
Structural and Functional Change in Albino Rat Retina Induced by Various Visible Light Wavelengths



Abstract
:1. Introduction
2. Results
2.1. ERG
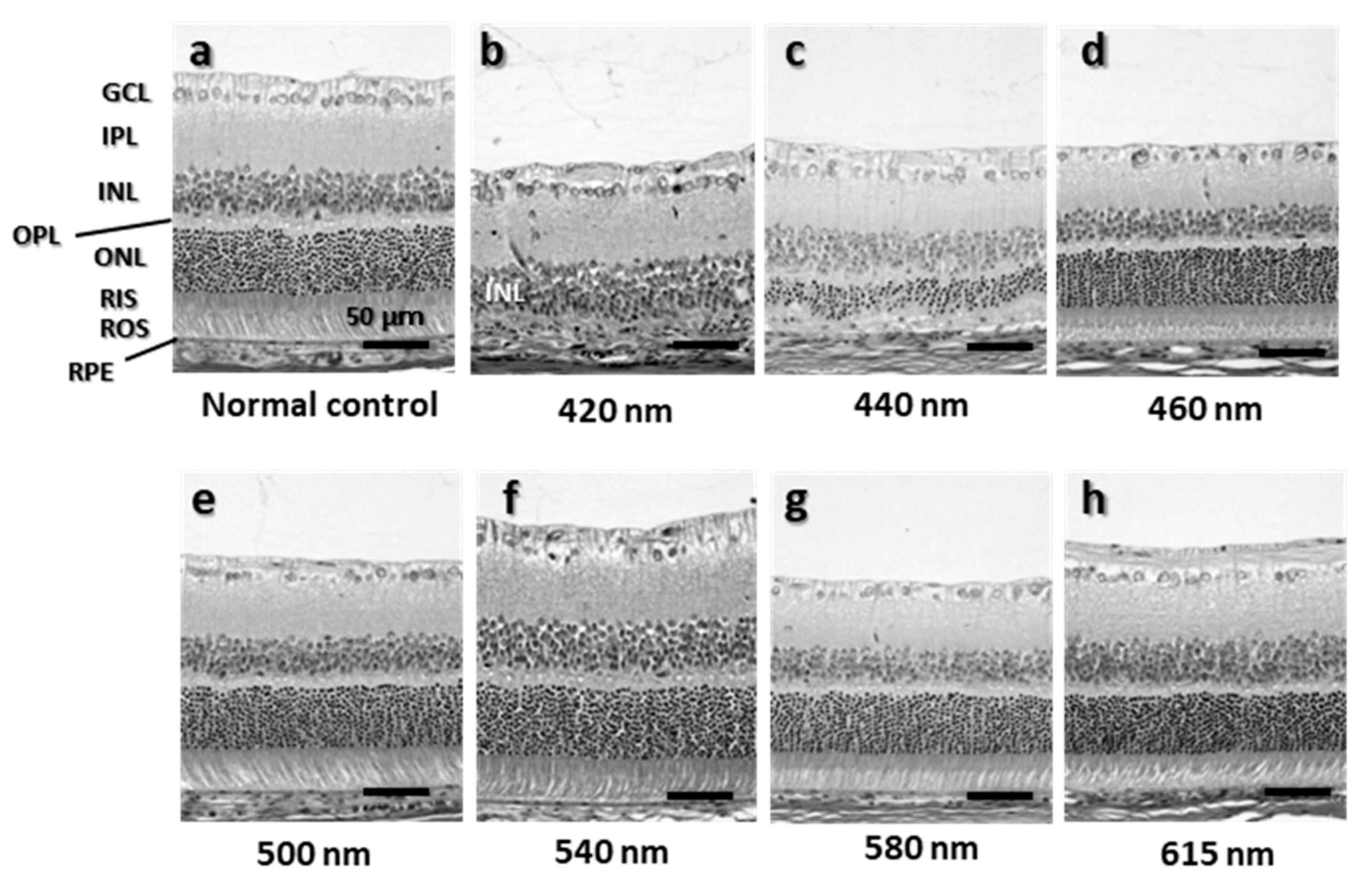
2.2. Morphologic Evaluation by Quantitative Histology
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Light Exposure
4.3. Determination of Duration of Light Exposure
4.4. ERG
4.5. Histology
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noell, W.K.; Walker, V.S.; Kang, B.S.; Berman, S. Retinal damage by light in rats. Investig. Ophthalmol. 1966, 5, 450–473. [Google Scholar]
- Ham, W.T., Jr.; Mueller, H.A.; Sliney, D.H. Retinal sensitivity to damage from short wavelength light. Nature 1976, 260, 153–155. [Google Scholar] [CrossRef]
- Ham, W.T., Jr.; Mueller, H.A.; Ruffolo, J.J., Jr.; Guerry, D., 3rd; Guerry, R.K. Action spectrum for retinal injury from near-ultraviolet radiation in the aphakic monkey. Am. J. Ophthalmol. 1982, 93, 299–306. [Google Scholar] [CrossRef]
- van Norren, D.; Schellekens, P. Blue light hazard in rat. Vision Res. 1990, 30, 1517–1520. [Google Scholar] [CrossRef]
- Rapp, L.M.; Smith, S.C. Morphologic comparisons between rhodopsin-mediated and short-wavelength classes of retinal light damage. Investig. Ophthalmol. Vis. Sci. 1992, 33, 3367–3377. [Google Scholar]
- Gorgels, T.G.; van Norren, D. Ultraviolet and green light cause different types of damage in rat retina. Investig. Ophthalmol. Vis. Sci. 1995, 36, 851–863. [Google Scholar] [CrossRef] [Green Version]
- Tosini, G.; Ferguson, I.; Tsubota, K. Effects of blue light on the circadian system and eye physiology. Mol. Vis. 2016, 22, 61–72. [Google Scholar] [PubMed]
- Ishihara, T.; Kaidzu, S.; Kimura, H.; Koyama, Y.; Matsuoka, Y.; Ohira, A. Protective Effect of Highly Polymeric A-Type Proanthocyanidins from Seed Shells of Japanese Horse Chestnut (Aesculus turbinata BLUME) against Light-Induced Oxidative Damage in Rat Retina. Nutrients 2018, 10, 593. [Google Scholar] [CrossRef] [Green Version]
- Koyama, Y.; Kaidzu, S.; Kim, Y.C.; Matsuoka, Y.; Ishihara, T.; Ohira, A. Suppression of Light-Induced Retinal Degeneration by Quercetin via the AP-1 Pathway in Rats. Antioxidants 2019, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, A.; Grimm, C.; Samardzija, M.; Remé, C.E. Molecular mechanisms of light-induced photoreceptor apoptosis and neuroprotection for retinal degeneration. Prog. Retin. Eye Res. 2005, 24, 275–306. [Google Scholar] [CrossRef]
