
Active Components from Cassia abbreviata Prevent HIV-1 Entry by Distinct Mechanisms of Action


 , ,
, ,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
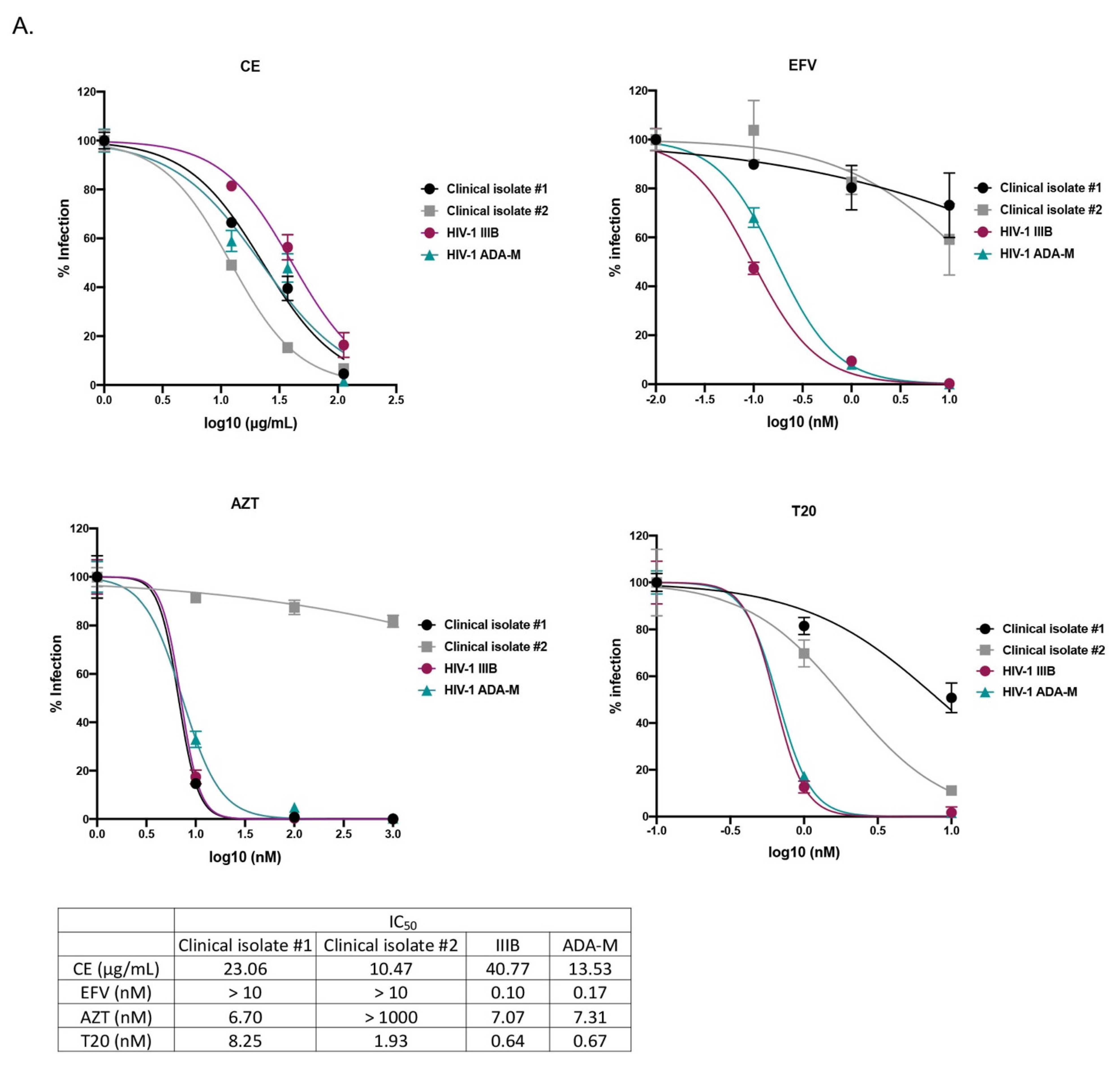
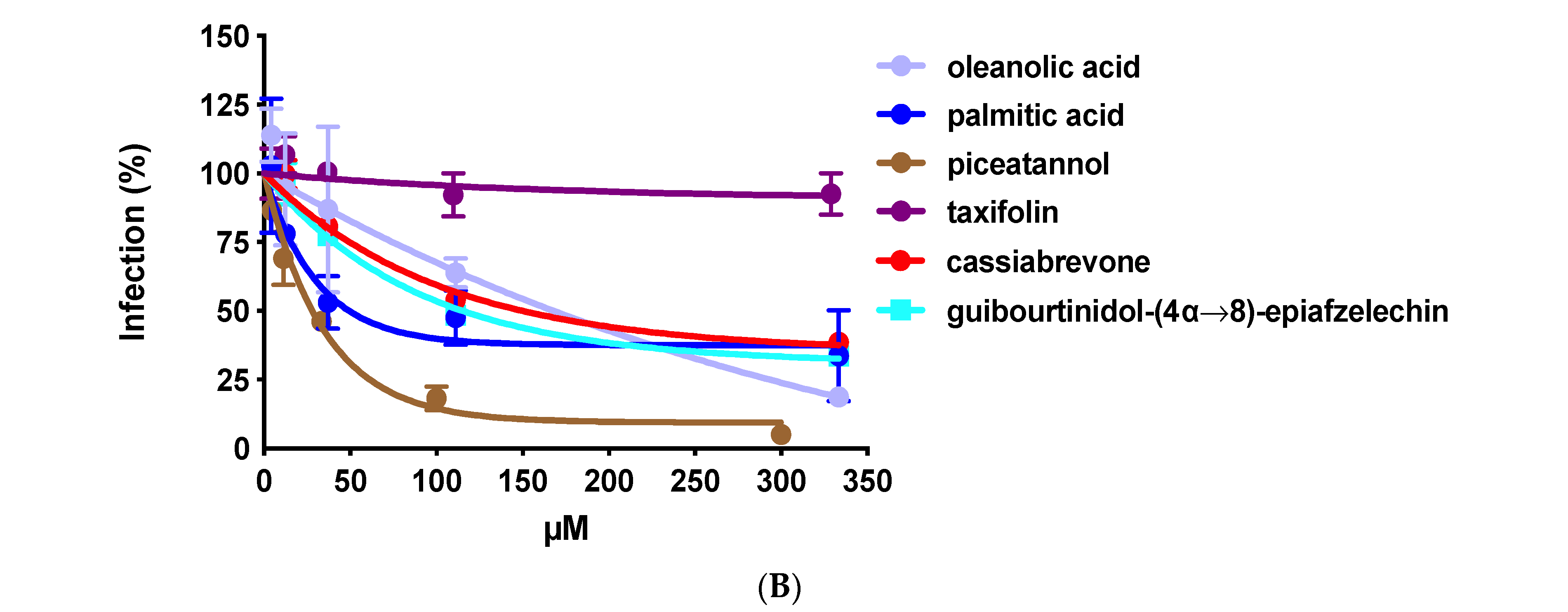
2.1. The Crude Extract and Purified Compounds of C. abbreviata Inhibit HIV-1 Entry
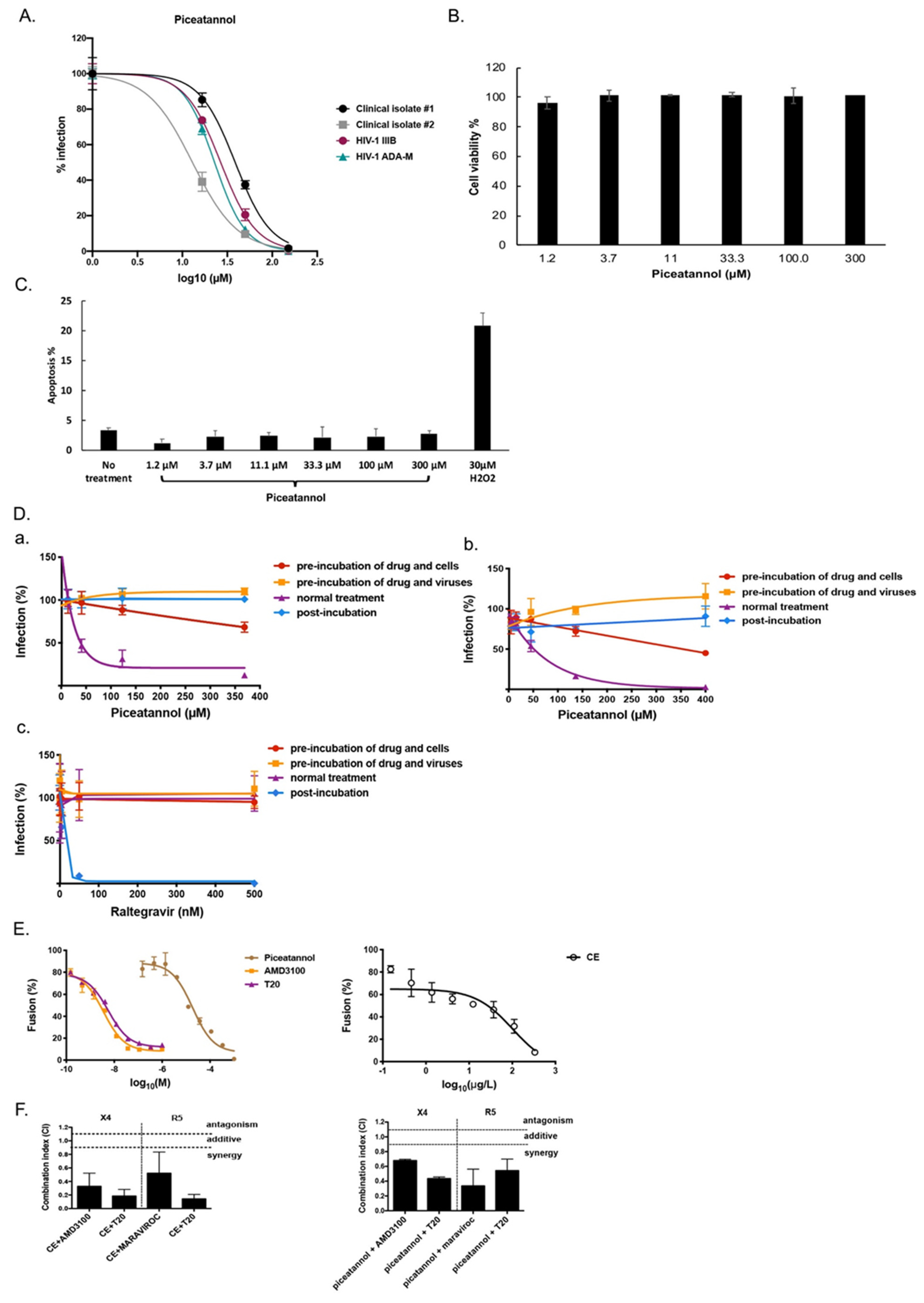
2.2. Piceatannol Interacts with Both Cell and Viral Membranes and has a Synergistic Effect with HIV-1 Entry Inhibitors
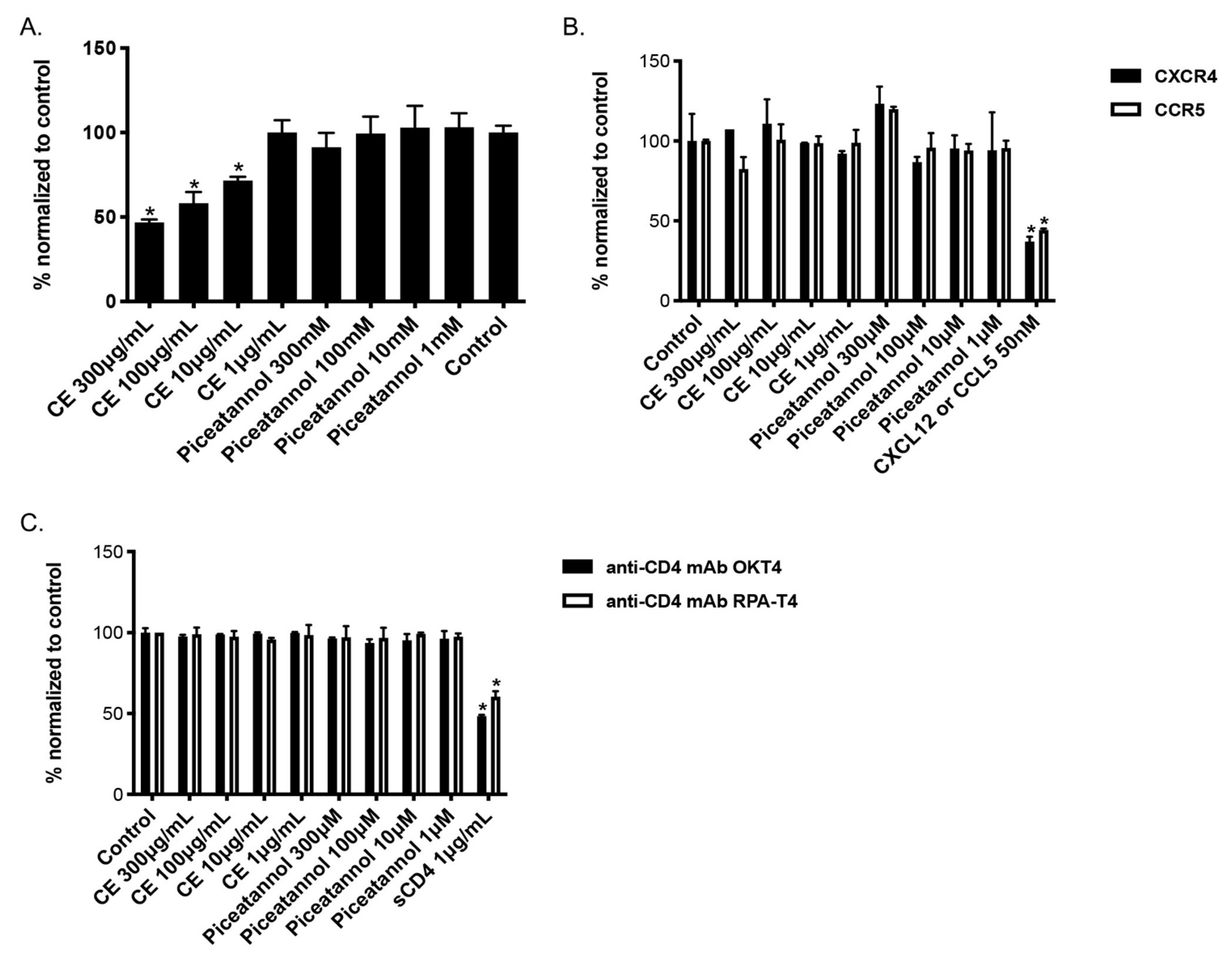
2.3. CE of C. abbreviata Affects gp120/CD4 Binding Whereas Piceatannol Interacts with Cellular Membranes
2.4. Microbicide Activity of Piceatannol
2.5. Structure-Activity Relationship of Piceatannol
2.6. Mode of Action of the Two Flavonoids Inhibiting HIV-1 Entry
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Extraction and Compounds Purification
5.2. Cell Cultures
5.3. Viral Infection with HIV-1 Reference Strains and Clinical Isolates
5.4. Cytotoxicity and Apoptosis Assays
5.5. Multi-Dosing Time Assay and Drug Combination Assay
5.6. Fusion Assay
5.7. Broad Spectrum Antiviral Activity
5.8. CD4-gp120 Interaction Assay
5.9. Binding Assays and Co-Receptor Internalization
5.10. Dual-Chamber and DC-SIGN Transmission Assays
5.11. In Silico Ligand-Based Study
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Este, J.A.; Cihlar, T. Current status and challenges of antiretroviral research and therapy. Antivir. Res. 2010, 85, 25–33. [Google Scholar] [CrossRef]
- Palmisano, L.; Vella, S. A brief history of antiretroviral therapy of HIV infection: Success and challenges. Ann. Ist. Super Sanita 2011, 47, 44–48. [Google Scholar] [PubMed]
- Glynn, J.R.; Carael, M.; Auvert, B.; Kahindo, M.; Chege, J.; Musonda, R.; Kaona, F.; Buve, A. Study Group on the Heterogeneity of, H.I.V.E. i. A. C., Why do young women have a much higher prevalence of HIV than young men? A study in Kisumu, Kenya and Ndola, Zambia. AIDS 2001, 15 (Suppl. 4), S51–S60. [Google Scholar] [CrossRef] [PubMed]
