
GCR1 Positively Regulates UV-B- and Ethylene-Induced Stomatal Closure via Activating GPA1-Dependent ROS and NO Production

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
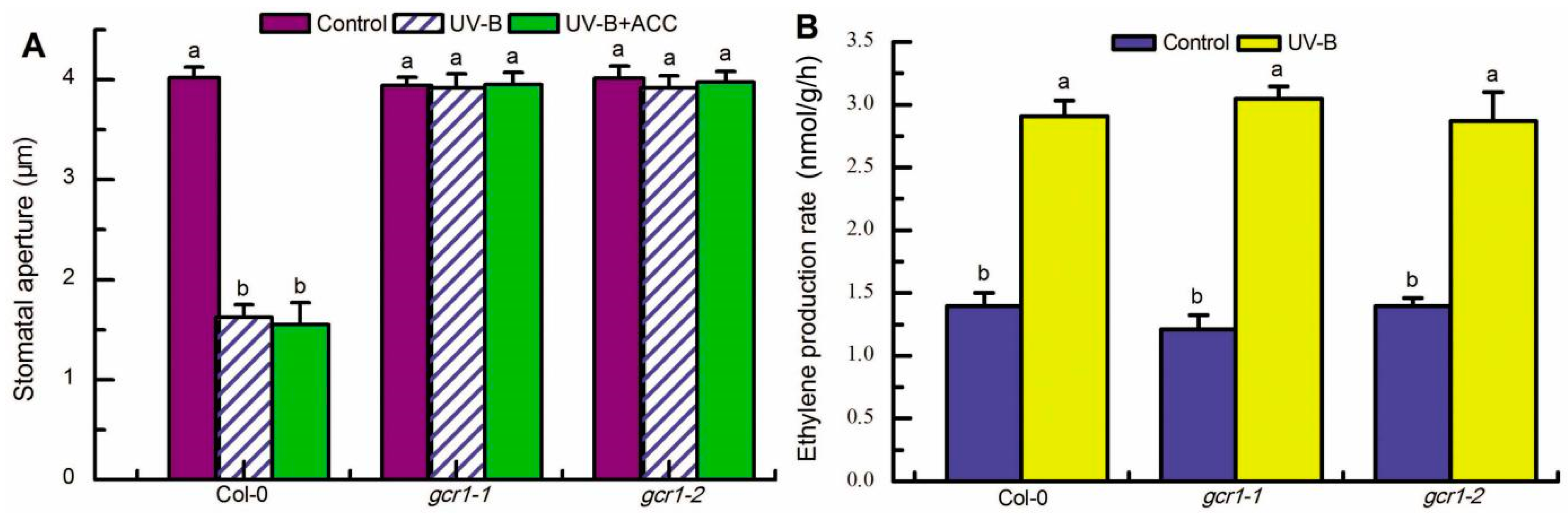
2.1. GCR1 Positively Regulates UV-B- and Ethylene-Induced Stomatal Closure
2.2. GCR1 Mediates UV-B- and Ethylene-Induced Stomatal Closure via Activating Gα Subunit GPA1
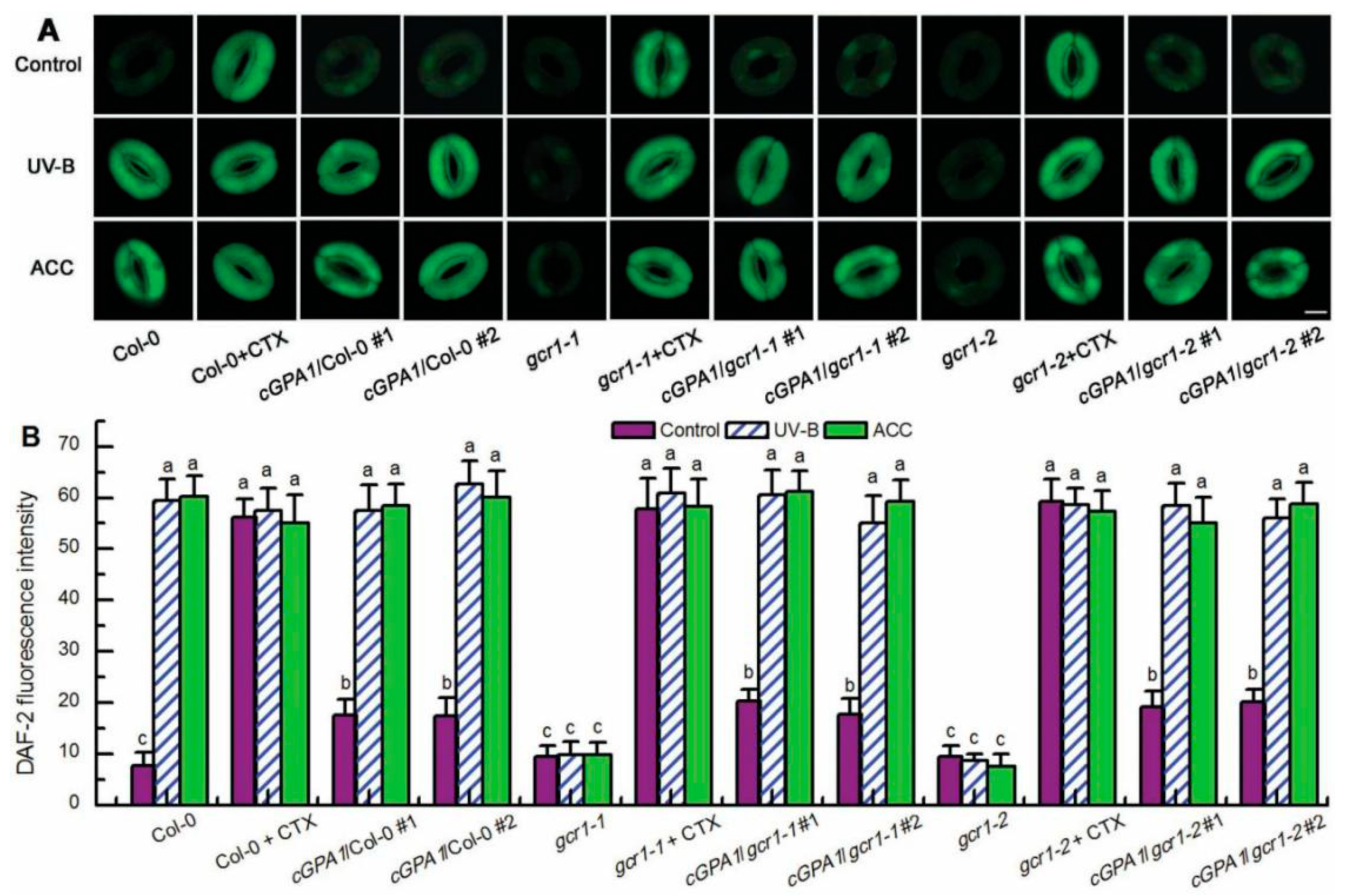
2.3. GCR1 Mediates UV-B- and Ethylene-Induced Stomatal Closure by Activating GPA1-Dependent ROS Production in Guard Cells
2.4. GCR1 Mediates UV-B- and Ethylene-Induced Stomatal Closure by Activating GPA1-Dependent NO Production in Guard Cells
2.5. Ethylene Functions Upstream of GCR1 to Transduce UV-B Signaling in Guard Cells
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. UV-B Treatments
4.3. Stomatal Bioassays
4.4. Measurement of Leaf Water Loss Rate
4.5. Measurement of ROS and NO Levels in Guard Cells
4.6. Generation of Transgenic Plants Overexpressing cGPA1
4.7. RNA Extraction and qPCR
4.8. Ethylene Measurement
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oldham, W.M.; Hamm, H.E. Heterotrimeric G protein activation by G-protein-coupled receptors. Nat. Rev. Mol. Cell Biol. 2008, 9, 60–71. [Google Scholar] [CrossRef]
- Josefsson, L.-G.; Rask, L. Cloning of a putative G-protein-coupled receptor from Arabidopsis thaliana. Eur. J. Biochem. 1997, 249, 415–420. [Google Scholar] [CrossRef] [Green Version]
- Plakidou-Dymock, S.; Dymock, D.; Hooley, R. A higher plant seven-transmembrane receptor that influences sensitivity to cytokinins. Curr. Biol. 1998, 8, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Moriyama, E.N.; Strope, P.K.; Opiyo, S.O.; Chen, Z.; Jones, A.M. Mining the Arabidopsis thaliana genome for highly-divergent seven transmembrane receptors. Genome Biol. 2006, 7, R96. [Google Scholar] [CrossRef] [Green Version]
- Gookin, T.E.; Kim, J.; Assmann, S.M. Whole proteome identification of plant candidate G-protein coupled receptors in Arabidopsis, rice, and poplar: Computational prediction and in-vivo protein coupling. Genome Biol. 2008, 9, R120. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.; Nelson, D.C.; Assmann, S.M. Two novel GPCR-type G proteins are abscisic acid receptors in Arabidopsis. Cell 2009, 136, 136–148. [Google Scholar] [CrossRef] [Green Version]
