
HOXC6-Mediated miR-188-5p Expression Induces Cell Migration through the Inhibition of the Tumor Suppressor FOXN2


Abstract
:1. Introduction
2. Results
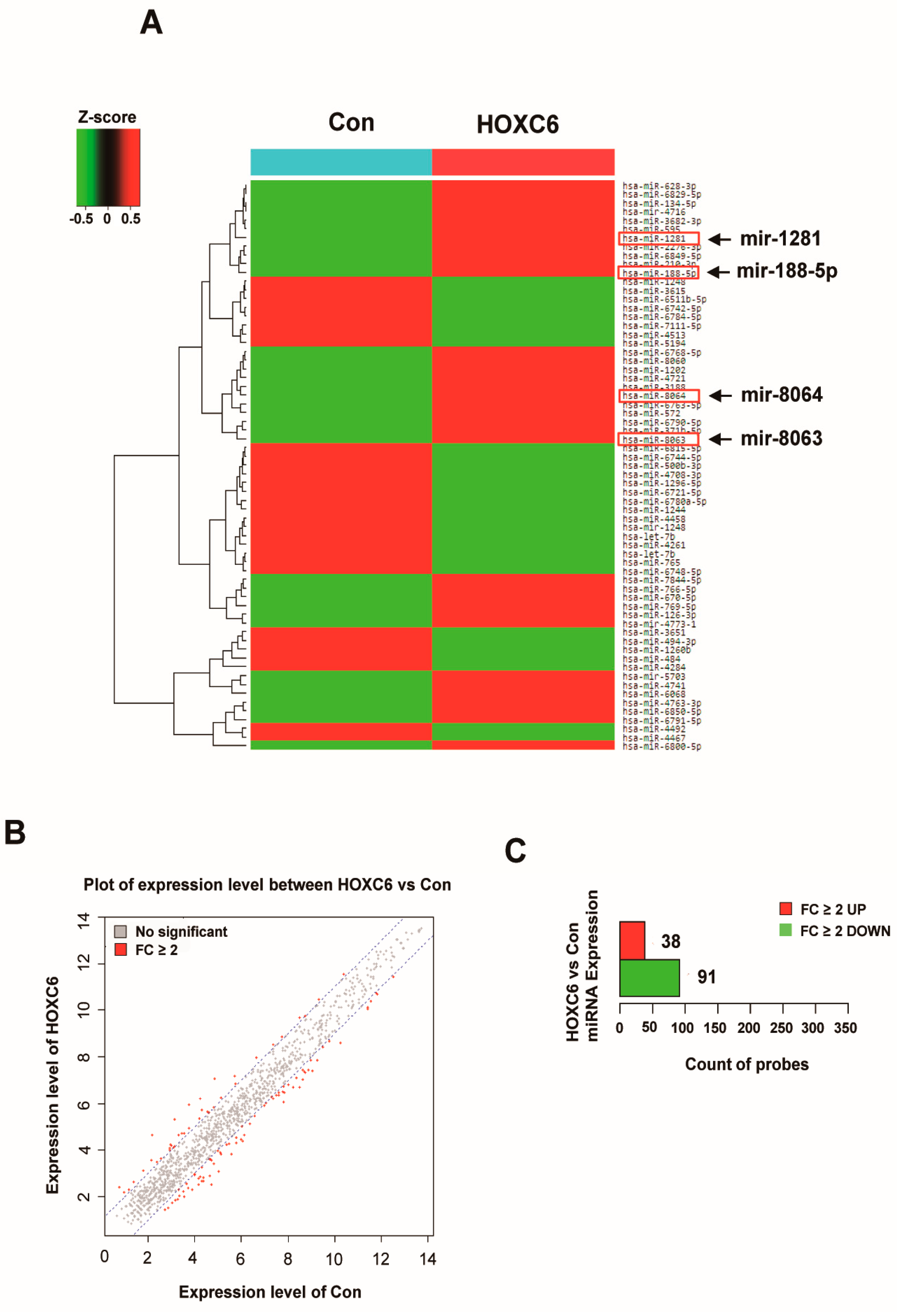
2.1. Differential Expression of MicroRNAs in HOXC6-Expressing Cells
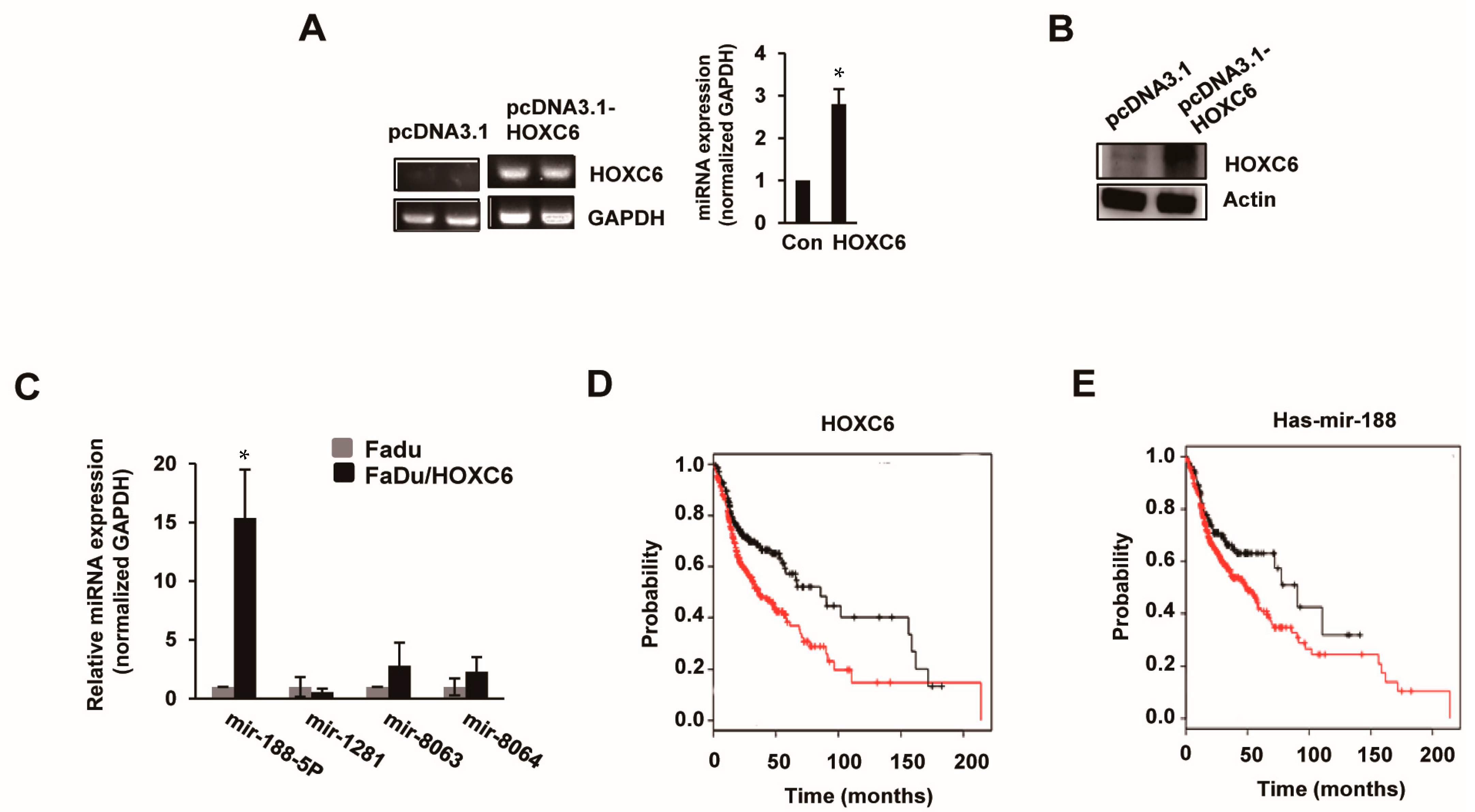
2.2. MicroRNA-188-5p Is Markedly Upregulated in FaDu/HOXC6 Cells
2.3. Prognostic and Predictive Value of miRNA-188-5p in Oral Cancer
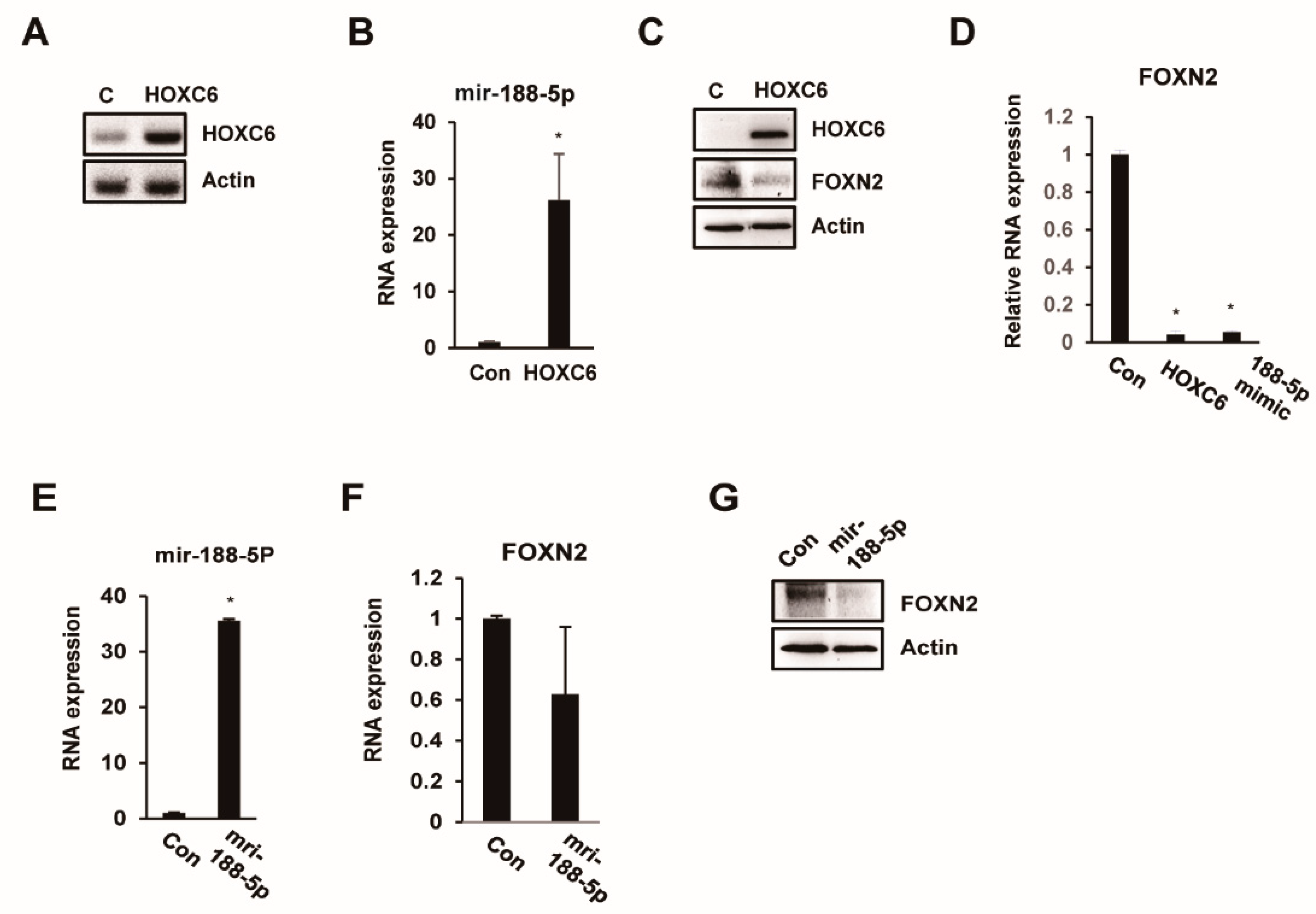
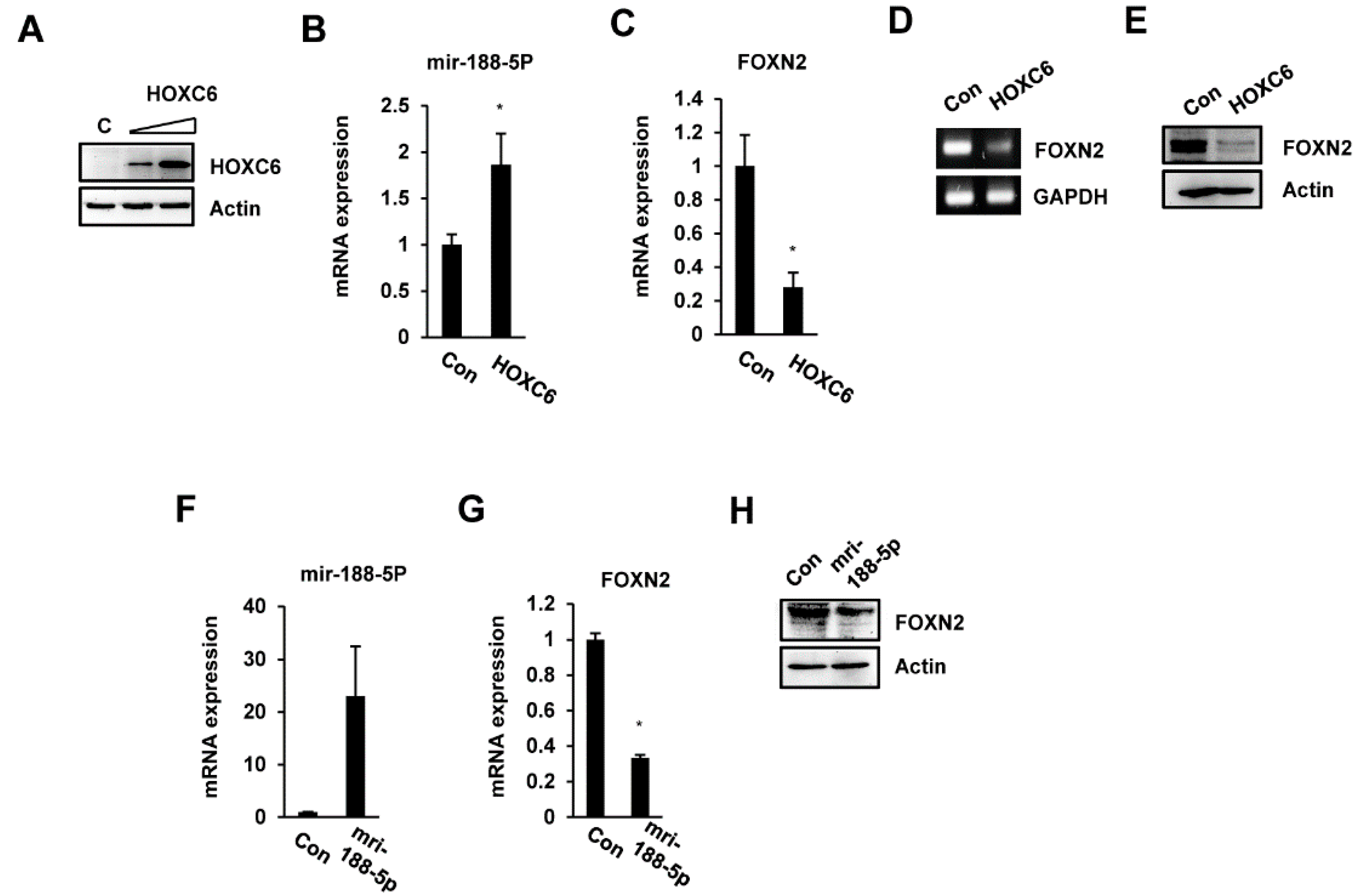
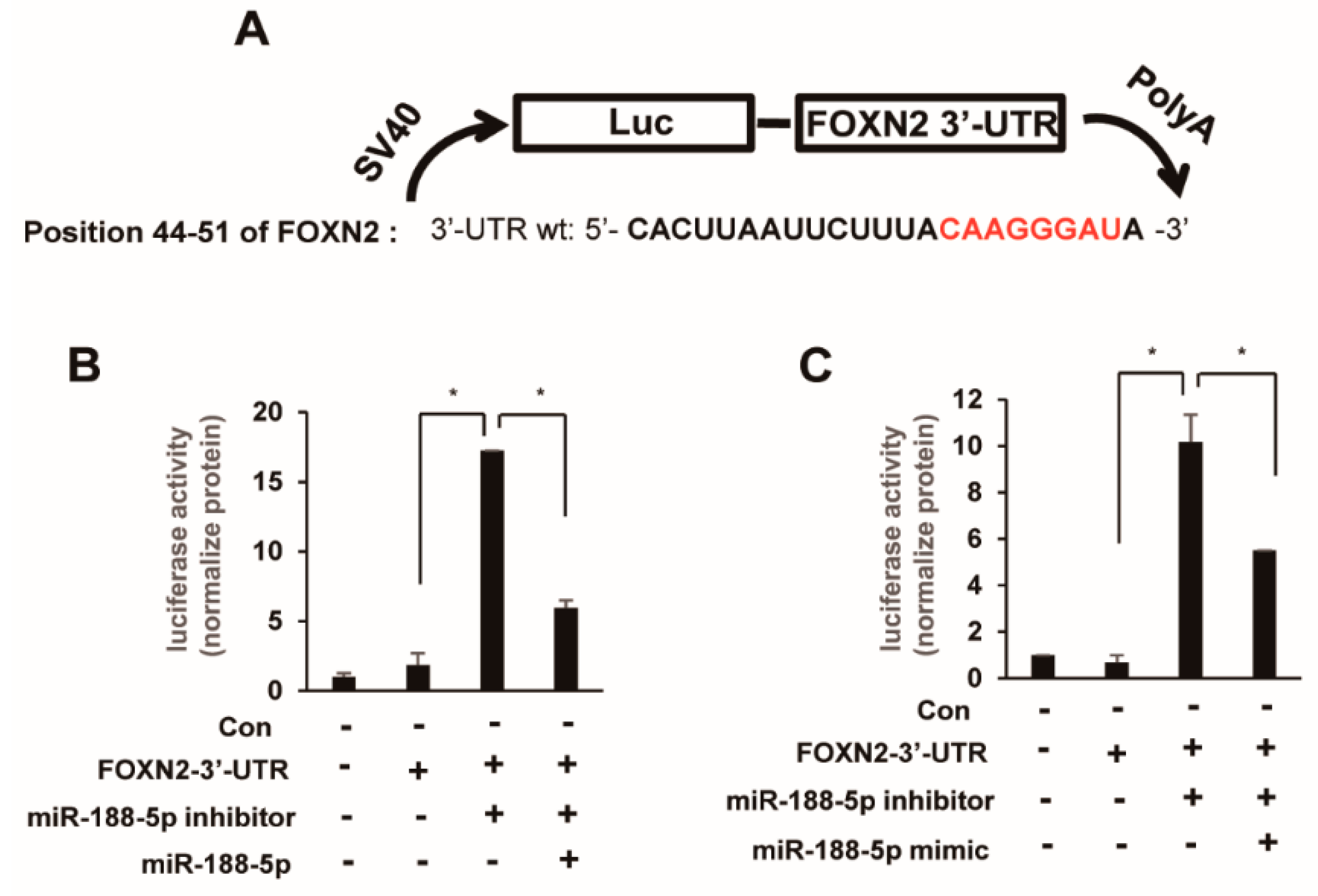
2.4. FOXN2 Is a Target of miR-188-5p
2.5. miR-188-5p Downregulates FOXN2-UTR Luciferase Reporter Activity
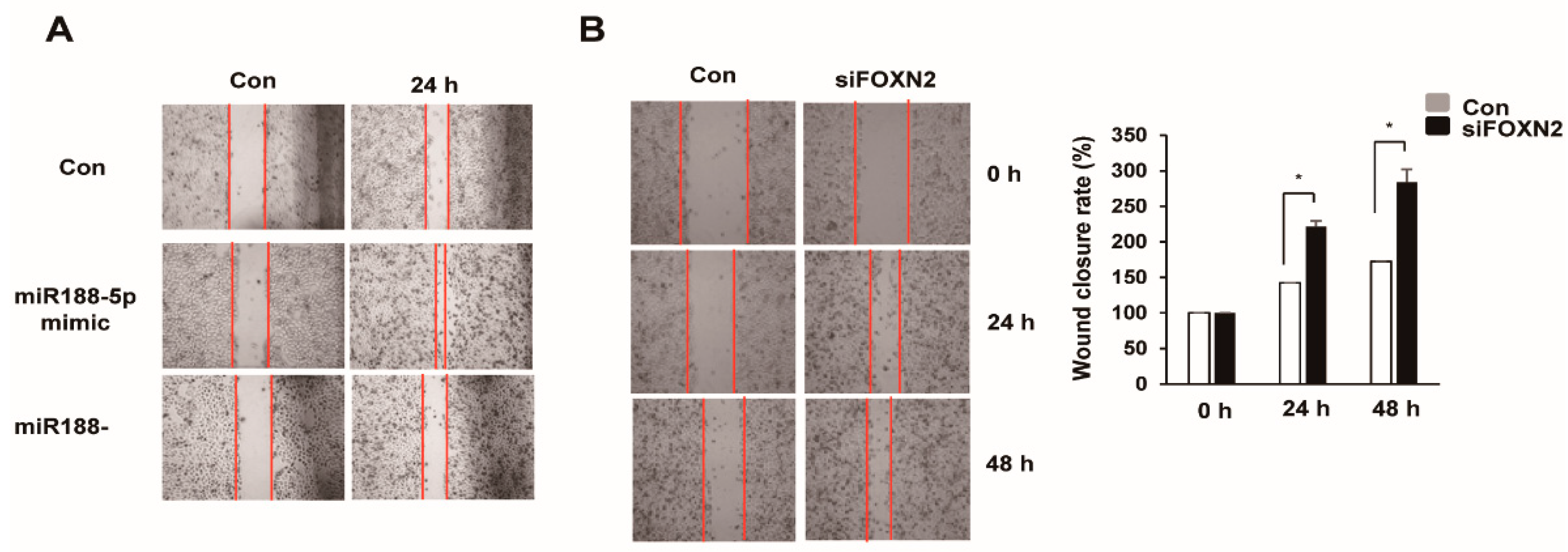
2.6. miR-188-5p Regulates Cell Migration by Inhibiting FOXN2
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. miRNA Microarray and Analysis
4.3. Bioinformatics Analysis
4.4. RT-PCR and Quantitative RT-PCR
4.5. Construction of the pMIR-REPORT-FOXN2 Promoter and Luciferase Reporter Assay
4.6. Transfection of the Hsa-Mir-188-5p Plasmid and Hsa-Mir-188-5p Mimic
4.7. Western Blotting Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ladam, F.; Sagerström, C.G. Hox regulation of transcription—more complex(es). Dev. Dyn. 2014, 243, 4–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, S.; Liu, P.; Young, A.N.; Yin-Goen, Q.; Lim, S.D.; Laycock, N.; Amin, M.B.; Carney, J.K.; Marshall, F.F.; Petros, J.A.; et al. Loss of HOXC6 expression induces apoptosis in prostate cancer cells. Oncogene 2005, 24, 188–198. [Google Scholar] [CrossRef] [Green Version]