- Mehellou, Y.; De Clercq, E. Twenty-six years of anti-HIV drug discovery: Where do we stand and where do we go? J. Med. Chem. 2010, 53, 521–538. [Google Scholar] [CrossRef] [PubMed]
- Vigant, F.; Santos, N.C.; Lee, B. Broad-spectrum antivirals against viral fusion. Nat. Rev. Microbiol. 2015, 13, 426–437. [Google Scholar] [CrossRef]
- Wilen, C.B.; Tilton, J.C.; Doms, R.W. HIV: Cell binding and entry. Cold Spring Harb. Perspect. Med. 2012, 2, a006866. [Google Scholar] [CrossRef]
- St Vincent, M.R.; Colpitts, C.C.; Ustinov, A.V.; Muqadas, M.; Joyce, M.A.; Barsby, N.L.; Epand, R.F.; Epand, R.M.; Khramyshev, S.A.; Valueva, O.A.; et al. Rigid amphipathic fusion inhibitors, small molecule antiviral compounds against enveloped viruses. Proc. Natl. Acad. Sci. USA 2010, 107, 17339–17344. [Google Scholar] [CrossRef] [Green Version]
- Dias, D.A.; Urban, S.; Roessner, U. A historical overview of natural products in drug discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurice, M.I. Handbook of African Medicinal Plants, 2nd ed.; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar]
- Mongalo, N.I.; Mafoko, B.J. Cassia abbreviata Oliv, a review of its ethnomedicinal uses, toxicology, phytochemistry, possible propagation techniques and pharmacology. Afr. J. Pharm. Pharmacol. 2013, 7, 6. [Google Scholar]
- Ribeiro, A.; Romeiras, M.M.; Tavares, J.; Faria, M.T. Ethnobotanical survey in Canhane village, district of Massingir, Mozambique: Medicinal plants and traditional knowledge. J. Ethnobiol. Ethnomed. 2010, 6, 33. [Google Scholar]
- Yang, X.; He, Z.; Zheng, Y.; Wang, N.; Mulinge, M.; Schmit, J.C.; Steinmetz, A.; Seguin-Devaux, C. Chemical Constituents of Cassia abbreviata and Their Anti-HIV-1 Activity. Molecules 2021, 26, 2455. [Google Scholar] [CrossRef]