- Taddese, B.; Upton, G.J.; Bailey, G.R.; Jordan, S.R.; Abdulla, N.Y.; Reeves, P.J.; Reynolds, C.A. Do plants contain G protein-coupled receptors? Plant Physiol. 2014, 164, 287–307. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.; Assmann, S.M. The Arabidopsis putative G protein-coupled receptor GCR1 interacts with the G protein alpha subunit GPA1 and regulates abscisic acid signaling. Plant Cell 2004, 16, 1616–1632. [Google Scholar] [CrossRef] [Green Version]
- Urano, D.; Jones, A.M. “Round up the usual suspects”: A comment on nonexistent plant G protein-coupled receptors. Plant Physiol. 2013, 161, 1097–1102. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Yanofsky, M.F.; Meyerowitz, E.M. Molecular cloning and characterization of GPA1, a G protein alpha subunit gene from Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1990, 87, 3821–3825. [Google Scholar] [CrossRef] [Green Version]
- Heo, J.B.; Sung, S.; Assmann, S.M. Ca2+-dependent GTPase, extra-large G protein 2 (XLG2), promotes activation of DNA-binding protein related to vernalization 1 (RTV1), leading to activation of floral integrator genes and early flowering in Arabidopsis. J. Biol. Chem. 2012, 287, 8242–8253. [Google Scholar] [CrossRef] [Green Version]
- Weiss, C.A.; Garnaat, C.W.; Mukai, K.; Hu, Y.; Ma, H. Isolation of cDNAs encoding guanine nucleotide-binding protein betasubunit homologues from maize (ZGB1) and Arabidopsis (AGB1). Proc. Natl. Acad. Sci. USA 1994, 91, 9554–9558. [Google Scholar] [CrossRef] [Green Version]
- Mason, M.G.; Botella, J.R. Completing the heterotrimer: Isolation and characterization of an Arabidopsis thaliana G protein γ-subunit cDNA. Proc. Natl. Acad. Sci. USA 2000, 97, 14784–14788. [Google Scholar] [CrossRef] [Green Version]
- Mason, M.G.; Botella, J.R. Isolation of a novel G-protein γ-subunit from Arabidopsis thaliana and its interaction with Gβ. Biochim. Biophys. Acta 2001, 1520, 147–153. [Google Scholar] [CrossRef]
- Chen, J.G.; Willard, F.S.; Huang, J.; Liang, J.; Chasse, S.A.; Jones, A.M.; Siderovski, D.P. A seven-transmembrane RGS protein that modulates plant cell proliferation. Science 2003, 30, 1728–1731. [Google Scholar] [CrossRef] [Green Version]
- Wess, J. Molecular basis of receptor/G-protein-coupling selectivity. Pharmacol. Ther. 1998, 80, 231–264. [Google Scholar] [CrossRef]
- Ullah, H.; Chen, J.G.; Young, J.C.; Im, K.H.; Sussman, M.R.; Jones, A.M. Modulation of cell proliferation by heterotrimeric G protein in Arabidopsis. Science 2001, 292, 2066–2069. [Google Scholar] [CrossRef] [Green Version]
- Ullah, H.; Chen, J.G.; Wang, S.; Jones, A.M. Role of a heterotrimeric G protein in regulation of Arabidopsis seed germination. Plant Physiol. 2002, 129, 897–907. [Google Scholar] [CrossRef] [Green Version]