- American Conference of Governmental Industrial Hygienists (ACGIH). Ultraviolet Radiation: TLV(R) Physical Agents Documentation, Notice of Intended Change; ACGIH: Cincinnati, OH, USA, 2020. [Google Scholar]
- Begum, R.; Powner, M.B.; Hudson, N.; Hogg, C.; Jeffery, G. Treatment with 670 nm light up regulates cytochrome C oxidase expression and reduces inflammation in an age-related macular degeneration model. PLoS ONE 2013, 8, e57828. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Du, Y.; Liu, H.; Tang, J.; Veenstra, A.; Kern, T.S. Photobiomodulation Inhibits Long-term Structural and Functional Lesions of Diabetic Retinopathy. Diabetes 2018, 67, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Natoli, R.; Valter, K.; Barbosa, M.; Dahlstrom, J.; Rutar, M.; Kent, A.; Provis, J. 670nm photobiomodulation as a novel protection against retinopathy of prematurity: Evidence from oxygen induced retinopathy models. PLoS ONE 2013, 8, e72135. [Google Scholar] [CrossRef]
- Sivapathasuntharam, C.; Sivaprasad, S.; Hogg, C.; Jeffery, G. Aging retinal function is improved by near infrared light (670 nm) that is associated with corrected mitochondrial decline. Neurobiol. Aging 2017, 52, 66–70. [Google Scholar] [CrossRef]
- Humphries, M.M.; Rancourt, D.; Farrar, G.J.; Kenna, P.; Hazel, M.; Bush, R.A.; Sieving, P.A.; Sheils, D.M.; McNally, N.; Creighton, P.; et al. Retinopathy induced in mice by targeted disruption of the rhodopsin gene. Nat. Genet. 1997, 15, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Grimm, C.; Wenzel, A.; Hafezi, F.; Yu, S.; Redmond, T.M.; Remé, C.E. Protection of Rpe65-deficient mice identifies rhodopsin as a mediator of light-induced retinal degeneration. Nat. Genet. 2000, 25, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Grimm, C.; Wenzel, A.; Williams, T.; Rol, P.; Hafezi, F.; Remé, C. Rhodopsin-mediated blue-light damage to the rat retina: Effect of photoreversal of bleaching. Investig. Ophthalmol. Vis. Sci. 2001, 42, 497–505. [Google Scholar]
- Hafezi, F.; Steinbach, J.P.; Marti, A.; Munz, K.; Wang, Z.Q.; Wagner, E.F.; Aguzzi, A.; Remé, C. The absence of c-fos prevents light-induced apoptotic cell death of photoreceptors in retinal degeneration in vivo. Nat. Med. 1997, 3, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, A.; Grimm, C.; Marti, A.; Kueng-Hitz, N.; Hafezi, F.; Niemeyer, G.; Remé, C. c-fos controls the "private pathway" of light-induced apoptosis of retinal photoreceptors. J. Neurosci. 2000, 20, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Rapp, L.M.; Tolman, B.L.; Dhindsa, H.S. Separate mechanisms for retinal damage by ultraviolet-A and mid-visible light. Investig. Ophthalmol. Vis. Sci. 1990, 31, 1186–1190. [Google Scholar]