- Leteane, M.M.; Ngwenya, B.N.; Muzila, M.; Namushe, A.; Mwinga, J.; Musonda, R.; Moyo, S.; Mengestu, Y.B.; Abegaz, B.M.; Andrae-Marobela, K. Old plants newly discovered: Cassia sieberiana D.C. and Cassia abbreviata Oliv. Oliv. root extracts inhibit in vitro HIV-1c replication in peripheral blood mononuclear cells (PBMCs) by different modes of action. J. Ethnopharmacol. 2012, 141, 48–56. [Google Scholar] [CrossRef]
- Chou, T.C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [PubMed] [Green Version]
- Cirigliano, A.; Stirpe, A.; Menta, S.; Mori, M.; Dell’Edera, D.; Pick, E.; Negri, R.; Botta, B.; Rinaldi, T. Yeast as a tool to select inhibitors of the cullin deneddylating enzyme Csn5. J. Enzym. Inhib. Med. Chem. 2016, 31, 1632–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghirga, F.; Quaglio, D.; Mori, M.; Cammarone, S.; Iazzetti, A.; Goggiamani, A.; Ingallina, C.; Botta, B.; Calcaterra, A. A unique high-diversity natural product collection as a reservoir of new therapeutic leads. Org. Chem. Front. 2021, 8, 996–1025. [Google Scholar] [CrossRef]
- Infante, P.; Alfonsi, R.; Ingallina, C.; Quaglio, D.; Ghirga, F.; D’Acquarica, I.; Bernardi, F.; Di Magno, L.; Canettieri, G.; Screpanti, I.; et al. Inhibition of Hedgehog-dependent tumors and cancer stem cells by a newly identified naturally occurring chemotype. Cell Death Dis. 2016, 7, e2376. [Google Scholar]
- Infante, P.; Mori, M.; Alfonsi, R.; Ghirga, F.; Aiello, F.; Toscano, S.; Ingallina, C.; Siler, M.; Cucchi, D.; Po, A.; et al. Gli1/DNA interaction is a druggable target for Hedgehog-dependent tumors. EMBO J. 2015, 34, 200–217. [Google Scholar] [CrossRef] [PubMed]
- Mascarello, A.; Mori, M.; Chiaradia-Delatorre, L.D.; Menegatti, A.C.; Delle Monache, F.; Ferrari, F.; Yunes, R.A.; Nunes, R.J.; Terenzi, H.; Botta, B.; et al. Discovery of Mycobacterium tuberculosis protein tyrosine phosphatase B (PtpB) inhibitors from natural products. PLoS ONE 2013, 8, e77081. [Google Scholar]