- Colucci, G.; Apone, F.; Alyeshmerni, N.; Chalmers, D.; Chrispeels, M.J. GCR1, the putative Arabidopsis G protein-coupled receptor gene is cell cycle-regulated, and its overexpression abolishes seed dormancy and shortens time to flowering. Proc. Natl. Acad. Sci. USA 2002, 99, 4736–4741. [Google Scholar] [CrossRef] [Green Version]
- Apone, F.; Alyeshmerni, N.; Wiens, K.; Chalmers, D.; Chrispeels, M.J.; Colucci, G. The G-protein-coupled receptor GCR1 regulates DNA synthesis through activation of phosphatidylinositol specific phospholipase C. Plant Physiol. 2003, 133, 571–579. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.C.; Jones, A.M.; Temple, B.R.S.; Dohlman, H.G. Differences in intradomain and interdomain motion confer distinct activation properties to structurally similar Gα proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 7275–7279. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.C.; Temple, B.R.S.; Jones, A.M.; Dohlman, H.G. Functional reconstitution of an atypical G Protein heterotrimer and regulator of G Protein signaling protein (RGS1) from Arabidopsis thaliana. J. Biol. Chem. 2011, 286, 13143–13150. [Google Scholar] [CrossRef] [Green Version]
- Bommert, P.; Il Je, B.; Goldshmidt, A.; Jackson, D. The maize Galpha gene COMPACT PLANT2 functions in CLAVATA signalling to control shoot meristem size. Nature 2013, 502, 555–558. [Google Scholar] [CrossRef]
- Choudhury, S.R.; Pandey, S. Specific subunits of heterotrimeric G proteins play important roles during nodulation in Soybean. Plant Physiol. 2013, 162, 522–533. [Google Scholar] [CrossRef] [Green Version]
- Aranda-Sicilia, M.N.; Trusov, Y.; Maruta, N.; Chakravorty, D.; Zhang, Y.L.; Botella, J.R. Heterotrimeric G proteins interact with defense-related receptor-like kinases in Arabidopsis. J. Plant Physiol. 2015, 188, 44–48. [Google Scholar] [CrossRef]
- Liang, X.; Ding, P.; Lian, K.; Wang, J.; Ma, M.; Li, L.; Li, L.; Li, M.; Zhang, X.; Chen, S.; et al. Arabidopsis heterotrimeric G proteins regulate immunity by directly coupling to the FLS2 receptor. eLife 2016, 5, e13568. [Google Scholar] [CrossRef]
