
Cardiolipin Stabilizes and Increases Catalytic Efficiency of Carnitine Palmitoyltransferase II and Its Variants S113L, P50H, and Y479F

 and
and
Abstract
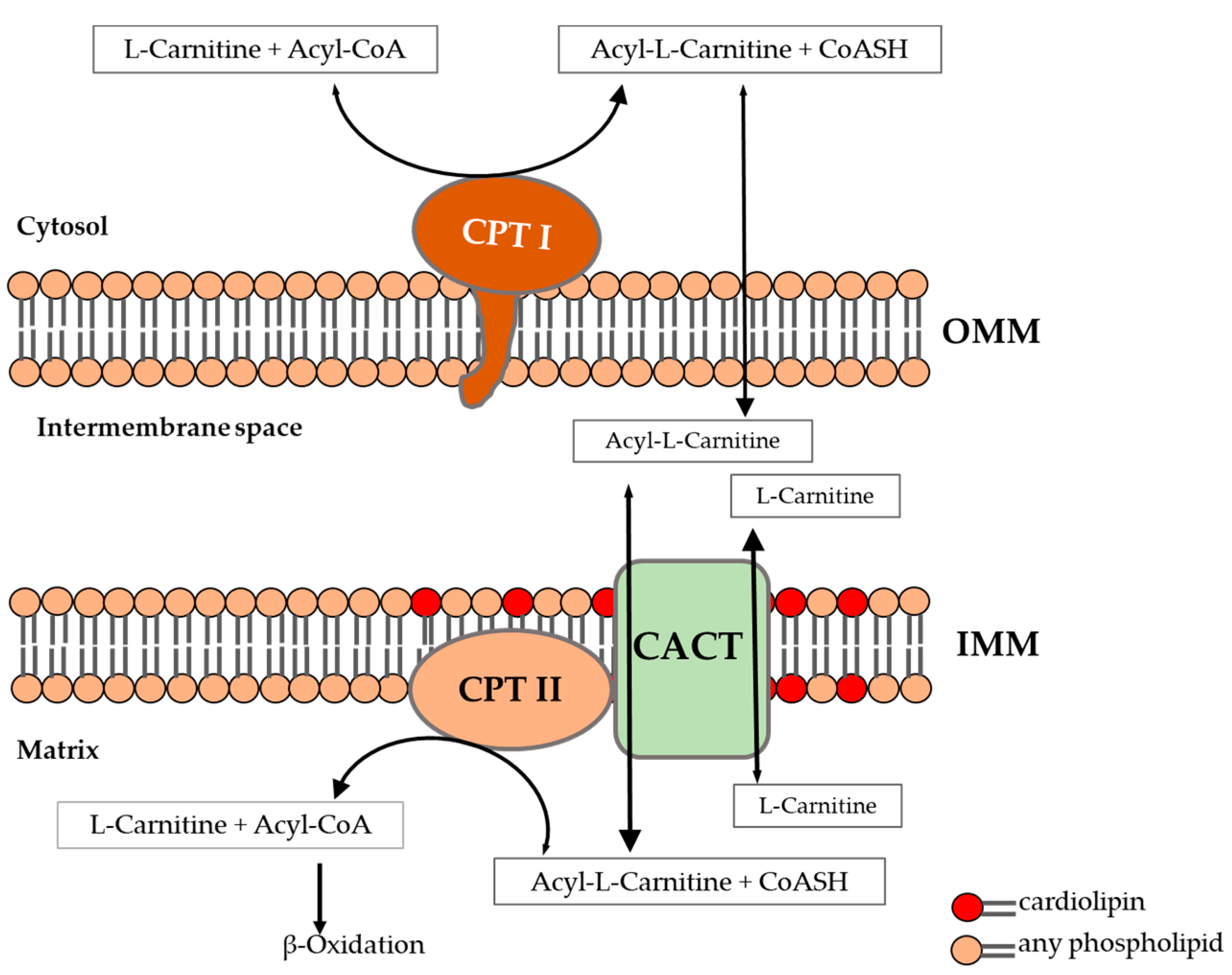
:1. Introduction
2. Results
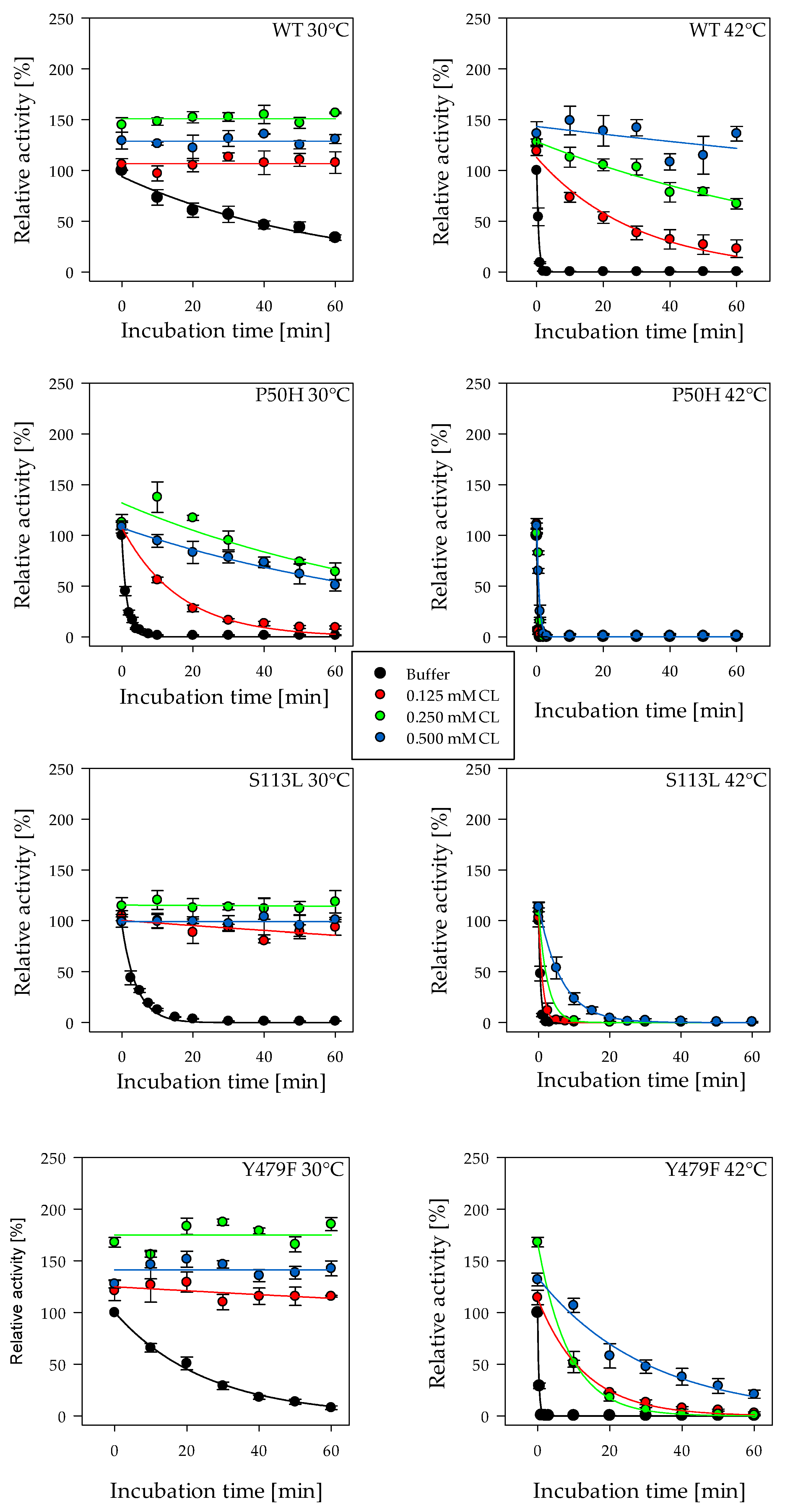
2.1. Thermosensitivity of CPT II Variants
2.2. Effect of Cardiolipin on Thermosensitivity of the Enzyme Activity
2.3. Effect of Cardiolipin on the Protein Stability
2.4. Effect of CL on the Catalytic Efficiency of CPT II
2.4.1. Conversion of Palmitoyl-CoA
2.4.2. Conversion of L-carnitine
2.5. Effect of CL on the Inhibition by R-amino Carnitine
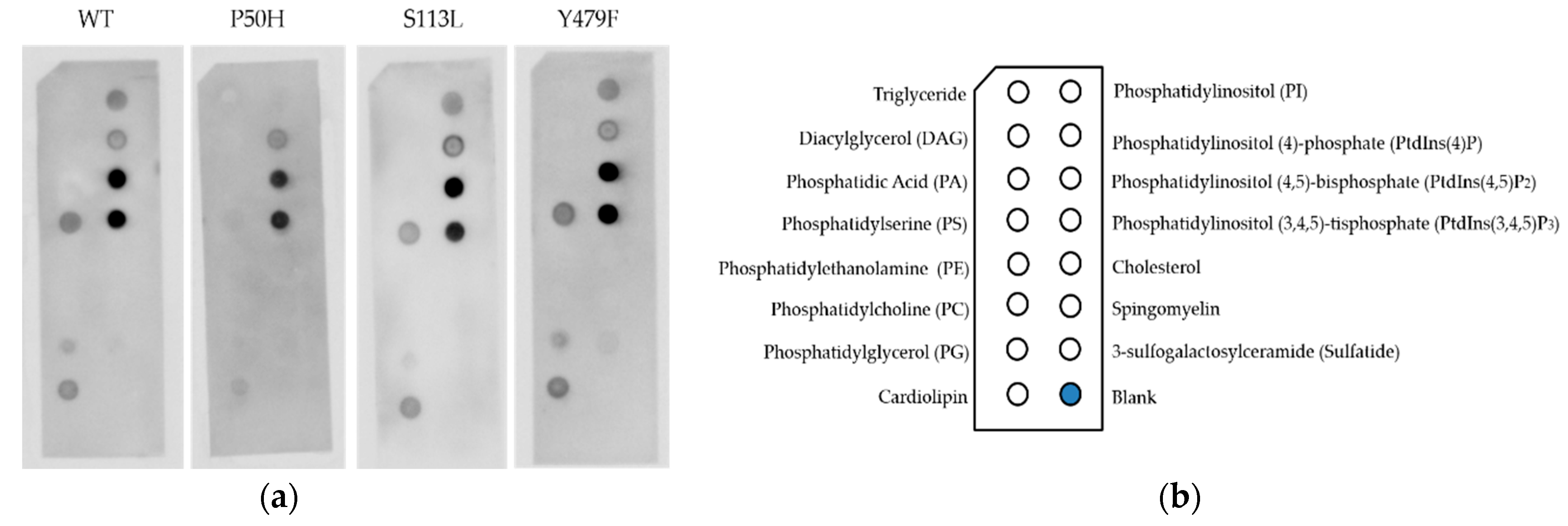
2.6. Lipid Binding Assay
3. Discussion
4. Materials and Methods
4.1. Generation of CPT II Variants S113L, P50H, Y479F
4.2. Recombinant Expression and Purification of CPT II WT and Variants
4.3. Enzyme Activity Assay
4.4. Influence of Temperature and CL on the Enzyme Activity
4.5. Assessment of Protein Stability
4.6. Determination of the Kinetic Parameters
4.7. Determination of Inhibition Constant of for R-amino Carnitine for CPT II
4.8. Lipid Binding Assay
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CPT | Carnitine palmitoyltransferase |
| CACT | Carnitine:acylcarnitine translocase |
| LCFA | Long chain fatty acid |
| CL | Cardiolipin |
| Pal-CoA | Palmitoyl-Coenzyme A |
| CoA | Coenzyme A |
| WT | Wild type |
| PBS | Phosphate buffered saline |
| R-AC | R-Aminocarnitine |
| IMM | Inner mitochondrial membrane |
| β-OG | n-octyl-β-d-glucopyranoside |
| nanoDSF | Nano differential scanning fluorimetry |
References
- Houten, S.M.; Wanders, R.J. A General Introduction to the Biochemistry of Mitochondrial Fatty Acid Beta-Oxidation. J. Inherit. Metab. Dis. 2010, 33, 469–477. [Google Scholar] [CrossRef] [Green Version]
- McGarry, J.D.; Brown, N.F. The Mitochondrial Carnitine Palmitoyltransferase System. From Concept to Molecular Analysis. Eur. J. Biochem. 1997, 244, 1–14. [Google Scholar] [CrossRef]
- McGarry, J. Travels with Carnitine Palmitoyltransferase I: From Liver to Germ Cell with Stops in Between; Portland Press Ltd.: London, UK, 2001. [Google Scholar]
