
Myoparr-Associated and -Independent Multiple Roles of Heterogeneous Nuclear Ribonucleoprotein K during Skeletal Muscle Cell Differentiation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
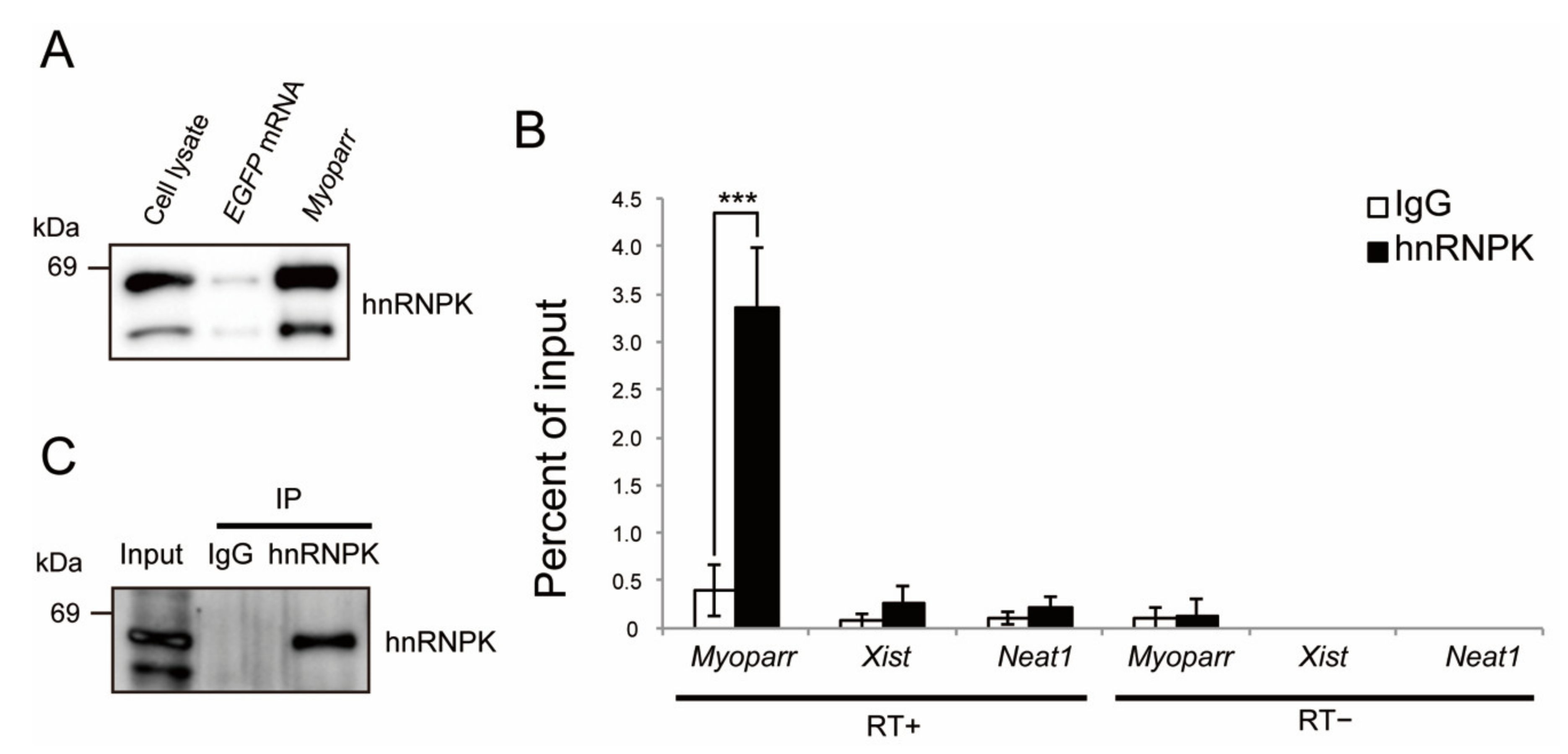
2.1. Identification of hnRNPK as a Myoparr-Binding Protein in Skeletal Muscle Cells
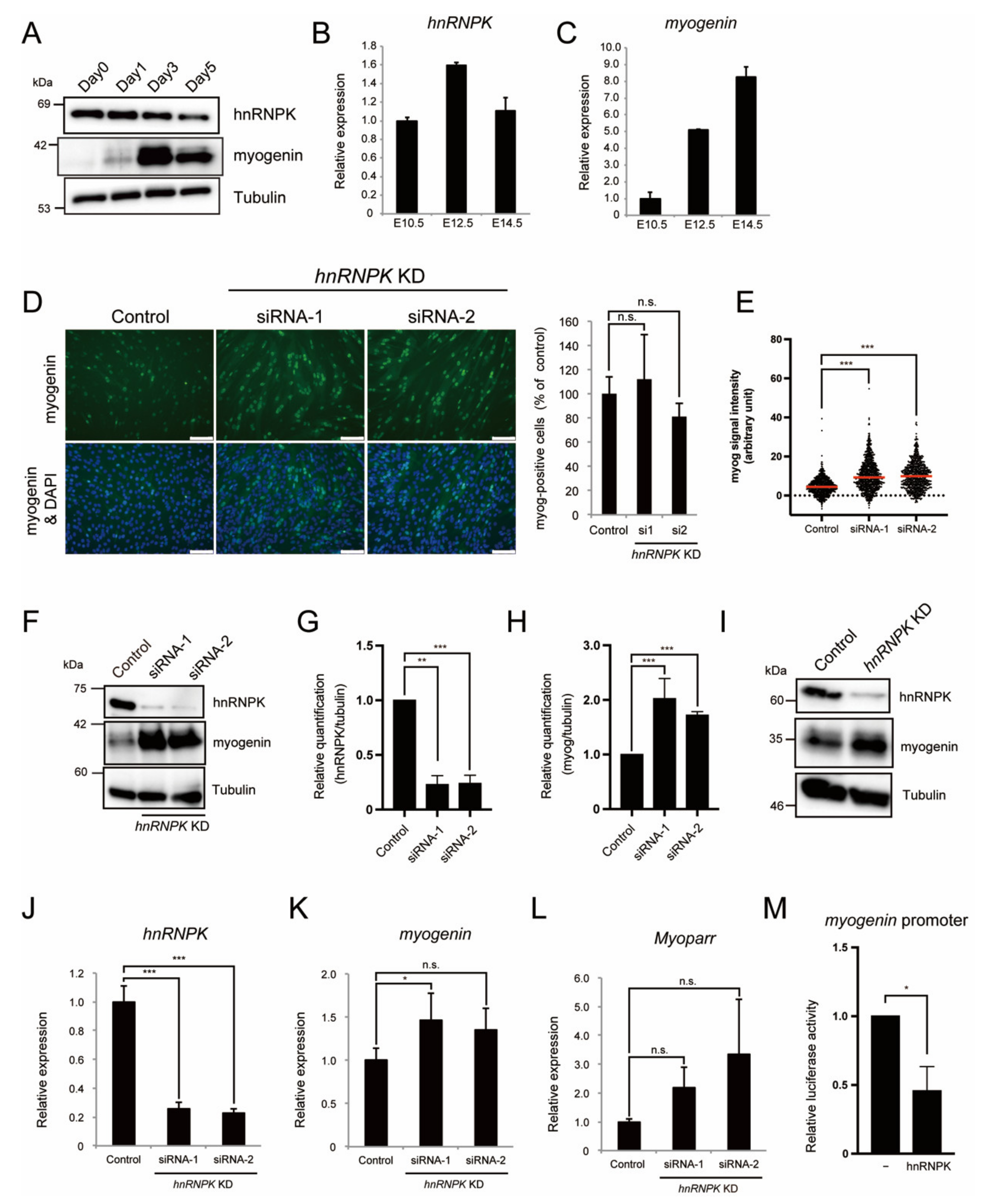
2.2. hnRNPK Represses the Expression of Myogenin in Differentiating Myoblasts
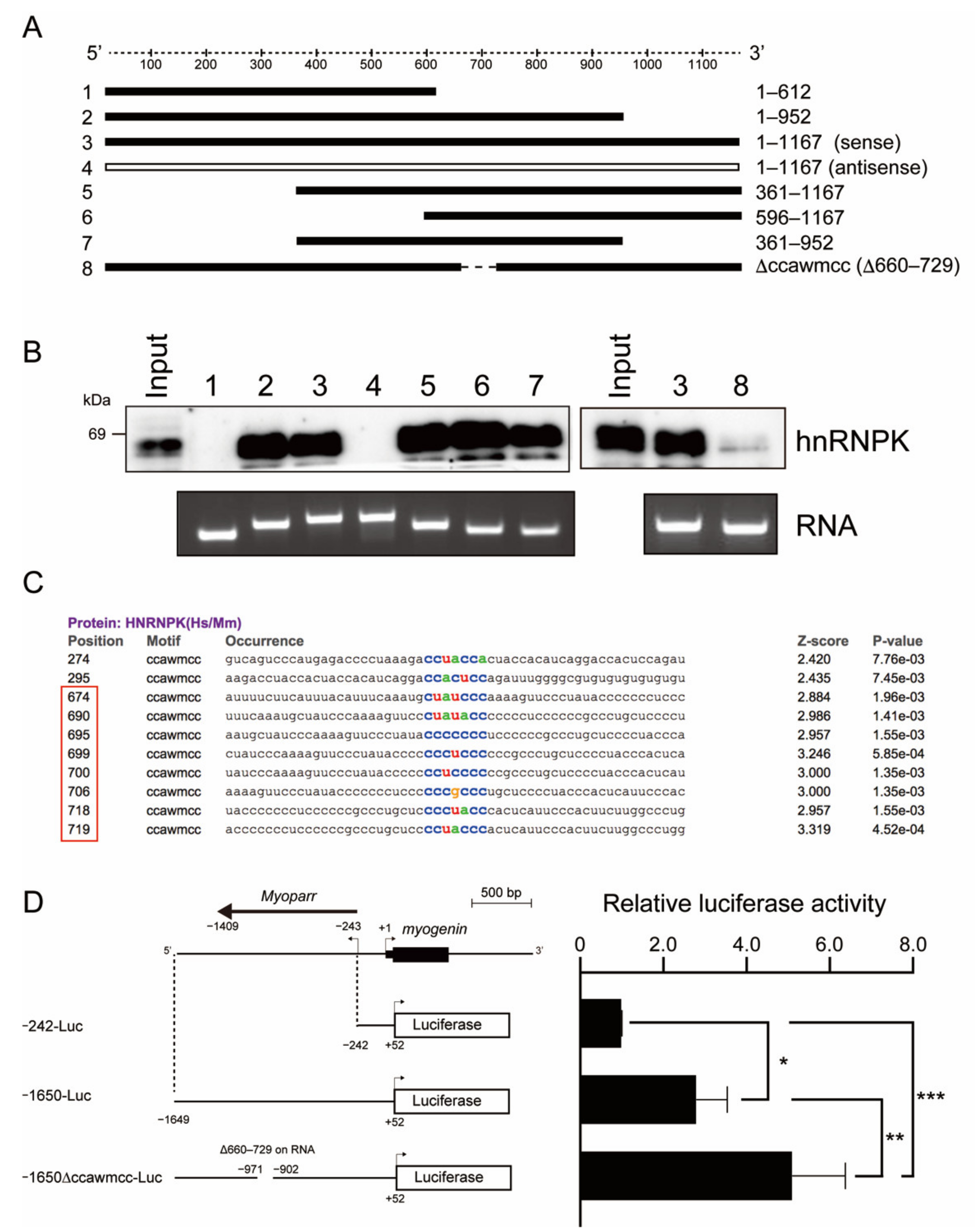
2.3. The hnRNPK-Binding Region of Myoparr Is Required to Repress Myogenin Expression
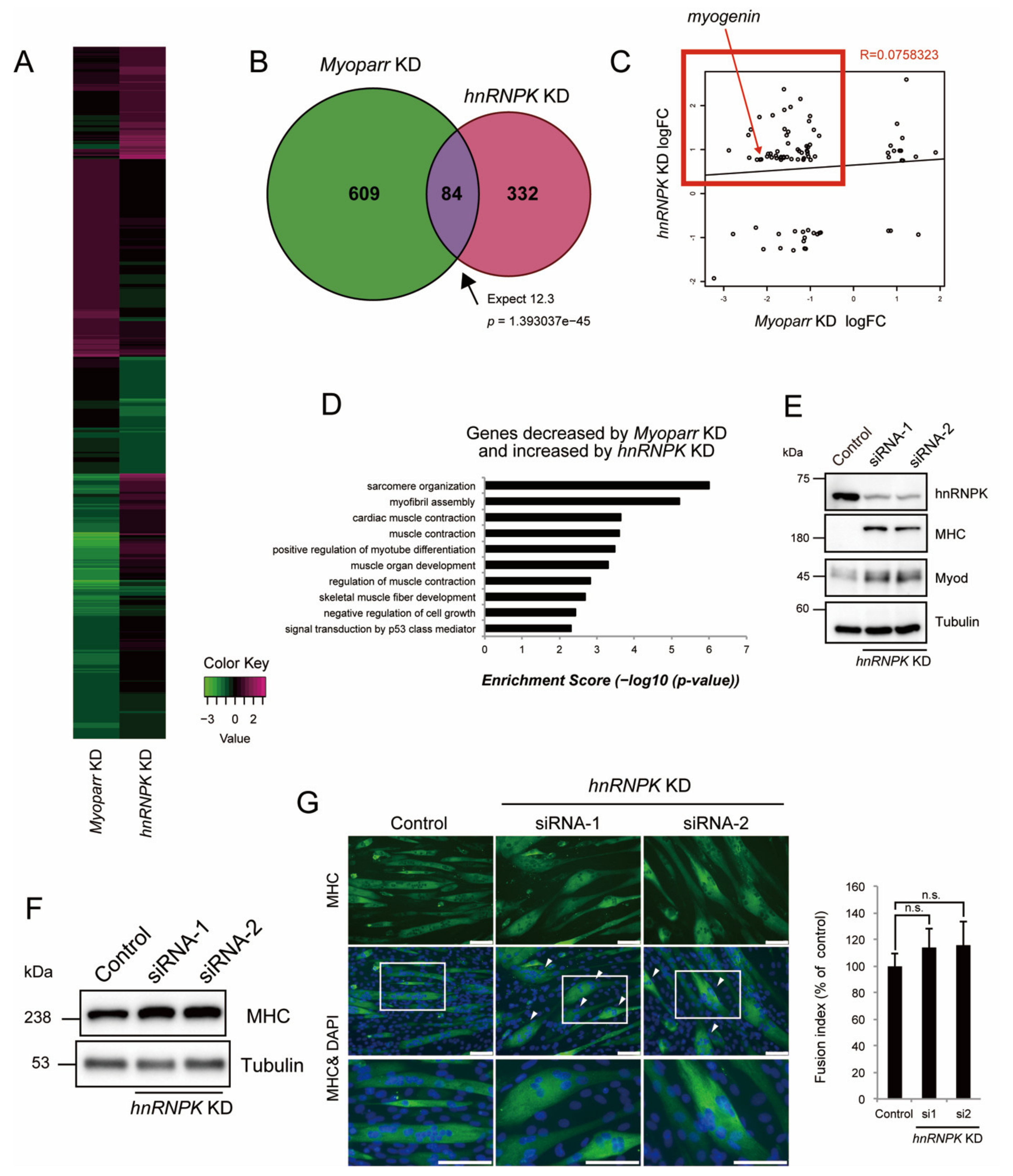
2.4. hnRNPK Inhibits Skeletal Muscle Differentiation but Is Required for Normal Myotube Formation
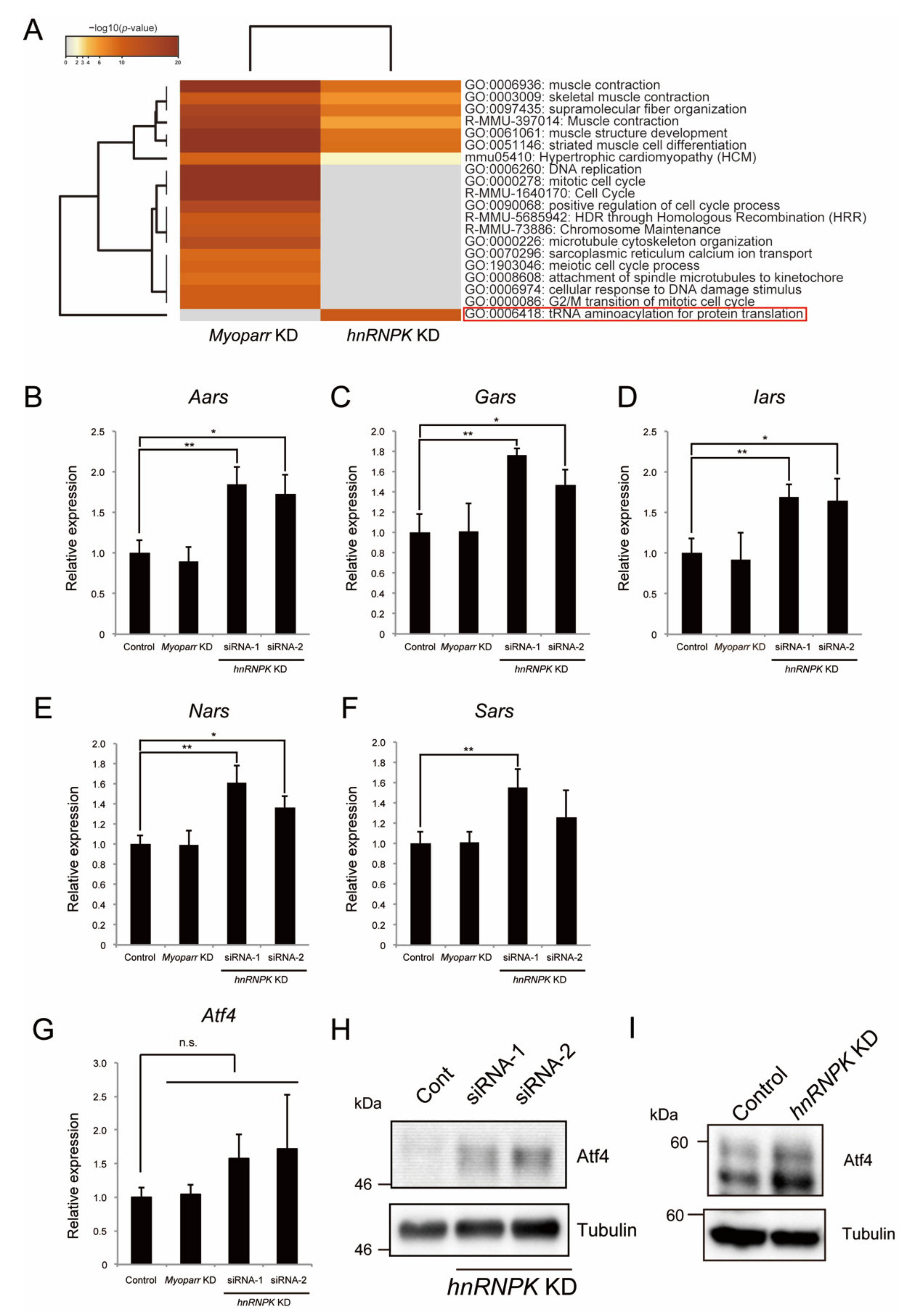
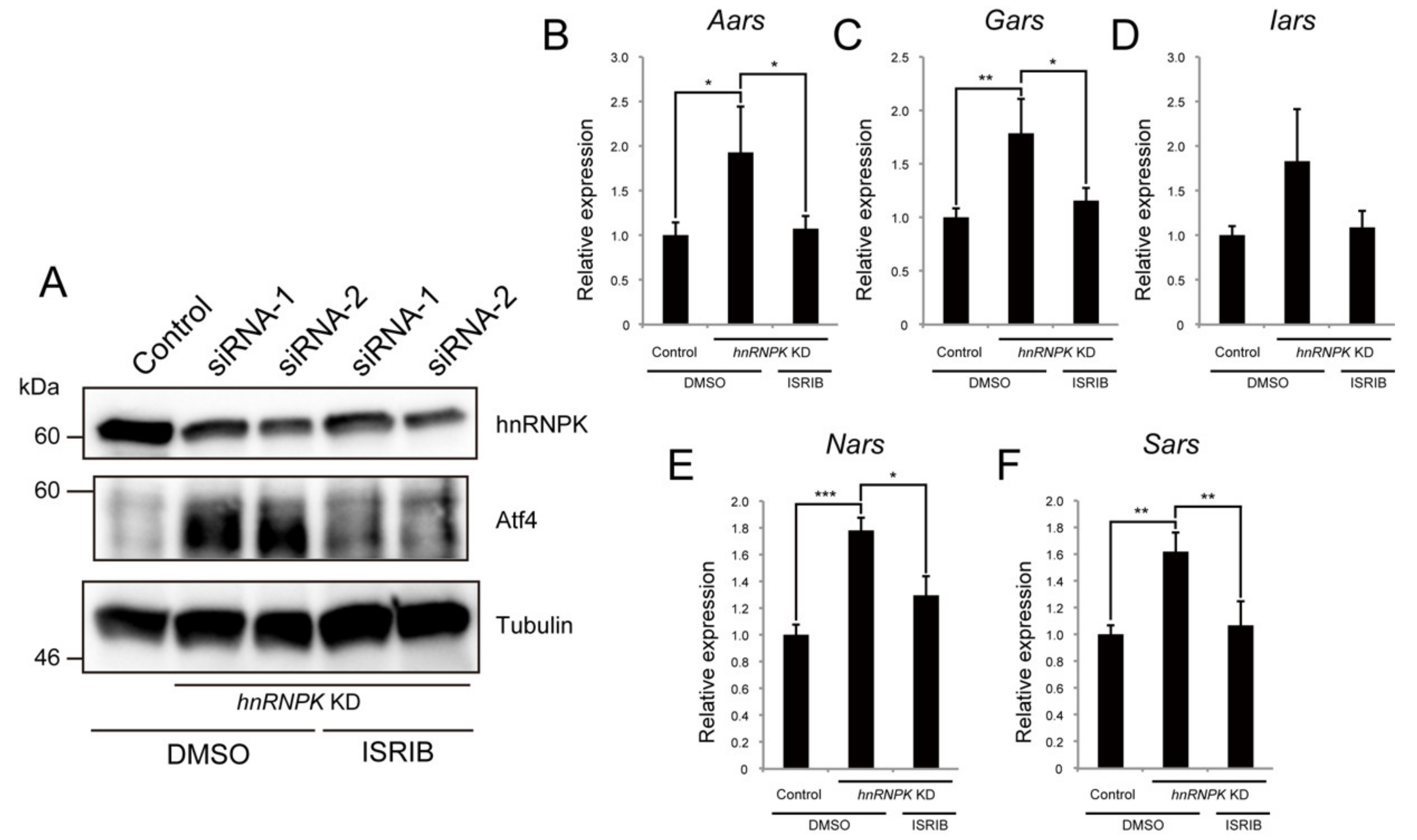
2.5. hnRNPK Represses the Expression of Aminoacyl-tRNA Synthetases via the eIF2α/Atf4 Pathway
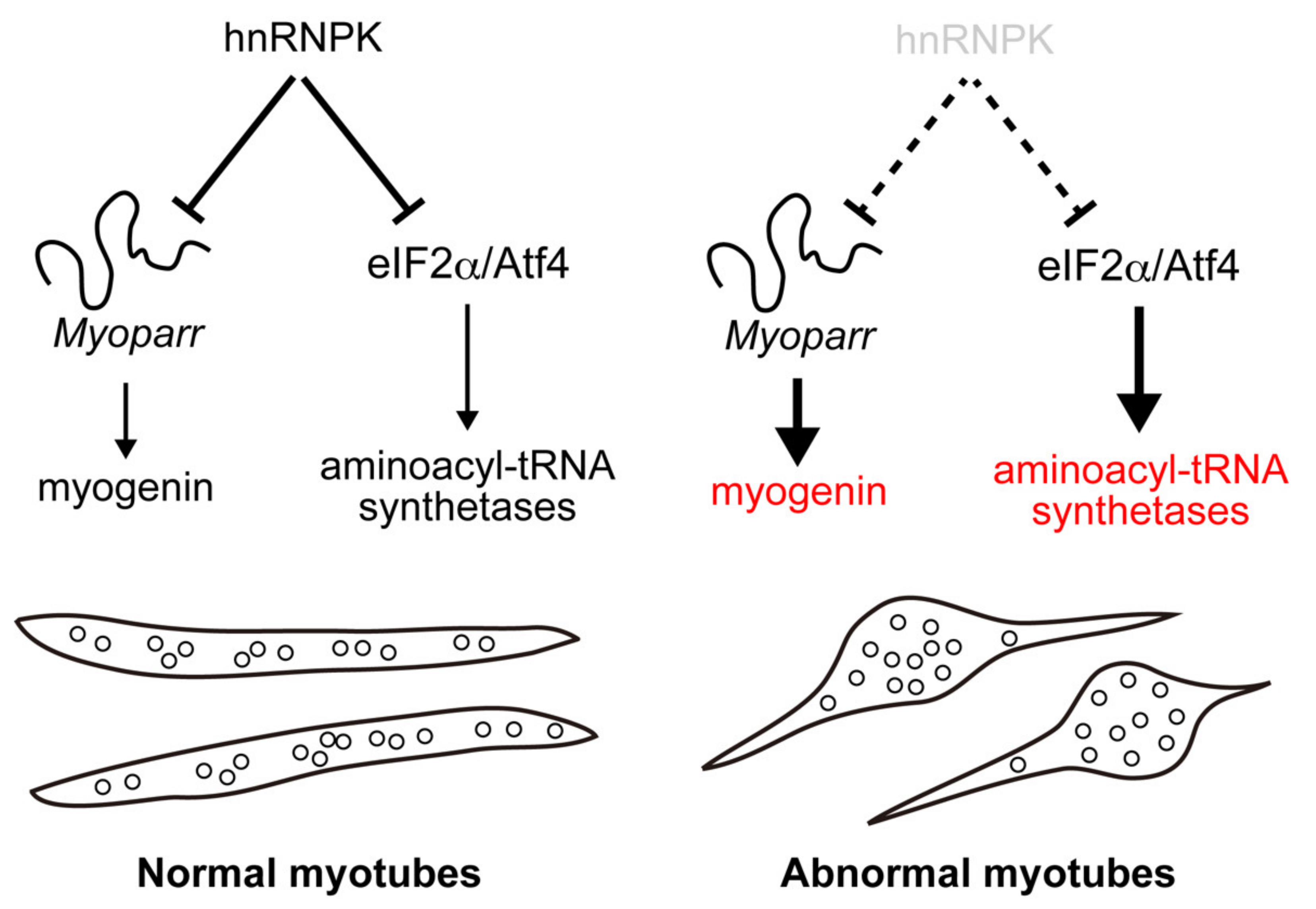
3. Discussion
4. Materials and Methods
4.1. Cell Cultures, siRNA Transfection, and ISRIB Treatment
4.2. RNA Isolation, Reverse Transcription Reaction, and Quantitative RT-PCR
4.3. Protein Extraction and Western Blotting
4.4. Immunofluorescence Assay
4.5. Identification of Myoparr-Binding Proteins
4.6. RNA Immunoprecipitation and the RNA Pull-Down Assay
4.7. Luciferase Reporter Assay
4.8. Analysis of Downstream Genes Regulated by Myoparr KD and hnRNPK KD
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Deb, A.; Maji, R.K.; Saha, S.; Ghosh, Z. LncRBase: An Enriched Resource for LncRNA Information. PLoS ONE 2014, 9, e108010. [Google Scholar]