- Liang, X.; Ma, M.; Zhou, Z.; Wang, J.; Yang, X.; Rao, S.; Bi, G.; Li, L.; Zhang, X.; Chai, J.; et al. Ligand-triggered de-repression of Arabidopsis heterotrimeric G proteins coupled to immune receptor kinases. Cell Res. 2018, 28, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Tunc-Ozdemir, M.; Urano, D.; Jia, H.; Werth, E.G.; Mowrey, D.D.; Hicks, L.M.; Dokholyan, N.V.; Torres, M.P.; Jones, A.M. Tyrosine phosphorylation switching of a G protein. J. Biol. Chem. 2018, 293, 4752–4766. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Chen, L.; Li, S.; Zhang, Y.; Xu, R.; Liu, Z.; Liu, W.; Kong, J.; Huang, X.; Wang, Y.; et al. BRI1 and BAK1 interact with G proteins and regulate sugar-responsive growth and development in Arabidopsis. Nat. Commun. 2018, 9, 1522. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Chakravorty, D.; Assmann, S.M. The G protein β-subunit, AGB1, interacts with FERONIA in RALF1-regulated stomatal movement. Plant Physiol. 2018, 176, 2426–2440. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Gong, B.-Q.; Yao, X.; Huang, X.; Li, J.-F. BAK1-mediated phosphorylation of canonical G protein alpha during flagellin signaling in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 690–701. [Google Scholar] [CrossRef]
- Stateczny, D.; Oppenheimer, J.; Bommert, P. G protein signaling in plants: Minus times minus equals plus. Curr. Opin. Plant Biol. 2016, 34, 127–135. [Google Scholar] [CrossRef]
- Trusov, Y.; Botella, J.R. Plant G-proteins come of age: Breaking the bond with animal models. Front. Chem. 2016, 4, 24. [Google Scholar] [CrossRef] [Green Version]
- Coursol, S.; Fan, L.M.; Le Stunff, H.; Spiegel, S.; Gilroy, S.; Assmann, S.M. Sphingolipid signalling in Arabidopsis guard cells involves heterotrimeric G proteins. Nature 2003, 423, 651–654. [Google Scholar] [CrossRef]
- Coursol, S.; Le Stunff, H.; Lynch, D.V.; Gilroy, S.; Assmann, S.M.; Spiegel, S. Arabidopsis sphingosine kinase and the effects of phytosphingosine-1-phosphate on stomatal aperture. Plant Physiol. 2005, 137, 724–737. [Google Scholar] [CrossRef] [Green Version]
- Mishra, G.; Zhang, W.; Deng, F.; Zhao, J.; Wang, X. A bifurcating pathway directs abscisic acid effects on stomatal closure and opening in Arabidopsis. Science 2006, 312, 264–266. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.L.; Huang, R.; Xiao, Y.M.; Lü, P.; Chen, J.; Wang, X.C. Extracellular calmodulin-induced stomatal closure is mediated by heterotrimeric G protein and H2O2. Plant Physiol. 2004, 136, 4096–4103. [Google Scholar] [CrossRef] [Green Version]