- Krigel, A.; Berdugo, M.; Picard, E.; Levy-Boukris, R.; Jaadane, I.; Jonet, L.; Dernigoghossian, M.; Andrieu-Soler, C.; Torriglia, A.; Behar-Cohen, F. Light-induced retinal damage using different light sources, protocols and rat strains reveals LED phototoxicity. Neuroscience 2016, 339, 296–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapp, L.M.; Williams, T.P. A Parametric Study of Retinal Light Damage in Albino and Pigmented Rats; Plenum Press: New York, NY, USA, 1980. [Google Scholar]
- LaVail, M.M.; Gorrin, G.M.; Repaci, M.A.; Yasumura, D. Light-induced retinal degeneration in albino mice and rats: Strain and species differences. Prog. Clin. Biol. Res. 1987, 247, 439–454. [Google Scholar]
- Organisciak, D.T.; Darrow, R.A.; Barsalou, L.; Darrow, R.M.; Lininger, L.A. Light-induced damage in the retina: Differential effects of dimethylthiourea on photoreceptor survival, apoptosis and DNA oxidation. Photochem. Photobiol. 1999, 70, 261–268. [Google Scholar] [CrossRef]
- Ranchon, I.; LaVail, M.M.; Kotake, Y.; Anderson, R.E. Free radical trap phenyl-N-tert-butylnitrone protects against light damage but does not rescue P23H and S334ter rhodopsin transgenic rats from inherited retinal degeneration. J. Neurosci. 2003, 23, 6050–6057. [Google Scholar] [CrossRef] [PubMed]
- Tanito, M.; Kaidzu, S.; Ohira, A.; Anderson, R.E. Topography of retinal damage in light-exposed albino rats. Exp. Eye Res. 2008, 87, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Rapp, L.M.; Williams, T.P. Rhodopsin content and electroretinographic sensitivity in light-damaged rat retina. Nature 1977, 267, 835–836. [Google Scholar] [CrossRef]
- Anderson, R.E.; Penn, J.S. Environmental light and heredity are associated with adaptive changes in retinal DHA levels that affect retinal function. Lipids 2004, 39, 1121–1124. [Google Scholar] [CrossRef]
- Faktorovich, E.G.; Steinberg, R.H.; Yasumura, D.; Matthes, M.T.; LaVail, M.M. Basic fibroblast growth factor and local injury protect photoreceptors from light damage in the rat. J. Neurosci. 1992, 12, 3554–3567. [Google Scholar] [CrossRef] [Green Version]
- LaVail, M.M.; Unoki, K.; Yasumura, D.; Matthes, M.T.; Yancopoulos, G.D.; Steinberg, R.H. Multiple growth factors, cytokines, and neurotrophins rescue photoreceptors from the damaging effects of constant light. Proc. Natl. Acad. Sci. USA 1992, 89, 11249–11253. [Google Scholar] [CrossRef] [Green Version]