- Lee, D.Y.; Lin, X.; Paskaleva, E.E.; Liu, Y.; Puttamadappa, S.S.; Thornber, C.; Drake, J.R.; Habulin, M.; Shekhtman, A.; Canki, M. Palmitic Acid Is a Novel CD4 Fusion Inhibitor That Blocks HIV Entry and Infection. Aids Res. Hum. Retrovir. 2009, 25, 1231–1241. [Google Scholar] [PubMed]
- Harada, S.; Ogihara, K.; Hikichi, Y.; Matano, T.; Narumi, K.Y. Oleanolic acid derivative OKS3-019 as a novel bifunctional HIV-1 entry inhibitor. In Proceedings of the Conference on Retroviruses and Opportunistic Infections, Seattle, WA, USA, 13–16 February 2017. [Google Scholar]
- Clouser, C.L.; Chauhan, J.; Bess, M.A.; van Oploo, J.L.; Zhou, D.; Dimick-Gray, S.; Mansky, L.M.; Patterson, S.E. Anti-HIV-1 activity of resveratrol derivatives and synergistic inhibition of HIV-1 by the combination of resveratrol and decitabine. Bioorg. Med. Chem. Lett. 2012, 22, 6642–6646. [Google Scholar]
- Kashiwada, Y.; Wang, H.K.; Nagao, T.; Kitanaka, S.; Yasuda, I.; Fujioka, T.; Yamagishi, T.; Cosentino, L.M.; Kozuka, M.; Okabe, H.; et al. Anti-AIDS agents. 30. Anti-HIV activity of oleanolic acid, pomolic acid, and structurally related triterpenoids. J. Nat. Prod. 1998, 61, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Mengoni, F.; Lichtner, M.; Battinelli, L.; Marzi, M.; Mastroianni, C.M.; Vullo, V.; Mazzanti, G. In vitro anti-HIV activity of oleanolic acid on infected human mononuclear cells. Planta Med. 2002, 68, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Min, B.S.; Lee, H.K.; Lee, S.M.; Kim, Y.H.; Bae, K.H.; Otake, T.; Nakamura, N.; Hattori, M. Anti-human immunodeficiency virus-type 1 activity of constituents from Juglans mandshurica. Arch Pharm. Res. 2002, 25, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Bar, S.; Alizon, M. Role of the ectodomain of the gp41 transmembrane envelope protein of human immunodeficiency virus type 1 in late steps of the membrane fusion process. J. Virol. 2004, 78, 811–820. [Google Scholar] [CrossRef] [Green Version]