- Li, J.H.; Liu, Y.Q.; Lü, P.; Lin, H.F.; Bai, Y.; Wang, X.C.; Chen, Y.L. A signaling pathway linking nitric oxide production to heterotrimeric G protein and hydrogen peroxide regulates extracellular calmodulin induction of stomatal closure in Arabidopsis. Plant Physiol. 2009, 150, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Hao, L.H.; Wang, W.X.; Chen, C.; Wang, Y.F.; Liu, T.; Li, X.; Shang, Z.L. Extracellular ATP promotes stomatal opening of Arabidopsis thaliana through heterotrimeric G protein a subunit and reactive oxygen species. Mol. Plant 2012, 5, 852–864. [Google Scholar] [CrossRef] [Green Version]
- He, J.M.; Ma, X.G.; Zhang, Y.; Sun, T.F.; Xu, F.F.; Chen, Y.P.; Liu, X.; Yue, M. Role and interrelationship of Ga protein, hydrogen peroxide, and nitric oxide in ultraviolet B-induced stomatal closure in Arabidopsis leaves. Plant Physiol. 2013, 161, 1570–1583. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.M.; Cai, H.L.; Lei, X.; Zhou, X.; Yue, M.; He, J.M. Heterotrimeric G protein mediates ethylene-induced stomatal closure via hydrogen peroxide synthesis in Arabidopsis. Plant J. 2015, 82, 138–150. [Google Scholar] [CrossRef]
- Shi, C.; Qi, C.; Ren, H.; Huang, A.; Hei, S.; She, X. Ethylene mediates brassinosteroid-induced stomatal closure via Galpha protein-activated hydrogen peroxide and nitric oxide production in Arabidopsis. Plant J. 2015, 82, 280–301. [Google Scholar] [CrossRef]
- Zhang, W.; He, S.Y.; Assmann, S.M. The plant innate immunity response in stomatal guard cells invokes G-protein-dependent ion channel regulation. Plant J. 2008, 56, 984–996. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Q.; Ullah, H.; Jones, A.M.; Assmann, S.M. G protein regulation of ion channels and abscisic acid signaling in Arabidopsis guard cells. Science 2001, 292, 2070–2072. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Jeon, B.W.; Assmann, S.M. Heterotrimeric G-protein regulation of ROS signalling and calcium currents in Arabidopsis guard cells. J. Exp. Bot. 2011, 62, 2371–2379. [Google Scholar] [CrossRef] [Green Version]
- Temple, B.R.S.; Jones, A.M. The plant heterotrimeric G-protein complex. Annu. Rev. Plant Biol. 2007, 58, 249–266. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, H.; Matsui, M.; Deng, X.W. Overexpression of the heterotrimeric G-protein α-subunit enhances phytochrome-mediated inhibition of hypocotyl elongation in Arabidopsis. Plant Cell 2001, 13, 1639–1652. [Google Scholar]
- Zhang, T.Y.; Li, Z.Q.; Zhao, Y.D.; Shen, W.J.; Chen, M.S.; Gao, H.Q.; Ge, X.M.; Wang, H.Q.; Li, X.; He, J.M. Ethylene-induced stomatal closure is mediated via MKK1/3-MPK3/6 cascade to EIN2 and EIN3. J. Integr. Plant Biol. 2021, 63, 1324–1340. [Google Scholar] [CrossRef]