- Manfra, M.; Feliciello, I.; Bolognese, A.; Basilicata, M.G.; Pepe, G.; Campiglia, P.; Procino, A. Novel anticancer drug 5h-pyro [3,2-a] phenoxazin-5-one (PPH) regulates lncRNA HOTAIR and HOXC genes in human MCF-7 cells. Bahrain Med. Bull. 2021, 43, 334–341. [Google Scholar]
- Ji, M.; Feng, Q.; He, G.; Yang, L.; Tang, W.; Lao, X.; Zhu, D.; Lin, Q.; Xu, P.; Wei, Y.; et al. Silencing homeobox C6 inhibits colorectal cancer cell proliferation. Oncotarget 2016, 7, 29216–29227. [Google Scholar] [CrossRef] [PubMed]
- Kanai, M.; Hamada, J.-I.; Takada, M.; Asano, T.; Murakawa, K.; Takahashi, Y.; Murai, T.; Tada, M.; Miyamoto, M.; Kondo, S.; et al. Aberrant expressions of HOX genes in colorectal and hepatocellular carcinomas. Oncol. Rep. 2010, 23, 843–851. [Google Scholar]
- Zhang, Q.; Jin, X.S.; Yang, Z.Y.; Wei, M.; Liu, B.Y.; Gu, Q.L. Upregulated Hoxc6 expression is associated with poor survival in gastric cancer patients. Neoplasma 2013, 60, 439–445. [Google Scholar] [CrossRef]
- Cantile, M.; Franco, R.; Schiavo, G.; Procino, A.; Cindolo, L.; Botti, G.; Cillo, C. The HOX genes network in uro-genital cancers: Mechanisms and potential therapeutic implications. Curr. Med. Chem. 2011, 18, 4872–4884. [Google Scholar] [CrossRef]
