
Behavioral Evidence for a Tau and HIV-gp120 Interaction




{kind=link}
{kind=link}
Abstract
:1. Introduction
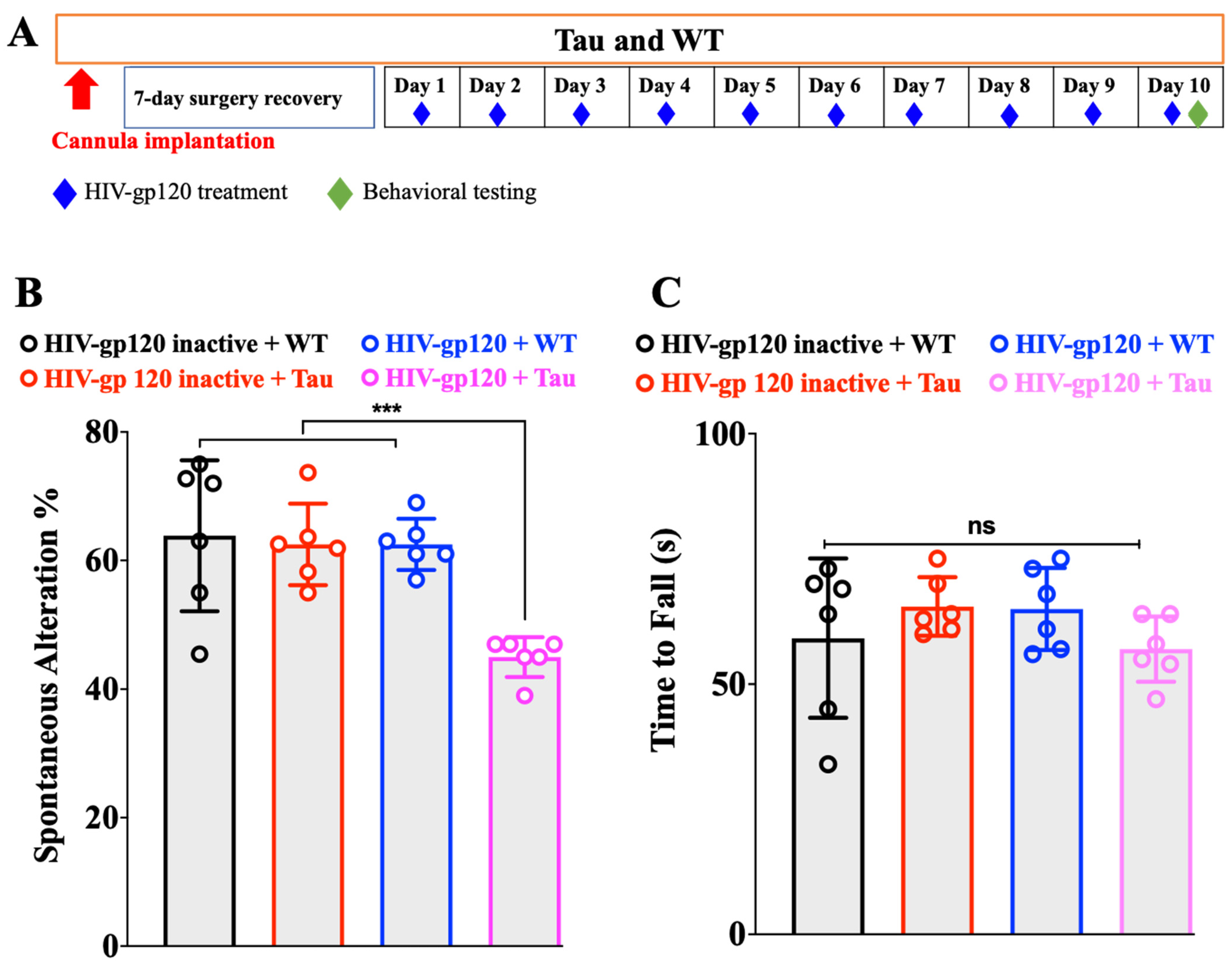
2. Results
2.1. HIV-gp120 Promotes the Onset of Cognitive Deficit of Tau Mice
2.2. Effect of Mitochondrial Division Inhibitor 1 (Mdivi1) on the Cognitive Interaction between HIV-gp120 and Tau Mice
3. Discussion
4. Methods
4.1. Animals
4.2. Behavior
4.3. Cannula Implantation
4.4. Statistical and Histologic Analysis
4.5. Drugs
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- The Global Impact of Dementia 2013–2050. 2013. Available online: https://www.alz.co.uk/research/GlobalImpactDementia2013.pdf (accessed on 1 April 2022).