- Ge, X.M.; Hu, X.; Zhang, J.; Huang, Q.M.; Gao, Y.; Li, Z.Q.; Li, S.; He, J.M. UV resistance LOCUS8 mediates ultraviolet-B-induced stomatal closure in an ethylene-dependent manner. Plant Sci. 2020, 301, e110679. [Google Scholar] [CrossRef]
- Urano, D.; Chen, J.-G.; Botella, J.R.; Jones, A.M. Heterotrimeric G protein signalling in the plant kingdom. Open Biol 2013, 3, 120186. [Google Scholar] [CrossRef] [Green Version]
- Desikan, R.; Last, K.; Harrett-Williams, R.; Tagliavia, C.; Harter, K.; Hooley, R.; Hancock, J.T.; Neill, S.J. Ethylene-induced stomatal closure in Arabidopsis occurs via AtrbohF-mediated hydrogen peroxide synthesis. Plant J. 2006, 47, 907–916. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sano, T.; Tamaoki, M.; Nakajima, N.; Kondo, N.; Hasezawa, S. Ethylene inhibits abscisic acid-induced stomatal closure in Arabidopsis. Plant Physiol. 2005, 138, 2337–2343. [Google Scholar] [CrossRef] [Green Version]
- Aharon, G.; Gelli, A.; Snedden, W.A.; Blumwald, E. Activation of a plant plasma membrane Ca2+ channel by TGα1, a heterotrimeric G protein α-subunit homologue. FEBS Lett. 1998, 424, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Li, F.C.; Wang, J.; Wu, M.M.; Fan, C.M.; Li, X.; He, J.M. Mitogen-activated protein kinase phosphatases affect UV-B-induced stomatal closure via controlling NO in guard cells. Plant Physiol. 2017, 173, 760–770. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Fu, Q.; Zhao, F.-X.; Wu, Y.-Q.; Zhang, T.-Y.; Li, Z.-Q.; He, J.-M. GCR1 Positively Regulates UV-B- and Ethylene-Induced Stomatal Closure via Activating GPA1-Dependent ROS and NO Production. Int. J. Mol. Sci. 2022, 23, 5512. https://doi.org/10.3390/ijms23105512
Li X, Fu Q, Zhao F-X, Wu Y-Q, Zhang T-Y, Li Z-Q, He J-M. GCR1 Positively Regulates UV-B- and Ethylene-Induced Stomatal Closure via Activating GPA1-Dependent ROS and NO Production. International Journal of Molecular Sciences. 2022; 23(10):5512. https://doi.org/10.3390/ijms23105512
Chicago/Turabian StyleLi, Xue, Qi Fu, Fu-Xing Zhao, Yi-Qing Wu, Teng-Yue Zhang, Zhong-Qi Li, and Jun-Min He. 2022. "GCR1 Positively Regulates UV-B- and Ethylene-Induced Stomatal Closure via Activating GPA1-Dependent ROS and NO Production" International Journal of Molecular Sciences 23, no. 10: 5512. https://doi.org/10.3390/ijms23105512
APA StyleLi, X., Fu, Q., Zhao, F.-X., Wu, Y.-Q., Zhang, T.-Y., Li, Z.-Q., & He, J.-M. (2022). GCR1 Positively Regulates UV-B- and Ethylene-Induced Stomatal Closure via Activating GPA1-Dependent ROS and NO Production. International Journal of Molecular Sciences, 23(10), 5512. https://doi.org/10.3390/ijms23105512