- Hon, C.-C.; Ramilowski, J.A.; Harshbarger, J.; Bertin, N.; Rackham, O.J.L.; Gough, J.; Denisenko, E.; Schmeier, S.; Poulsen, T.M.; Severin, J.; et al. An Atlas of Human Long Non-Coding RNAs with Accurate 5′ Ends. Nature 2017, 543, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Cao, J.; Liu, L.; Du, Q.; Li, Z.; Zou, D.; Bajic, V.B.; Zhang, Z. LncBook: A Curated Knowledgebase of Human Long Non-Coding RNAs. Nucleic Acids Res. 2019, 47, D128–D134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hitachi, K.; Tsuchida, K. The Chemical Biology of Long Noncoding RNAs. Rna Technol. 2020, 11, 431–463. [Google Scholar]
- Gerstberger, S.; Hafner, M.; Tuschl, T. A Census of Human RNA-Binding Proteins. Nat. Rev. Genet. 2014, 15, 829–845. [Google Scholar] [CrossRef] [PubMed]
- Caretti, G.; Schiltz, R.L.; Dilworth, F.J.; Padova, M.D.; Zhao, P.; Ogryzko, V.; Fuller-Pace, F.V.; Hoffman, E.P.; Tapscott, S.J.; Sartorelli, V. The RNA Helicases P68/P72 and the Noncoding RNA SRA Are Coregulators of MyoD and Skeletal Muscle Differentiation. Dev. Cell 2006, 11, 547–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hitachi, K.; Nakatani, M.; Takasaki, A.; Ouchi, Y.; Uezumi, A.; Ageta, H.; Inagaki, H.; Kurahashi, H.; Tsuchida, K. Myogenin Promoter-associated LncRNA Myoparr Is Essential for Myogenic Differentiation. EMBO Rep. 2019, 20, e47468. [Google Scholar] [CrossRef] [PubMed]
- Kataruka, S.; Akhade, V.S.; Kayyar, B.; Rao, M.R.S. Mrhl Long Noncoding RNA Mediates Meiotic Commitment of Mouse Spermatogonial Cells by Regulating Sox8 Expression. Mol. Cell Biol. 2017, 37, e00632-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallam, T.; Jones, M.; Thomas, B.J.; Wu, X.; Gilliland, T.; Qian, K.; Eskin, A.; Casero, D.; Zhang, Z.; Sandhu, J.; et al. Transcriptional Regulation of Macrophage Cholesterol Efflux and Atherogenesis by a Long Noncoding RNA. Nat. Med. 2018, 24, 304–312. [Google Scholar] [CrossRef]
- Hirose, T.; Yamazaki, T.; Nakagawa, S. Molecular Anatomy of the Architectural NEAT1 Noncoding RNA: The Domains, Interactors, and Biogenesis Pathway Required to Build Phase-separated Nuclear Paraspeckles. Wiley Interdiscip. Rev. RNA 2019, 10, e1545. [Google Scholar] [CrossRef]