- Finocchiaro, G.; Colombo, I.; DiDonato, S. Purification, Characterization and Partial Amino Acid Sequences of Carnitine Palmitoyl-Transferase from Human Liver. FEBS Lett. 1990, 274, 163–166. [Google Scholar]
- Roe, C.; Coates, P. Disorders of Mitochondrial Function. In The Metabolic and Molecular Ba Inherited Disease; Scriver, C., Beaudet, A.L., Sly, W.S., Valle, D., Eds.; McGraw-Hill: New York, NY, USA, 1995; pp. 1501–1533. [Google Scholar]
- Console, L.; Giangregorio, N.; Indiveri, C.; Tonazzi, A. Carnitine/Acylcarnitine Translocase and Carnitine Palmitoyltransferase 2 form a Complex in the Inner Mitochondrial Membrane. Mol. Cell. Biochem. 2014, 394, 307–314. [Google Scholar] [CrossRef]
- Rufer, A.C.; Thoma, R.; Hennig, M. Structural Insight into Function and Regulation of Carnitine Palmitoyltransferase. Cell Mol. Life Sci. 2009, 66, 2489–2501. [Google Scholar] [CrossRef]
- Osman, C.; Voelker, D.R.; Langer, T. Making Heads or Tails of Phospholipids in Mitochondria. J. Cell Biol. 2011, 192, 7–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradies, G.; Paradies, V.; De Benedictis, V.; Ruggiero, F.M.; Petrosillo, G. Functional Role of Cardiolipin in Mitochondrial Bioenergetics. Biochim. Biophys. Acta 2014, 1837, 408–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikon, N.; Ryan, R.O. Cardiolipin and Mitochondrial Cristae Organization. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1156–1163. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Bayir, H.A.; Belikova, N.A.; Kapralov, O.; Tyurina, Y.Y.; Tyurin, V.A.; Jiang, J.; Stoyanovsky, D.A.; Wipf, P.; Kochanek, P.M.; et al. Cytochrome C/Cardiolipin Relations in Mitochondria: A Kiss of Death. Free Radic. Biol. Med. 2009, 46, 1439–1453. [Google Scholar] [CrossRef] [Green Version]
- Schlame, M.; Rua, D.; Greenberg, M.L. The biosynthesis and functional role of cardiolipin. Prog. Lipid Res. 2000, 39, 257–288. [Google Scholar] [CrossRef]