- Gordts, S.C.; Ferir, G.; D’Huys, T.; Petrova, M.I.; Lebeer, S.; Snoeck, R.; Andrei, G.; Schols, D. The Low-Cost Compound Lignosulfonic Acid (LA) Exhibits Broad-Spectrum Anti-HIV and Anti-HSV Activity and Has Potential for Microbicidal Applications. PLoS ONE 2015, 10, e0131219. [Google Scholar] [CrossRef]
- Pasetto, S.; Pardi, V.; Murata, R.M. Anti-HIV-1 activity of flavonoid myricetin on HIV-1 infection in a dual-chamber in vitro model. PLoS ONE 2014, 9, e115323. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Ohmoto, N.; Tamotsu, K.; Tomimori, T.; Miyaichi, Y.; Shirataki, Y.; Monache, F.; Botta, B. On the relationship between the chemical structure and the cyclic AMP phosphodiesterase inhibitory activity of flavonoids as studied by carbon13 NMR. Bull. Chem. Soc. Jpn. 1989, 62, 2450–2452. [Google Scholar] [CrossRef]
- Panthong, P.; Bunluepuech, K.; Boonnak, N.; Chaniad, P.; Pianwanit, S.; Wattanapiromsakul, C.; Tewtrakul, S. Anti-HIV-1 integrase activity and molecular docking of compounds from Albizia procera bark. Pharm. Biol. 2015, 53, 1861–1866. [Google Scholar] [CrossRef] [Green Version]
- Piao, Z.S.; Feng, Y.B.; Wang, L.; Zhang, X.Q.; Lin, M. Synthesis and HIV-1 inhibitory activity of natural products isolated from Gnetum parvifolium and their analogues. Yao Xue Xue Bao 2010, 45, 1509–1515. [Google Scholar]
- Han, Y.S.; Quashie, P.K.; Mesplede, T.; Xu, H.; Quan, Y.; Jaeger, W.; Szekeres, T.; Wainberg, M.A. A resveratrol analog termed 3,3′,4,4′,5,5′-hexahydroxy-trans-stilbene is a potent HIV-1 inhibitor. J. Med. Virol. 2015, 87, 2054–2060. [Google Scholar] [CrossRef]