- Wimo, A.; Guerchet, M.; Ali, G.C.; Wu, Y.T.; Prina, A.M.; Winblad, B.; Jonsson, L.; Liu, Z.; Prince, M. The worldwide costs of dementia 2015 and comparisons with 2010. Alzheimers Dement. 2017, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Global HIV & AIDS Statistics 2019. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 1 April 2022).
- Diagnoses of HIV Infection in the United States and Dependent Areas. 2016. Available online: https://www.cdc.gov/hiv/pdf/library/reports/surveillance/cdc-hiv-surveillance-report-2016-vol-28.pdf (accessed on 1 April 2022).
- Morgello, S.; Jacobs, M.; Murray, J.; Byrd, D.; Neibart, E.; Mintz, L.; Meloni, G.; Chon, C.; Crary, J. Alzheimer’s disease neuropathology may not predict functional impairment in HIV: A report of two individuals. J. Neurovirol. 2018, 24, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, M.; Yadav, S.; Kumar, V.; Kumar, R.; Tripathi, M.; Gaikwad, S.; Kumar, P.; Bal, C. HIV encephalitis with subcortical tau deposition: Imaging pathology in vivo using F-18 THK 5117. Eur. J. Nucl. Med. Mol. Imag. 2016, 43, 2456–2457. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.S.; Chadwick, M.; Horton, W.A.; Simon, G.L.; Jiang, X.; Esposito, G. An individual with human immunodeficiency virus, dementia, and central nervous system amyloid deposition. Alzheimers Dement. 2016, 4, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Clifford, D.B.; Fagan, A.M.; Holtzman, D.M.; Morris, J.C.; Teshome, M.; Shah, A.R.; Kauwe, J.S. CSF biomarkers of Alzheimer disease in HIV-associated neurologic disease. Neurology 2009, 73, 1982–1987. [Google Scholar] [CrossRef]
- Patrick, C.; Crews, L.; Desplats, P.; Dumaop, W.; Rockenstein, E.; Achim, C.L.; Everall, I.P.; Masliah, E. Increased CDK5 expression in HIV encephalitis contributes to neurodegeneration via tau phosphorylation and is reversed with Roscovitine. Am. J. Pathol. 2011, 178, 1646–1661. [Google Scholar] [CrossRef]
- Aksenov, M.Y.; Aksenova, M.V.; Mactutus, C.F.; Booze, R.M. HIV-1 protein-mediated amyloidogenesis in rat hippocampal cell cultures. Neurosci. Lett. 2010, 475, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Bae, M.; Patel, N.; Xu, H.; Lee, M.; Tominaga-Yamanaka, K.; Nath, A.; Geiger, J.; Gorospe, M.; Mattson, M.P.; Haughey, N.J. Activation of TRPML1 clears intraneuronal Abeta in preclinical models of HIV infection. J. Neurosci. 2014, 34, 11485–11503. [Google Scholar] [CrossRef] [Green Version]
- Guindon, J.; Blanton, H.; Brauman, S.; Donckels, K.; Narasimhan, M.; Benamar, K. Sex Differences in a Rodent Model of HIV-1-Associated Neuropathic Pain. Int. J. Mol. Sci. 2019, 20, 1196. [Google Scholar] [CrossRef] [Green Version]
- Herzberg, U.; Sagen, J. Peripheral nerve exposure to HIV viral envelope protein gp120 induces neuropathic pain and spinal gliosis. J. Neuroimmunol. 2001, 116, 29–39. [Google Scholar] [CrossRef]