- Noël, H.; Pande, S.V. An Essential Requirement of Cardiolipin for Mitochondrial Carnitine Acylcarnitine Translocase Activity: Lipid Requirement of Carnitine Acylcarnitine Translocase. Eur. J. Biochem. 1986, 155, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Gozalbo, M.; Bakker, J.; Waterham, H.; Wanders, R. Carnitine–Acylcarnitine Translocase Deficiency, Clinical, Biochemical and Genetic Aspects. Mol. Asp. Med. 2004, 25, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Palsdottir, H.; Hunte, C. Lipids in Membrane Protein Structures. Biochim. Biophys. Acta 2004, 1666, 2–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rufer, A.C.; Lomize, A.; Benz, J.; Chomienne, O.; Thoma, R.; Hennig, M. Carnitine Palmitoyltransferase 2: Analysis of Membrane Association and Complex Structure with a Substrate Analog. FEBS Lett. 2007, 581, 3247–3252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashfi, K.; Mynatt, R.L.; Park, E.A.; Cook, G.A. Membrane Microenvironment Regulation of Carnitine Palmitoyltranferases I and II. Biochem. Soc. Trans. 2011, 39, 833–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isackson, P.J.; Bennett, M.J.; Lichter-Konecki, U.; Willis, M.; Nyhan, W.L.; Sutton, V.R.; Tein, I.; Vladutiu, G.D. CPT2 Gene Mutations Resulting in Lethal Neonatal or Severe Infantile Carnitine Palmitoyltransferase II Deficiency. Mol. Genet. Metab. 2008, 94, 422–427. [Google Scholar] [CrossRef]
- DiMauro, S.; DiMauro, P.M.M. Muscle Carnitine Palmityltransferase Deficiency and Myoglobinuria. Science 1973, 182, 929–931. [Google Scholar] [CrossRef]
- Bonnefont, J.P.; Djouadi, F.; Prip-Buus, C.; Gobin, S.; Munnich, A.; Bastin, J. Carnitine Palmitoyltransferases 1 and 2: Biochemical, Molecular and Medical Aspects. Mol. Asp. Med. 2004, 25, 495–520. [Google Scholar] [CrossRef] [PubMed]
- Zierz, S. Carnitine Palmitoyltransferase Deficiency. Myopathy 1994, 124, 851–854. [Google Scholar]