- Anggakusuma Colpitts, C.C.; Schang, L.M.; Rachmawati, H.; Frentzen, A.; Pfaender, S.; Behrendt, P.; Brown, R.J.; Bankwitz, D.; Steinmann, J.; Ott, M.; et al. Turmeric curcumin inhibits entry of all hepatitis C virus genotypes into human liver cells. Gut 2014, 63, 1137–1149. [Google Scholar] [CrossRef]
- Wolf, M.C.; Freiberg, A.N.; Zhang, T.; Akyol-Ataman, Z.; Grock, A.; Hong, P.W.; Li, J.; Watson, N.F.; Fang, A.Q.; Aguilar, H.C.; et al. A broad-spectrum antiviral targeting entry of enveloped viruses. Proc. Natl. Acad. Sci. USA 2010, 107, 3157–3162. [Google Scholar] [CrossRef] [Green Version]
- Docherty, J.J.; Fu, M.M.; Hah, J.M.; Sweet, T.J.; Faith, S.A.; Booth, T. Effect of resveratrol on herpes simplex virus vaginal infection in the mouse. Antivir. Res. 2005, 67, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Malancona, S.; Mori, M.; Fezzardi, P.; Santoriello, M.; Basta, A.; Nibbio, M.; Kovalenko, L.; Speziale, R.; Battista, M.R.; Cellucci, A.; et al. 5,6-Dihydroxypyrimidine Scaffold to Target HIV-1 Nucleocapsid Protein. ACS Med. Chem. Lett 2020, 11, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Ciaco, S.; Mely, Y.; Karioti, A. Inhibitory Effect of Lithospermic Acid on the HIV-1 Nucleocapsid Protein. Molecules 2020, 25, 5434. [Google Scholar] [CrossRef] [PubMed]
- Huth, J.R.; Mendoza, R.; Olejniczak, E.T.; Johnson, R.W.; Cothron, D.A.; Liu, Y.; Lerner, C.G.; Chen, J.; Hajduk, P.J. ALARM NMR: A rapid and robust experimental method to detect reactive false positives in biochemical screens. J. Am. Chem. Soc. 2005, 127, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Kovalenko, L.; Malancona, S.; Saladini, F.; De Forni, D.; Pires, M.; Humbert, N.; Real, E.; Botzanowski, T.; Cianferani, S.; et al. Structure-Based Identification of HIV-1 Nucleocapsid Protein Inhibitors Active against Wild-Type and Drug-Resistant HIV-1 Strains. ACS Chem. Biol. 2018, 13, 253–266. [Google Scholar] [CrossRef]