- Legnini, I.; Morlando, M.; Mangiavacchi, A.; Fatica, A.; Bozzoni, I. A Feedforward Regulatory Loop between HuR and the Long Noncoding RNA Linc-MD1 Controls Early Phases of Myogenesis. Mol. Cell 2014, 53, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.-H.; Abdelmohsen, K.; Srikantan, S.; Yang, X.; Martindale, J.L.; De, S.; Huarte, M.; Zhan, M.; Becker, K.G.; Gorospe, M. LincRNA-P21 Suppresses Target MRNA Translation. Mol. Cell 2012, 47, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelaini, S.; Chan, C.; Cornelius, V.A.; Margariti, A. RNA-Binding Proteins Hold Key Roles in Function, Dysfunction, and Disease. Biology 2021, 10, 366. [Google Scholar] [CrossRef] [PubMed]
- Briata, P.; Gherzi, R. Long Non-Coding RNA-Ribonucleoprotein Networks in the Post-Transcriptional Control of Gene Expression. Non-Coding RNA 2020, 6, 40. [Google Scholar] [CrossRef] [PubMed]
- Jonas, K.; Calin, G.A.; Pichler, M. RNA-Binding Proteins as Important Regulators of Long Non-Coding RNAs in Cancer. Int. J. Mol. Sci. 2020, 21, 2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nostrand, E.L.V.; Freese, P.; Pratt, G.A.; Wang, X.; Wei, X.; Xiao, R.; Blue, S.M.; Chen, J.-Y.; Cody, N.A.L.; Dominguez, D.; et al. A Large-Scale Binding and Functional Map of Human RNA-Binding Proteins. Nature 2020, 583, 711–719. [Google Scholar] [CrossRef]
- Gebauer, F.; Schwarzl, T.; Valcárcel, J.; Hentze, M.W. RNA-Binding Proteins in Human Genetic Disease. Nat. Rev. Genet. 2021, 22, 185–198. [Google Scholar] [CrossRef]
- Bomsztyk, K.; Denisenko, O.; Ostrowski, J. HnRNP K: One Protein Multiple Processes. Bioessays 2004, 26, 629–638. [Google Scholar] [CrossRef]