- Deschauer, M.; Wieser, T.; Zierz, S. Muscle Carnitine Palmitoyltransferase II Deficiency: Clinical and Molecular Genetic Features and Diagnostic Aspects. Arch. Neurol. 2005, 62, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Joshi, P.R.; Deschauer, M.; Zierz, S. Carnitine Palmitoyltransferase II (CPT II) Deficiency: Genotype-Phenotype Analysis of 50 Patients. J. Neurol. Sci. 2014, 338, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M. Die Regulation der Körpertemperatur bei Muskelarbeit 1. Skand. Arch. Für Physiol. 1938, 79, 193–230. [Google Scholar] [CrossRef]
- PELLEGRINI, A.; RIVA, G.; Margaria, R. La Termoregolazione Nel Lavoro Muscolare. Arch. Di. Fisiol. 1947, 46, 111–133. [Google Scholar]
- Jameson, J.L. Harrison’s Principles of Internal Medicine; McGraw-Hill Education: New York, NY, USA, 2018. [Google Scholar]
- Kluger, M.J. Fever: Its Biology, Evolution, and Function; Princeton University Press: Princeton, NJ, USA, 2015. [Google Scholar]
- Chung, N.; Park, J.; Lim, K. The Effects of Exercise and Cold Exposure on Mitochondrial Biogenesis in Skeletal Muscle and White Adipose Tissue. J. Exerc. Nutr. Biochem. 2017, 21, 39. [Google Scholar] [CrossRef] [PubMed]
- Motlagh, L.; Golbik, R.; Sippl, W.; Zierz, S. Stabilization of the Thermolabile Variant S113L of Carnitine Palmitoyltransferase II. Neurol. Genet. 2016, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rufer, A.C.; Thoma, R.; Benz, J.; Stihle, M.; Gsell, B.; De Roo, E.; Banner, D.W.; Mueller, F.; Chomienne, O.; Hennig, M. The Crystal Structure of Carnitine Palmitoyltransferase 2 and Implications for Diabetes Treatment. Structure 2006, 14, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Isackson, P.J.; Bennett, M.J.; Vladutiu, G.D. Identification of 16 New Disease-Causing Mutations in the CPT2 Gene Resulting in Carnitine Palmitoyltransferase II deficiency. Mol. Genet. Metab. 2006, 89, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Motlagh, L.; Golbik, R.; Sippl, W.; Zierz, S. Malony-CoA Inhibits the S113L Variant of Carnitine-Palmitoyltransferase II. Biochim. Biophys. Acta 2016, 1861, 34–40. [Google Scholar] [CrossRef]