- Hassan, N.M.M.; Hamada, J.-I.; Murai, T.; Seino, A.; Takahashi, Y.; Tada, M.; Zhang, X.; Kashiwazaki, H.; Yamazaki, Y.; Inoue, N.; et al. Aberrant Expression of HOX Genes in Oral Dysplasia and Squamous Cell Carcinoma Tissues. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2006, 16, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Li, P.D.; Chen, P.; Peng, X.; Ma, C.; Zhang, W.J.; Dai, X.F. HOXC6 predicts invasion and poor survival in hepatocellular car-cinoma by driving epithelial-mesenchymal transition. Aging 2018, 10, 115–130. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.-L.; Chan, T.-C.; Chen, T.-J.; Lee, S.-W.; Lin, L.-C.; Win, K.T. HOXC6 Overexpression Is Associated with Ki-67 Expression and Poor Survival in NPC Patients. J. Cancer 2017, 8, 1647–1654. [Google Scholar] [CrossRef] [Green Version]
- McCabe, C.D.; Spyropoulos, D.D.; Martin, D.; Moreno, C.S. Genome-Wide Analysis of the Homeobox C6 Transcriptional Network in Prostate Cancer. Cancer Res. 2008, 68, 1988–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Ren, C.-C.; Liu, L.; Chen, Y.-N.; Yang, L.; Zhang, X.-A. HOXC6 gene silencing inhibits epithelial-mesenchymal transition and cell viability through the TGF-β/smad signaling pathway in cervical carcinoma cells. Cancer Cell Int. 2018, 18, 204. [Google Scholar] [CrossRef]