- Wang, Z.; Qiu, H.; He, J.; Liu, L.; Xue, W.; Fox, A.; Tickner, J.; Xu, J. The Emerging Roles of HnRNPK. J. Cell Physiol. 2020, 235, 1995–2008. [Google Scholar] [CrossRef]
- Dimitrova, N.; Zamudio, J.R.; Jong, R.M.; Soukup, D.; Resnick, R.; Sarma, K.; Ward, A.J.; Raj, A.; Lee, J.T.; Sharp, P.A.; et al. LincRNA-P21 Activates P21 In Cis to Promote Polycomb Target Gene Expression and to Enforce the G1/S Checkpoint. Mol. Cell 2014, 54, 777–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howarth, M.M.; Simpson, D.; Ngok, S.P.; Nieves, B.; Chen, R.; Siprashvili, Z.; Vaka, D.; Breese, M.R.; Crompton, B.D.; Alexe, G.; et al. Long Noncoding RNA EWSAT1-Mediated Gene Repression Facilitates Ewing Sarcoma Oncogenesis. J. Clin. Investig. 2014, 124, 5275–5290. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Xie, Z.; Wang, P.; Li, J.; Wang, S.; Liu, W.; Li, M.; Wu, X.; Su, H.; Cen, S.; et al. LncRNA-OG Promotes the Osteogenic Differentiation of Bone Marrow-Derived Mesenchymal Stem Cells Under the Regulation of HnRNPK. Stem. Cells 2019, 37, 270–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, C.; Zhang, Q.C.; da Rocha, S.T.; Flynn, R.A.; Bharadwaj, M.; Calabrese, J.M.; Magnuson, T.; Heard, E.; Chang, H.Y. Systematic Discovery of Xist RNA Binding Proteins. Cell 2015, 161, 404–416. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, T.; Tanigawa, A.; Naganuma, T.; Ohkawa, Y.; Souquere, S.; Pierron, G.; Hirose, T. SWI/SNF Chromatin-Remodeling Complexes Function in Noncoding RNA-Dependent Assembly of Nuclear Bodies. Proc. Natl. Acad. Sci. USA 2015, 112, 4304–4309. [Google Scholar] [CrossRef] [Green Version]