- Verderio, E.; Cavadini, P.; Montermini, L.; Wang, H.; Lamantea, E.; Finocchiaro, G.; DiDonato, S.; Gellera, C.; Taroni, F. Carnitine Palmitoyltransferase II Deficiency: Structure of the Gene and Characterization of Two Novel Disease-Causing Mutations. Hum. Mol. Genet. 1995, 4, 19–29. [Google Scholar] [CrossRef]
- Wieser, T.; Deschauer, M.; Olek, K.; Hermann, T.; Zierz, S. Carnitine Palmitoyltransferase II Deficiency: Molecular and Biochemical Analysis of 32 Patients. Neurology 2003, 60, 1351–1353. [Google Scholar] [CrossRef]
- Hsiao, Y.-S.; Jogl, G.; Esser, V.; Tong, L. Crystal Structure of Rat Carnitine Palmitoyltransferase II (CPT-II). Biochem. Biophys. Res. Commun. 2006, 346, 974–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, J.; Lord, H.; Knutson, N.; Wikstrom, M. Nano Differential Scanning Fluorimetry for Comparability Studies of Therapeutic Proteins. Anal. Biochem. 2020, 593, 113581. [Google Scholar] [CrossRef] [PubMed]
- Gihaz, S.; Weiser, D.; Dror, A.; Satorhelyi, P.; Jerabek-Willemsen, M.; Poppe, L.; Fishman, A. Creating an Efficient Methanol-Stable Biocatalyst by Protein and Immobilization Engineering Steps towards Efficient Biosynthesis of Biodiesel. ChemSusChem 2016, 9, 3161–3170. [Google Scholar] [CrossRef]
- Taroni, F.; Verderio, E.; Dworzak, F.; Willems, P.J.; Cavadini, P.; DiDonato, S. Identification of a common mutation in the carnitine palmitoyltransferase II gene in familial recurrent myoglobinuria patients. Nat. Genet. 1993, 4, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Yasuno, T.; Kaneoka, H.; Tokuyasu, T.; Aoki, J.; Yoshida, S.; Takayanagi, M.; Ohtake, A.; Kanazawa, M.; Ogawa, A.; Tojo, K.; et al. Mutations of Carnitine Palmitoyltransferase II (CPT II) in Japanese Patients with CPT II Deficiency. Clin. Genet. 2008, 73, 496–501. [Google Scholar] [CrossRef]
- Yao, D.; Mizuguchi, H.; Yamaguchi, M.; Yamada, H.; Chida, J.; Shikata, K.; Kido, H. Thermal Instability of Compound Variants of Carnitine Palmitoyltransferase II and Impaired Mitochondrial Fuel Utilization in Influenza-Associated Encephalopathy. Hum. Mutat. 2008, 29, 718–727. [Google Scholar] [CrossRef]
