of the light-responsive and clock-controlled output pathways in Lotus japonicus with special emphasis of its uniqueness. Plant Cell Physiol. 2010, 51, 1800–1814. [Google Scholar] [CrossRef] [Green Version]
- Marcolino-Gomes, J.; Rodrigues, F.A.; Fuganti-Pagliarini, R.; Bendix, C.; Nakayama, T.J.; Celaya, B.; Molinari, H.B.C.; de Oliveira, M.C.N.; Harmon, F.G.; Nepomuceno, A. Diurnal oscillations of soybean circadian clock and drought responsive genes. PLoS ONE 2014, 9, e86402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.-W.; Lam, H.-M. The modification of circadian clock components in soybean during domestication and improvement. Front. Genet. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Badhan, S.; Ball, A.S.; Mantri, N. First report of CRISPR/Cas9 mediated DNA-free editing of 4CL and RVE7 genes in chickpea protoplasts. Int. J. Mol. Sci. 2021, 22, 396. [Google Scholar] [CrossRef] [PubMed]
- Young, N.D.; Debellé, F.; Oldroyd, G.E.; Geurts, R.; Cannon, S.B.; Udvardi, M.K.; Benedito, V.A.; Mayer, K.F.; Gouzy, J.; Schoof, H. The Medicago genome provides insight into the evolution of rhizobial symbioses. Nature 2011, 480, 520–524. [Google Scholar] [CrossRef] [Green Version]
- Crane, C.; Dixon, R.A.; Wang, Z.-Y. Medicago truncatula transformation using root explants. Methods Mol. Biol. 2006, 343, 137–142. [Google Scholar]
- Cerri, M.R.; Frances, L.; Kelner, A.; Fournier, J.; Middleton, P.H.; Auriac, M.-C.; Mysore, K.S.; Wen, J.; Erard, M.; Barker, D.G. The symbiosis-related ERN transcription factors act in concert to coordinate rhizobial host root infection. Plant Physiol. 2016, 171, 1037–1054. [Google Scholar] [CrossRef]
- Schiessl, K.; Lilley, J.L.S.; Lee, T.; Tamvakis, I.; Kohlen, W.; Bailey, P.C.; Thomas, A.; Luptak, J.; Ramakrishnan, K.; Carpenter, M.D.; et al. NODULE INCEPTION recruits the lateral root developmental program for symbiotic nodule organogenesis in Medicago truncatula. Curr. Biol. 2019, 29, 3657–3668. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Yi, D.; Yang, J.; Liu, X.; Pang, Y. Genome-wide identification, expression analysis and functional study of CCT Gene family in Medicago truncatula. Plants 2020, 9, 513. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, M. CCT family genes in cereal crops: A current overview. Crop J. 2017, 5, 449–458. [Google Scholar] [CrossRef]
- Duangkhet, M.; Thepsukhon, A.; Widyastuti, R.; Santosa, D.A.; Tajima, S.; Nomura, M. A MYB-related transcription factor affects nodule formation in Lotus japonicus. Plant Biotechnol. 2016, 33, 187–194. [Google Scholar]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Fujiwara, S.; Oda, A.; Yoshida, R.; Niinuma, K.; Miyata, K.; Tomozoe, Y.; Tajima, T.; Nakagawa, M.; Hayashi, K.; Coupland, G. Circadian clock proteins LHY and CCA1 regulate SVP protein accumulation to control flowering in Arabidopsis. Plant Cell 2008, 20, 2960–2971. [Google Scholar] [CrossRef] [Green Version]
- Boycheva, I.; Vassileva, V.; Revalska, M.; Zehirov, G.; Iantcheva, A. Cyclin-like F-box protein plays a role in growth and development of the three model species Medicago truncatula, Lotus japonicus, and Arabidopsis thaliana. Res. Rep. Biol. 2015, 6, 117–130. [Google Scholar]
- Shah, N.; Wakabayashi, T.; Kawamura, Y.; Skovbjerg, C.K.; Wang, M.-Z.; Mustamin, Y.; Isomura, Y.; Gupta, V.; Jin, H.; Mun, T. Extreme genetic signatures of local adaptation during Lotus japonicus colonization of Japan. Nat. Commun. 2020, 11, 253. [Google Scholar] [CrossRef] [Green Version]
- Homrich, M.S.; Wiebke-Strohm, B.; Weber, R.L.M.; Bodanese-Zanettini, M.H. Soybean genetic transformation: A valuable tool for the functional study of genes and the production of agronomically improved plants. Genet. Mol. Biol. 2012, 35 (Suppl. 4), 998–1010. [Google Scholar] [CrossRef] [Green Version]
- Christou, P.; McCabe, D.E.; Martinell, B.J.; Swain, W.F. Soybean genetic engineering-commercial production of transgenic plants. Trends Biotechnol. 1990, 8, 145–151. [Google Scholar] [CrossRef]
- Bahrman, N.; Hascoët, E.; Jaminon, O.; Dépta, F.; Hû, J.-F.; Bouchez, O.; Lejeune-Hénaut, I.; Delbreil, B.; Legrand, S. Identification of genes differentially expressed in response to cold in Pisum sativum using RNA sequencing analyses. Plants 2019, 8, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohapatra, C.; Chand, R.; Tiwari, J.K.; Singh, A.K. Effect of heat stress during flowering and pod formation in pea (Pisum sativum L.). Physiol. Mol. Biol. Plants 2020, 26, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- García-García, M.C.; Celestino, M.R.; Gil-Izquierdo, A.; Egea-Gilabert, C.; Galano, J.M.; Durand, T.; Oger, C.; Fernández, J.A.; Ferreres, F.; Domínguez-Perles. The value of legume foods as a dietary source of phytoprostanes and phytofurans is dependent on species, variety, and growing conditions. Eur. J. Lipid Sci. Technol. 2019, 121. [Google Scholar] [CrossRef]