- Stone, J.; Maslim, J.; Valter-Kocsi, K.; Mervin, K.; Bowers, F.; Chu, Y.; Barnett, N.; Provis, J.; Lewis, G.; Fisher, S.K.; et al. Mechanisms of photoreceptor death and survival in mammalian retina. Prog. Retin. Eye Res. 1999, 18, 689–735. [Google Scholar] [CrossRef]
- Ohira, A.; Tanito, M.; Kaidzu, S.; Kondo, T. Glutathione peroxidase induced in rat retinas to counteract photic injury. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1230–1236. [Google Scholar] [CrossRef] [Green Version]
- Organisciak, D.T.; Darrow, R.M.; Barsalou, L.; Kutty, R.K.; Wiggert, B. Circadian-dependent retinal light damage in rats. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3694–3701. [Google Scholar]
- Vaughan, D.K.; Nemke, J.L.; Fliesler, S.J.; Darrow, R.M.; Organisciak, D.T. Evidence for a circadian rhythm of susceptibility to retinal light damage. Photochem. Photobiol. 2002, 75, 547–553. [Google Scholar] [CrossRef]
- Keller, C.; Grimm, C.; Wenzel, A.; Hafezi, F.; Remé, C. Protective effect of halothane anesthesia on retinal light damage: Inhibition of metabolic rhodopsin regeneration. Investig. Ophthalmol. Vis. Sci. 2001, 42, 476–480. [Google Scholar]
- Arango-Gonzalez, B.; Schatz, A.; Bolz, S.; Eslava-Schmalbach, J.; Willmann, G.; Zhour, A.; Zrenner, E.; Fischer, M.D.; Gekeler, F. Effects of combined ketamine/xylazine anesthesia on light induced retinal degeneration in rats. PLoS ONE 2012, 7, e35687. [Google Scholar] [CrossRef]
- Wasowicz, M.; Morice, C.; Ferrari, P.; Callebert, J.; Versaux-Botteri, C. Long-term effects of light damage on the retina of albino and pigmented rats. Investig. Ophthalmol. Vis. Sci. 2002, 43, 813–820. [Google Scholar]
- Polosa, A.; Bessaklia, H.; Lachapelle, P. Strain Differences in Light-Induced Retinopathy. PLoS ONE 2016, 11, e0158082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natoli, R.; Jiao, H.; Barnett, N.L.; Fernando, N.; Valter, K.; Provis, J.M.; Rutar, M. A model of progressive photo-oxidative degeneration and inflammation in the pigmented C57BL/6J mouse retina. Exp. Eye Res. 2016, 147, 114–127. [Google Scholar] [CrossRef] [Green Version]
- Merriam, J.C.; Löfgren, S.; Michael, R.; Söderberg, P.; Dillon, J.; Zheng, L.; Ayala, M. An action spectrum for UV-B radiation and the rat lens. Investig. Ophthalmol. Vis. Sci. 2000, 41, 2642–2647. [Google Scholar]
- Gorgels, T.G.; van Norren, D. Spectral transmittance of the rat lens. Vis. Res. 1992, 32, 1509–1512. [Google Scholar] [CrossRef]