- Motlagh Scholle, L.; Thaele, A.; Beckers, M.; Meinhardt, B.; Zierz, S. Lack of Activation of the S113L Variant of Carnitine Palmitoyltransfersase II by Cardiolipin. J. Bioenerg. Biomembr. 2018, 50, 461–466. [Google Scholar] [CrossRef]
- Ellman, G.L. A Colorimetric Method for Determining Low Concentrations of Mercaptans. Arch. Biochem. Biophys. 1958, 74, 443–450. [Google Scholar] [CrossRef]
- Nic a′ Bháird, N.; Kumaravel, G.; Gandour, R.; Krueger, M.; Ramsay, R. Comparison of the Active Sites of the Purified Carnitine Acyltransferases from Peroxisomes and Mitochondria by Using a Reaction-Intermediate Analogue. Biochem. J. 1993, 294, 645–651. [Google Scholar] [CrossRef] [Green Version]
- Brown, N.F.; Anderson, R.C.; Caplan, S.L.; Foster, D.W.; McGarry, J.D. Catalytically Important Domains of Rat Carnitine Palmitoyltransferase II as Determined by Site-Directed Mutagenesis and Chemical Modification. Evidence for a Critical Histidine Residue. J. Biol. Chem. 1994, 269, 19157–19162. [Google Scholar] [CrossRef]
- Copeland, R.A. Evaluation of Enzyme Inhibitors in Drug Discovery. Wiley-Interscience 2005, 34–37. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CPT II Variant | In Absence of CL | In Presence of CL | ||||
|---|---|---|---|---|---|---|
| kcat (s−1) | KM (µM) | kcat/KM (µM1 s−1) | kcat (s−1) | KM (µM) | kcat/KM (µM1 s−1) | |
| WT | 144.7 ± 3.1 | 7.10 ± 1.1 | 20.5 ± 3.9 | 145.3 ± 9.0 | 6.7 ± 1.1 | 21.8 ± 2.3 |
| P50H | 144.3 ± 18 | 5.50 ± 1.4 | 26.4 ± 7.5 | 138.2 ± 8.4 | 6.7 ±1.8 | 20.7 ± 3.8 |
| S113L | 142.1 ± 25 | 11.0 ± 3.6 | 12.9 ± 1.8 * | 145.8 ± 0.9 | 12.3 ± 3.0 | 11.9 ± 2.9 * |
| Y479F | 141.4 ± 1.7 | 5.10 ± 2.3 | 27.5 ± 4.4 | 144.8 ± 13 | 6.7 ± 0.3 | 21.7 ± 4.7 |
| CPT II Variant | In Absence of CL | In Presence of CL | ||||
|---|---|---|---|---|---|---|
| kcat (s−1) | KM (mM) | kcat/KM (mM1 s−1) | kcat (s−1) | KM (mM) | kcat/KM (mM1 s−1) | |
| WT | 148.6 ± 11.1 | 15.1 ± 3.4 | 9.8 ± 1.6 | 129.9 ± 8.1 | 5.5 ± 1.0 | 23.6 ± 3.1 * |
| P50H | 137.7 ± 4.9 | 12.8 ± 2.7 | 10.8 ± 2.6 | 138.1 ± 5.6 | 6.6 ±2.8 | 21.0 ± 1.8 * |
| S113L | 144.0 ± 5.0 | 15.5 ± 2.3 | 9.3 ± 1.4 | 135.2 ± 2.5 | 6.5 ± 0.2 | 20.7 ± 0.3 * |