- Kerr, S.C.; Gaiti, F.; Beveridge, C.A.; Tanurdzic, M. De novo transcriptome assembly reveals high transcriptional complexity in Pisum sativum axillary buds and shows rapid changes in expression of diurnally regulated genes. BMC Genom. 2017, 18, 221. [Google Scholar] [CrossRef] [Green Version]
- Creux, N.; Harmer, S. Circadian rhythms in plants. Cold Spring Harb. Perspect. Biol. 2019, 11. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene | Arabidopsis Homologue | Function(s) | Reference(s) |
|---|---|---|---|---|
| Barrel medic (Medicago truncatula) | MtGI | GI | Circadian clock component, photoperiod response | [47] |
| MtLHY | LHY | Regulation of circadian rhythm in nodules and nyctinastic leaf movement | [48] | |
| Birds-foot trefoil (Lotus japonicus) | LjaPRR5 | PRR5 | Component of the circadian rhythm | [49] |
| LjaPRR7 | PRR7 | |||
| LjaPRR9 | PRR9 | |||
| LjaLUX | LUX | |||
| LjaTOC1 | TOC1 | Central component of the circadian rhythm | ||
| LjaLHY | LHY | |||
| LjCCA1 | CCA1 | Central component of the circadian rhythm | [50] | |
| LjGI | GI | Possible regulation of flowering time | [51] | |
| Soybean (Glycine max) | GmTOC1 | TOC1 | Central component of the soybean circadian clock (expressed as an evening gene) | [52] |
| GmELF4 | ELF4 | Circadian clock function. | [53] | |
| GmGIa | GI | Photoperiod response, flowering time regulation | [54] | |
| GmLCL1 | LHY/CCA1 | Central component of the soybean circadian clock (expressed as a morning gene) | [52] | |
| GmLCL2 | ||||
| GmLHY | LHY | Regulate plant height | [55] | |
| GmLUXa | LUX | Control flowering time | [56] | |
| GmLUXb | ||||
| GmLUXc | ||||
| GmZTL3 | ZTL | Control of flowering time (inhibitor of flowering induction) and photoreceptor. | [57] | |
| GmPRR37 | PRR3 & 7 | Control of soybean photoperiodic flowering | [58] | |
| Common pea (Pisum sativum) | PsTOC1 | TOC1 | Circadian clock component | [17,59] |
| DNE | ELF4 | Circadian clock component, flowering time regulation | [60,61] | |
| HR | ELF3 | Circadian clock component, flowering time regulation, light response | [61] | |
| LATE1 | GI | Photoperiod response | [22,60] | |
| MYB1/LHY | CCA1/LHY | Circadian clock component | [60] | |
| SN | LUX | Circadian clock component | [62] | |
| PsPRR37 | PRR | Component of phospho-relay signal transduction system | [59] | |
| PsPRR59 | ||||
| Chickpea (Cicer arietinum) | Efl1 | ELF3 | Flowering regulation (light input to the circadian clock) | [63] |
| GI | GI | Flowering time regulation | [64] | |
| Common bean (Phaseolus vulgaris) | PvLHY | LHY | Circadian mechanism regulation | [65,66] |
| PvTOC1 | TOC1 | Mediating light responsiveness to circadian clock mechanism | [67,68] | |
| PvELF4 | ELF4 | Evening-expressed gene | [67] | |
| PvGI | GI | Circadian clock component | [69] | |
| PvZTL | ZTL | |||
| Pigeon pea (Cajanus cajan) | CcGI | GI | Determinacy and flower patterning | [69] |
| Cowpea (Vigna unguiculata) | VunTOC1 | TOC1 | Circadian clock function in seed filling and leaves | [70] |
| VunLHY | LHY | |||
| VunELF3 | ELF3 | |||
| VunGI | GI | |||
| Lentil (Lens culinaris) | HR | ELF3 | Flowering time regulation | [61] |
| White lupin (Lupinis albus) | GI | GI | Flowering regulation; anthracnose resistance | [64] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kugan, H.M.; Rejab, N.A.; Sahruzaini, N.A.; Harikrishna, J.A.; Baisakh, N.; Cheng, A. Circadian Rhythms in Legumes: What Do We Know and What Else Should We Explore? Int. J. Mol. Sci. 2021, 22, 4588. https://doi.org/10.3390/ijms22094588
Kugan HM, Rejab NA, Sahruzaini NA, Harikrishna JA, Baisakh N, Cheng A. Circadian Rhythms in Legumes: What Do We Know and What Else Should We Explore? International Journal of Molecular Sciences. 2021; 22(9):4588. https://doi.org/10.3390/ijms22094588
Chicago/Turabian StyleKugan, Hazel Marie, Nur Ardiyana Rejab, Nurul Amylia Sahruzaini, Jennifer Ann Harikrishna, Niranjan Baisakh, and Acga Cheng. 2021. "Circadian Rhythms in Legumes: What Do We Know and What Else Should We Explore?" International Journal of Molecular Sciences 22, no. 9: 4588. https://doi.org/10.3390/ijms22094588
APA StyleKugan, H. M., Rejab, N. A., Sahruzaini, N. A., Harikrishna, J. A., Baisakh, N., & Cheng, A. (2021). Circadian Rhythms in Legumes: What Do We Know and What Else Should We Explore? International Journal of Molecular Sciences, 22(9), 4588. https://doi.org/10.3390/ijms22094588