- Moon, S.-M.; Kim, S.-A.; Yoon, J.-H.; Ahn, S.-G. HOXC6 Is Deregulated in Human Head and Neck Squamous Cell Carcinoma and Modulates Bcl-2 Expression. J. Biol. Chem. 2012, 287, 35678–35688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.-J.; Moon, S.-M.; Kim, S.-A.; Kang, K.-W.; Yoon, J.-H.; Ahn, S.-G. Transcriptional regulation of MDR-1 by HOXC6 in multidrug-resistant cells. Oncogene 2012, 32, 3339–3349. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.; Croce, C.M. MicroRNA Signatures in Human Cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef]
- Kim, J.; Yao, F.; Xiao, Z.; Sun, Y.; Ma, L. MicroRNAs and metastasis: Small RNAs play big roles. Cancer Metastasis Rev. 2017, 37, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranwal, S.; Alahari, S.K. miRNA control of tumor cell invasion and metastasis. Int. J. Cancer 2009, 126, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.-B.; Dong, B.; Shen, L.-Y.; Yan, W.-P.; Dai, L.; Xiong, H.-C.; Liang, Z.; Kang, X.-Z.; Qin, B.; Chen, K.-N. The survival predictive significance of HOXC6 and HOXC8 in esophageal squamous cell carcinoma. J. Surg. Res. 2014, 188, 442–450. [Google Scholar] [CrossRef]
- Yu, M.; Zhan, J.; Zhang, H. HOX family transcription factors: Related signaling pathways and post-translational modifications in cancer. Cell. Signal 2019, 66, 109469. [Google Scholar] [CrossRef]
- You, X.; Zhou, Z.; Chen, W.; Wei, X.; Zhou, H.; Luo, W. MicroRNA-495 confers inhibitory effects on cancer stem cells in oral squamous cell carcinoma through the HOXC6-mediated TGF-β signaling pathway. Stem Cell Res. Ther. 