- Baeten, J.M.; Benki, S.; Chohan, V.; Lavreys, L.; McClelland, R.S.; Mandaliya, K.; Ndinya-Achola, J.O.; Jaoko, W.; Overbaugh, J. Hormonal contraceptive use, herpes simplex virus infection, and risk of HIV-1 acquisition among Kenyan women. AIDS 2007, 21, 1771–1777. [Google Scholar] [CrossRef] [PubMed]
- Baatz, F.; Nijhuis, M.; Lemaire, M.; Riedijk, M.; Wensing, A.M.; Servais, J.Y.; van Ham, P.M.; Hoepelman, A.I.; Koopmans, P.P.; Sprenger, H.G.; et al. Impact of the HIV-1 env genetic context outside HR1-HR2 on resistance to the fusion inhibitor enfuvirtide and viral infectivity in clinical isolates. PLoS ONE 2011, 6, e21535. [Google Scholar] [CrossRef] [Green Version]
- Bargellini, G.; Marini-Bettolo, G.B. Flavone, flavanone and flavonol derivatives of hydroxyhydroquinone. Gazz. Chim. Ital. 1940, 70, 170–178. [Google Scholar]
- Hawkins, P.C.; Skillman, A.G.; Nicholls, A. Comparison of shape-matching and docking as virtual screening tools. J. Med. Chem. 2007, 50, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, P.C.; Skillman, A.G.; Warren, G.L.; Ellingson, B.A.; Stahl, M.T. Conformer generation with OMEGA: Algorithm and validation using high quality structures from the Protein Databank and Cambridge Structural Database. J. Chem. Inf. Model 2010, 50, 572–584. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cells | Viruses | CE (µg/mL) | Piceatannol(µM) |
|---|---|---|---|
| HEL | Herpes simplex virus-1 (KOS) | 46.7 ± 2.9 | 47.5 ± 3.5 |
| HEL | Herpes simplex virus-2 (G) | 39.5 ± 5.5 | 45.0 ± 1.8 |
| HEL | Herpes simplex virus-1 TK KOS ACV | 45.0 ± 2.6 | 45.4 ± 4.0 |
| U87 | Vesicular stomatitis virus | >100 | 79.2± 17 |
| MDCK | Influenza A/H1N1 A/Ned/378/05 | >100 | >100 |
| MDCK | Influenza A/H3N2 A/HK/7/87 | >100 | >100 |
| MDCK | Influenza B B/Ned/537/05 | >100 | >100 |
| Huh 7-D | Hepatitis C virus (Jc1) | >100 | >100 |