- Tanito, M.; Kaidzu, S.; Anderson, R.E. Protective effects of soft acrylic yellow filter against blue light-induced retinal damage in rats. Exp. Eye Res. 2006, 83, 1493–1504. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Central Wavelength (nm) | Maximum Half Bandwidth (nm) | Transmittance (Cornea + Lens) (%) | Measured Irradiance at Corneal Surface (mW/cm2) | Estimated Radiant Exposure at Corneal Surface (J/cm2) | Estimated Irradiance at Retinas (mW/cm2) | Exposure Time (minutes) | Estimated Radiant Exposure at Retinas (J/cm2) |
|---|---|---|---|---|---|---|---|
| 421 | 8 | 0.626 | 56.0–67.8 | 1086 ± 8 | 35.0–42.4 | 265.0–321.0 | 680 ± 5 |
| 51.0–59.0 | 542 ± 3 | 32.0–36.9 | 153.0–178.0 | 339 ± 2 | |||
| 441 | 12 | 0.671 | 84.6–123.3 | 1011 ± 4 | 56.8–82.8 | 137.0–198.0 | 678 ± 3 |
| 75.6–82.5 | 506 ± 3 | 50.7–55.4 | 103.0–111.0 | 340 ± 2 | |||
| 459 | 11.5 | 0.708 | 114.7–120.5 | 963 ± 5 | 81.2–85.3 | 132.0–140.0 | 680 ± 4 |
| 113.0–126.0 | 480 ± 3 | 80.0–89.6 | 63.5–70.5 | 340 ± 2 | |||
| 501 | 9 | 0.770 | 85.5–129.0 | 891 ± 39 | 65.9–99.3 | 120.0–168.0 | 680 ± 30 |
| 77.0–108.9 | 443 ± 31 | 59.3–83.8 | 71.0–91.0 | 341 ± 24 | |||
| 541 | 14 | 0.796 | 59.1–60.5 | 863 ± 9 | 47.0–48.1 | 235.0–242.0 | 679 ± 7 |
| 58.3–59.4 | 427 ± 5 | 46.4–47.2 | 119.0–123.0 | 340 ± 4 | |||
| 581 | 14.5 | 0.818 | 59.2–170.2 | 833 ± 9 | 48.4–139.2 | 80.0–234.0 | 682 ± 8 |
| 58.7–155.3 | 416 ± 3 | 48.0–127.0 | 44.5–118 | 341 ± 3 | |||
| 615 | 9.5 | 0.841 | 34.1–57.4 | 810 ± 4 | 28.6–48.3 | 235.0–399.0 | 681 ± 3 |
| 37.6–42.6 | 405 ± 5 | 31.6–35.9 | 162.0–180.0 | 341 ± 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaidzu, S.; Okuno, T.; Tanito, M.; Ohira, A. Structural and Functional Change in Albino Rat Retina Induced by Various Visible Light Wavelengths. Int. J. Mol. Sci. 2022, 23, 309. https://doi.org/10.3390/ijms23010309
Kaidzu S, Okuno T, Tanito M, Ohira A. Structural and Functional Change in Albino Rat Retina Induced by Various Visible Light Wavelengths. International Journal of Molecular Sciences. 2022; 23(1):309. https://doi.org/10.3390/ijms23010309
Chicago/Turabian StyleKaidzu, Sachiko, Tsutomu Okuno, Masaki Tanito, and Akihiro Ohira. 2022. "Structural and Functional Change in Albino Rat Retina Induced by Various Visible Light Wavelengths" International Journal of Molecular Sciences 23, no. 1: 309. https://doi.org/10.3390/ijms23010309
APA StyleKaidzu, S., Okuno, T., Tanito, M., & Ohira, A. (2022). Structural and Functional Change in Albino Rat Retina Induced by Various Visible Light Wavelengths. International Journal of Molecular Sciences, 23(1), 309. https://doi.org/10.3390/ijms23010309