2020, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yan, X.; Shi, J.; He, Y.; Xu, J.; Lin, L.; Chen, W.; Lin, X.; Lin, X. Aberrantly expressed miR-188-5p promotes gastric cancer metastasis by activating Wnt/β-catenin signaling. BMC Cancer 2019, 19, 505. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Wang, P. MiR-188-5p and MiR-141-3p influence prognosis of bladder cancer and promote bladder cancer synergistically. Pathol. Res. Pract. 2019, 215, 152598. [Google Scholar] [CrossRef]
- Wang, D.; Chen, J.; Ding, Y.; Kong, H.; You, H.; Zhao, Y.; Wei, H.; Liu, Y. miR-188-5p Promotes Tumor Growth by Targeting CD2AP Through PI3K/AKT/mTOR Signaling in Children with Acute Promyelocytic Leukemia. OncoTargets Ther. 2020, 13, 6681–6697. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Cheng, Y.; Han, F.; Chang, Y.; Yang, Y.; Li, X.; Chen, L.; Lu, Y.; Sun, B.; Chen, L. Triptolide Attenuates Renal Tubular Epithelial-mesenchymal Transition Via the MiR-188-5p-mediated PI3K/AKT Pathway in Diabetic Kidney Disease. Int. J. Biol. Sci. 2018, 14, 1545–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Zhang, H.; Yang, F.; Qiu, R.; Zhao, X.; Gong, Z.; Yu, W.; Zhou, B.; Shen, B.; Zhu, W. miR-188-5p suppresses cellular proliferation and migration via IL6ST: A potential noninvasive diagnostic biomarker for breast cancer. J. Cell. Physiol. 2019, 235, 4890–4901. [Google Scholar] [CrossRef]