- Makeyev, A.V.; Liebhaber, S.A. The Poly(C)-Binding Proteins: A Multiplicity of Functions and a Search for Mechanisms. RNA 2002, 8, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Liu, H.; Chen, X.; Kirby, M.; Brown, P.O.; Zhao, K. Regulation of CSF1 Promoter by the SWI/SNF-like BAF Complex. Cell 2001, 106, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Hitachi, K.; Tsuchida, K. Data Describing the Effects of Depletion of Myoparr, Myogenin, Ddx17, and HnRNPK in Differentiating C2C12 Cells. Data Brief. 2019, 25, 104172. [Google Scholar] [CrossRef]
- Harding, H.P.; Zhang, Y.; Zeng, H.; Novoa, I.; Lu, P.D.; Calfon, M.; Sadri, N.; Yun, C.; Popko, B.; Paules, R.; et al. An Integrated Stress Response Regulates Amino Acid Metabolism and Resistance to Oxidative Stress. Mol. Cell 2003, 11, 619–633. [Google Scholar] [CrossRef]
- Shan, J.; Zhang, F.; Sharkey, J.; Tang, T.A.; Örd, T.; Kilberg, M.S. The C/Ebp-Atf Response Element (CARE) Location Reveals Two Distinct Atf4-Dependent, Elongation-Mediated Mechanisms for Transcriptional Induction of Aminoacyl-TRNA Synthetase Genes in Response to Amino Acid Limitation. Nucleic Acids Res. 2016, 44, 9719–9732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afroze, D.; Kumar, A. ER Stress in Skeletal Muscle Remodeling and Myopathies. FEBS J. 2017, 286, 379–398. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Li, R.; Zhang, K.; Wu, W.; Wang, S.; Zhang, P.; Xu, H. The Multifunctional RNA-Binding Protein HnRNPK Is Critical for the Proliferation and Differentiation of Myoblasts. BMB Rep. 2018, 51, 350–355. [Google Scholar] [CrossRef] [Green Version]
- Bröhl, D.; Vasyutina, E.; Czajkowski, M.T.; Griger, J.; Rassek, C.; Rahn, H.-P.; Purfürst, B.; Wende, H.; Birchmeier, C. Colonization of the Satellite Cell Niche by Skeletal Muscle Progenitor Cells Depends on Notch Signals. Dev. Cell 2012, 23, 469–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster-Gossler, K.; Cordes, R.; Gossler, A. Premature Myogenic Differentiation and Depletion of Progenitor Cells Cause Severe Muscle