| HeLa | Coxsackie virus B4 | >100 | >100 |
| vero | Coxsackie virus B4 | >100 | >100 |
| HeLa | Respiratory syncytial virus | >100 | >100 |
| vero | Para-influenza-3 virus | >100 | >100 |
| vero | Reovirus-1 | >100 | >100 |
| vero | sindbisvirus | >100 | >100 |
| vero | Punta toro virus | >100 | >100 |
| vero | Yellow fever virus | >100 | >100 |
| CRFK | Feline corona virus (FIPV) | >100 | >100 |
| CRFK | Feline herpes virus | >100 | >100 |
| HEL | Human corona virus | >100 | >100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Yang, X.-W.; Schols, D.; Mori, M.; Botta, B.; Chevigné, A.; Mulinge, M.; Steinmetz, A.; Schmit, J.-C.; Seguin-Devaux, C. Active Components from Cassia abbreviata Prevent HIV-1 Entry by Distinct Mechanisms of Action. Int. J. Mol. Sci. 2021, 22, 5052. https://doi.org/10.3390/ijms22095052
Zheng Y, Yang X-W, Schols D, Mori M, Botta B, Chevigné A, Mulinge M, Steinmetz A, Schmit J-C, Seguin-Devaux C. Active Components from Cassia abbreviata Prevent HIV-1 Entry by Distinct Mechanisms of Action. International Journal of Molecular Sciences. 2021; 22(9):5052. https://doi.org/10.3390/ijms22095052
Chicago/Turabian StyleZheng, Yue, Xian-Wen Yang, Dominique Schols, Mattia Mori, Bruno Botta, Andy Chevigné, Martin Mulinge, André Steinmetz, Jean-Claude Schmit, and Carole Seguin-Devaux. 2021. "Active Components from Cassia abbreviata Prevent HIV-1 Entry by Distinct Mechanisms of Action" International Journal of Molecular Sciences 22, no. 9: 5052. https://doi.org/10.3390/ijms22095052
APA StyleZheng, Y., Yang, X. -W., Schols, D., Mori, M., Botta, B., Chevigné, A., Mulinge, M., Steinmetz, A., Schmit, J. -C., & Seguin-Devaux, C. (2021). Active Components from Cassia abbreviata Prevent HIV-1 Entry by Distinct Mechanisms of Action. International Journal of Molecular Sciences, 22(9), 5052. https://doi.org/10.3390/ijms22095052