- Yan, S.; Yue, Y.; Wang, J.; Li, W.; Sun, M.; Gu, C.; Zeng, L. LINC00668 promotes tumorigenesis and progression through sponging miR-188–5p and regulating USP47 in colorectal cancer. Eur. J. Pharmacol. 2019, 858, 172464. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhao, Q.-H.; Liu, J.-Z.; Kong, D.-L. MiRNA-188-5p alleviates the progression of osteosarcoma via target degrading CCNT2. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 29–35. [Google Scholar] [PubMed]
- Liu, X.-H.; Liu, L.-P.; Xu, X.-M.; Hua, M.; Kang, Q.; Li, A.; Huang, L. FOXN2 suppresses the proliferation and invasion of human hepatocellular carcinoma cells. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 731–737. [Google Scholar]
- Ma, J.; Lu, Y.; Zhang, S.; Li, Y.; Huang, J.; Yin, Z.; Ren, J.; Huang, K.; Liu, L.; Yang, K.; et al. beta-Trcp ubiquitin ligase and RSK2 kinase-mediated degradation of FOXN2 promotes tumorigenesis and radioresistance in lung cancer. Cell Death Differ. 2018, 25, 1473–1485. [Google Scholar] [CrossRef] [PubMed]
- Myatt, S.S.; Lam, E. The emerging roles of forkhead box (Fox) proteins in cancer. Nat. Rev. Cancer 2007, 7, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Pommerenke, C.; Meyer, C.; Kaufmann, M.; MacLeod, R.A.F.; Drexler, H.G. Identification of a tumor suppressor network in T-cell leukemia. Leuk. Lymphoma 2017, 58, 2196–2207. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.; Duan, M. FOXN2 is downregulated in breast cancer and regulates migration, invasion, and epithelial-mesenchymal transition through regulation of SLUG. Cancer Manag. Res. 2019, 11, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | miRbase | Chromosome | Fold Change | Fold Change Direction |
|---|---|---|---|---|
| hsa-miR-1281 | MIMAT0005939 | 22q13.2 | 5.48 | up |
| hsa-miR-8063 | MIMAT0030990 | 15q14 | 4.68 | up |
| hsa-miR-188-5p | MIMAT0000457 | Xp11.23 | 4.55 | up |
| hsa-miR-8064 | MIMAT0030991 | 3p21.1 | 4.5 | up |
| hsa-miR-3188 | MIMAT0015070 | 19p13.11 | 4 | up |
| hsa-miR-6790-5p | MIMAT0027480 | 19p13.3 | 2.76 | up |
| hsa-miR-670-5p | MIMAT0010357 | 11p11.2 | 2.71 | up |
| hsa-miR-1202 | MIMAT0005865 | 6q25.3 | 2.63 | up |
| hsa-miR-6849-5p | MIMAT0027598 | 8q24.3 | 2.57 | up |
| hsa-miR-6068 | MIMAT0023693 | 1p31.3 | 2.52 | up |
| hsa-miR-769-5p | MIMAT0003886 | 19q13.32 | 2.47 | up |
| hsa-miR-2276-3p | MIMAT0011775 | 13q12.12 | 2.41 | up |
| hsa-miR-134-5p | MIMAT0000447 | 14q32.31 | 2.37 | up |
| hsa-miR-126-3p | MIMAT0000445 | 9q34.3 | 2.35 | up |
| hsa-miR-6721-5p | MIMAT0025852 | 6p21.32 | −4.65 | down |
| hsa-miR-4284 | MIMAT0016915 | 7q11.23 | −3.49 | down |
| hsa-miR-1296-5p | MIMAT0005794 | 10q21.3 | −3.36 | down |
| hsa-miR-1260b | MIMAT0015041 | 11q21 | −3.32 | down |
| hsa-miR-5194 | MIMAT0021125 | 8q24.21 | −3.31 | down |
| hsa-miR-500b-3p | MIMAT0027032 | Xp11.23 | −3.24 | down |
| hsa-miR-4708-3p | MIMAT0019810 | 14q23.3 | −3.16 | down |