Hypotrophy in Delta1 Mutants. Proc. Natl. Acad. Sci. USA 2007, 104, 537–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Cui, J.; Wong, W.M.; Li, X.; Xue, W.; Lin, R.; Wang, J.; Wang, P.; Tanner, J.A.; Cheah, K.S.E.; et al. Kif5b Controls the Localization of Myofibril Components for Their Assembly and Linkage to the Myotendinous Junctions. Development 2013, 140, 617–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelis, M.G.C.-D.; Lyons, G.; Sonnino, C.; Angelis, L.D.; Vivarelli, E.; Farmer, K.; Wright, W.E.; Molinaro, M.; Bouchè, M.; Buckingham, M. MyoD, Myogenin Independent Differentiation of Primordial Myoblasts in Mouse Somites. J. Cell Biol. 1992, 116, 1243–1255. [Google Scholar] [CrossRef] [Green Version]
- Hetz, C.; Chevet, E.; Oakes, S.A. Proteostasis Control by the Unfolded Protein Response. Nat. Cell Biol. 2015, 17, 829–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, K.; Sudo, T.; Morishima, N. Endoplasmic Reticulum Stress Signaling Transmitted by ATF6 Mediates Apoptosis during Muscle Development. J. Cell Biol. 2005, 169, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Alter, J.; Bengal, E. Stress-Induced C/EBP Homology Protein (CHOP) Represses MyoD Transcription to Delay Myoblast Differentiation. PLoS ONE 2011, 6, e29498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, K.; Dohmae, N.; Morishima, N. Endoplasmic Reticulum Stress Increases Myofiber Formation in Vitro. FASEB J. 2007, 21, 2994–3003. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Hindi, S.M.; Mann, A.K.; Gallot, Y.S.; Bohnert, K.R.; Cavener, D.R.; Whittemore, S.R.; Kumar, A. The PERK Arm of the Unfolded Protein Response Regulates Satellite Cell-Mediated Skeletal Muscle Regeneration. Elife 2017, 6, e22871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, M.; Chen, L.; Ravitz, M.J.; Mehtani, S.; Korenblat, K.; Pazin, M.J.; Schmidt, E.V. HnRNP