| Y479F | 140.5 ± 18.5 | 17.4 ± 4.4 | 8.1 ± 1.1 | 138.7 ± 1.4 | 4.5 ± 0.5 | 30.9 ± 2.6 * |
| CPT II Variant | Inhibition Constant Ki (mM) in Absence of CL | Inhibition Constant Ki (mM) in Presence of CL |
|---|---|---|
| WT | 0.493 ± 0.004 | 0.105 ± 0.002 ** |
| P50H | 0.134 ± 0.011 | 0.046 ± 0.006 ** |
| S113L | 0.173 ± 0.020 | 0.036 ± 0.016 ** |
| Y479F | 0.756 ± 0.062 | 0.108 ± 0.039 ** |
| CPT II Variant | Sense Sequence (5′-3′) | Antisense Sequence (5′-3′) |
|---|---|---|
| P50H | GGACAGCCTGCATAGGCTGCCTATTC | TGGTAGTGCATGGTGGGC |
| S113L | AAGCTACATTTTGGGACCCTGGT | GTATGTTTATTCTGTTTGTCCAGAG |
| Y479F | CTGCGGCAGTTTGGGCAGACAG | GAAGGCCATCTGGAATGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meinhardt, B.; Motlagh Scholle, L.; Seifert, F.; Anwand, M.; Pietzsch, M.; Zierz, S. Cardiolipin Stabilizes and Increases Catalytic Efficiency of Carnitine Palmitoyltransferase II and Its Variants S113L, P50H, and Y479F. Int. J. Mol. Sci. 2021, 22, 4831. https://doi.org/10.3390/ijms22094831
Meinhardt B, Motlagh Scholle L, Seifert F, Anwand M, Pietzsch M, Zierz S. Cardiolipin Stabilizes and Increases Catalytic Efficiency of Carnitine Palmitoyltransferase II and Its Variants S113L, P50H, and Y479F. International Journal of Molecular Sciences. 2021; 22(9):4831. https://doi.org/10.3390/ijms22094831
Chicago/Turabian StyleMeinhardt, Beate, Leila Motlagh Scholle, Franziska Seifert, Martina Anwand, Markus Pietzsch, and Stephan Zierz. 2021. "Cardiolipin Stabilizes and Increases Catalytic Efficiency of Carnitine Palmitoyltransferase II and Its Variants S113L, P50H, and Y479F" International Journal of Molecular Sciences 22, no. 9: 4831. https://doi.org/10.3390/ijms22094831
APA StyleMeinhardt, B., Motlagh Scholle, L., Seifert, F., Anwand, M., Pietzsch, M., & Zierz, S. (2021). Cardiolipin Stabilizes and Increases Catalytic Efficiency of Carnitine Palmitoyltransferase II and Its Variants S113L, P50H, and Y479F. International Journal of Molecular Sciences, 22(9), 4831. https://doi.org/10.3390/ijms22094831