| hsa-miR-4458 | MIMAT0018980 | 5p15.31 | −3.01 | down |
| hsa-miR-7111-5p | MIMAT0028119 | 6p21.31 | −2.77 | down |
| hsa-miR-6815-5p | MIMAT0027530 | 21q22.3 | −2.66 | down |
| hsa-let-7b | MI0000063 | 22q13.31 | −2.63 | down |
| hsa-miR-6744-5p | MIMAT0027389 | 11p15.5 | −2.62 | down |
| hsa-mir-1248 | MI0006383 | 3q27.3 | −2.61 | down |
| hsa-miR-3615 | MIMAT0017994 | 17q25.1 | −2.5 | down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, S.; Kim, S.-A.; Ahn, S.-G. HOXC6-Mediated miR-188-5p Expression Induces Cell Migration through the Inhibition of the Tumor Suppressor FOXN2. Int. J. Mol. Sci. 2022, 23, 9. https://doi.org/10.3390/ijms23010009
Jeong S, Kim S-A, Ahn S-G. HOXC6-Mediated miR-188-5p Expression Induces Cell Migration through the Inhibition of the Tumor Suppressor FOXN2. International Journal of Molecular Sciences. 2022; 23(1):9. https://doi.org/10.3390/ijms23010009
Chicago/Turabian StyleJeong, Seho, Soo-A Kim, and Sang-Gun Ahn. 2022. "HOXC6-Mediated miR-188-5p Expression Induces Cell Migration through the Inhibition of the Tumor Suppressor FOXN2" International Journal of Molecular Sciences 23, no. 1: 9. https://doi.org/10.3390/ijms23010009
APA StyleJeong, S., Kim, S. -A., & Ahn, S. -G. (2022). HOXC6-Mediated miR-188-5p Expression Induces Cell Migration through the Inhibition of the Tumor Suppressor FOXN2. International Journal of Molecular Sciences, 23(1), 9. https://doi.org/10.3390/ijms23010009