K Binds a Core Polypyrimidine Element in the Eukaryotic Translation Initiation Factor 4E (EIF4E) Promoter, and Its Regulation of EIF4E Contributes to Neoplastic Transformation. Mol. Cell Biol. 2005, 25, 6436–6453. [Google Scholar] [CrossRef] [Green Version]
- Yano, M.; Okano, H.J.; Okano, H. Involvement of Hu and Heterogeneous Nuclear Ribonucleoprotein K in Neuronal Differentiation through P21 MRNA Post-Transcriptional Regulation. J. Biol. Chem. 2005, 280, 12690–12699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulmi, J.J.; Hentilä, J.; DeRuisseau, K.C.; Oliveira, B.M.; Papaioannou, K.G.; Autio, R.; Kujala, U.M.; Ritvos, O.; Kainulainen, H.; Korkmaz, A.; et al. Effects of Muscular Dystrophy, Exercise and Blocking Activin Receptor IIB Ligands on the Unfolded Protein Response and Oxidative Stress. Free Radic. Bio. Med. 2016, 99, 308–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikezoe, K.; Nakamori, M.; Furuya, H.; Arahata, H.; Kanemoto, S.; Kimura, T.; Imaizumi, K.; Takahashi, M.P.; Sakoda, S.; Fujii, N.; et al. Endoplasmic Reticulum Stress in Myotonic Dystrophy Type 1 Muscle. Acta Neuropathol. 2007, 114, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, L.; Bonne, G.; Rivier, F.; Hamroun, D. The 2021 Version of the Gene Table of Neuromuscular Disorders (Nuclear Genome). Neuromuscul. Disord. 2020, 30, 1008–1048. [Google Scholar] [CrossRef] [PubMed]
- Targoff, I.N.; Trieu, E.P.; Miller, F.W. Reaction of Anti-OJ Autoantibodies with Components of the Multi-Enzyme Complex of Aminoacyl-TRNA Synthetases in Addition to Isoleucyl-TRNA Synthetase. J. Clin. Investig. 1993, 91, 2556–2564. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, K.; Kitajima, Y.; Okazaki, N.; Chiba, K.; Yonekura, A.; Ono, Y. A Modified Pre-Plating Method for High-Yield and High-Purity Muscle Stem Cell Isolation from Human/Mouse Skeletal Muscle Tissues. Front. Cell Dev. Biol. 2020, 8, 793. [Google Scholar] [CrossRef] [PubMed]
- Paz, I.; Kosti, I.; Ares, M.; Cline, M.; Mandel-Gutfreund, Y. RBPmap: A Web Server for Mapping Binding Sites of RNA-Binding Proteins. Nucleic Acids Res. 2014, 42, W361–W367. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating Viruses and Cellular Organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape Provides a Biologist-Oriented Resource for the Analysis of Systems-Level Datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hitachi, K.; Kiyofuji, Y.; Nakatani, M.; Tsuchida, K. Myoparr-Associated and -Independent Multiple Roles of Heterogeneous Nuclear Ribonucleoprotein K during Skeletal Muscle Cell Differentiation. Int. J. Mol. Sci. 2022, 23, 108. https://doi.org/10.3390/ijms23010108
Hitachi K, Kiyofuji Y, Nakatani M, Tsuchida K. Myoparr-Associated and -Independent Multiple Roles of Heterogeneous Nuclear Ribonucleoprotein K during Skeletal Muscle Cell Differentiation. International Journal of Molecular Sciences. 2022; 23(1):108. https://doi.org/10.3390/ijms23010108
Chicago/Turabian StyleHitachi, Keisuke, Yuri Kiyofuji, Masashi Nakatani, and Kunihiro Tsuchida. 2022. "Myoparr-Associated and -Independent Multiple Roles of Heterogeneous Nuclear Ribonucleoprotein K during Skeletal Muscle Cell Differentiation" International Journal of Molecular Sciences 23, no. 1: 108. https://doi.org/10.3390/ijms23010108
APA StyleHitachi, K., Kiyofuji, Y., Nakatani, M., & Tsuchida, K. (2022). Myoparr-Associated and -Independent Multiple Roles of Heterogeneous Nuclear Ribonucleoprotein K during Skeletal Muscle Cell Differentiation. International Journal of Molecular Sciences, 23(1), 108. https://doi.org/10.3390/ijms23010108