- Wallace, V.C.; Blackbeard, J.; Segerdahl, A.R.; Hasnie, F.; Pheby, T.; McMahon, S.B.; Rice, A.S. Characterization of rodent models of HIV-gp120 and anti-retroviral-associated neuropathic pain. Brain 2007, 130, 2688–2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, S.B.; Shi, Y.; Chen, J.; Zhou, X.; Li, G.; Gelman, B.B.; Lisinicchia, J.G.; Carlton, S.M.; Ferguson, M.R.; Tan, A.; et al. Gp120 in the pathogenesis of human immunodeficiency virus-associated pain. Ann. Neurol. 2014, 75, 837–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendash, G.W.; Lewis, J.; Leighty, R.E.; McGowan, E.; Cracchiolo, J.R.; Hutton, M.; Garcia, M.F. Multi-metric behavioral comparison of APPsw and P301L models for Alzheimer’s disease: Linkage of poorer cognitive performance to tau pathology in forebrain. Brain Res. 2004, 1012, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, M.; George, M.; Bunquin, L.E.; Bose, C.; Reddy, P.H. Protective effects of a small-molecule inhibitor DDQ against tau-induced toxicities in a transgenic tau mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2022, 31, 1022–1034. [Google Scholar] [CrossRef]
- Rappold, P.M.; Cui, M.; Grima, J.C.; Fan, R.Z.; de Mesy-Bentley, K.L.; Chen, L.; Zhuang, X.; Bowers, W.J.; Tieu, K. Drp1 inhibition attenuates neurotoxicity and dopamine release deficits in vivo. Nat. Commun. 2014, 5, 5244. [Google Scholar] [CrossRef]
- Benamar, K.; Addou, S.; Yondorf, M.; Geller, E.B.; Eisenstein, T.K.; Adler, M.W. Intrahypothalamic injection of the HIV-1 envelope glycoprotein induces fever via interaction with the chemokine system. J. Pharmacol. Exp. Ther. 2010, 332, 549–553. [Google Scholar] [CrossRef] [Green Version]
- Barak, O.; Goshen, I.; Ben-Hur, T.; Weidenfeld, J.; Taylor, A.N.; Yirmiya, R. Involvement of brain cytokines in the neurobehavioral disturbances induced by HIV-1 glycoprotein120. Brain Res. 2002, 933, 98–108. [Google Scholar] [CrossRef]
- Opp, M.R.; Rady, P.L.; Hughes, T.K., Jr.; Cadet, P.; Tyring, S.K.; Smith, E.M. Human immunodeficiency virus envelope glycoprotein 120 alters sleep and induces cytokine mRNA expression in rats. Am. J. Physiol. 1996, 270, R963–R970. [Google Scholar]
- Shanmugam, S.; Patel, D.; Guindon, J.; Reddy, P.H.; Narasimhan, M.; Benamar, K. Gene expression of endocannabinoid system in HIV-1-related neuropathic pain model. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165891. [Google Scholar] [CrossRef]
- Lewis, J.; McGowan, E.; Rockwood, J.; Melrose, H.; Nacharaju, P.; Van Slegtenhorst, M.; Gwinn-Hardy, K.; Paul Murphy, M.; Baker, M.; Yu, X.; et al. Neurofibrillary tangles, amyotrophy and progressive motor disturbance in mice expressing mutant (P301L) tau protein. Nat. Genet. 2000, 25, 402–405. [Google Scholar] [CrossRef]
- Kandimalla, R.; Manczak, M.; Yin, X.; Wang, R.; Reddy, P.H. Hippocampal phosphorylated tau induced cognitive decline, dendritic spine loss and mitochondrial abnormalities in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2018, 27, 30–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, P.H.; McWeeney, S. Mapping cellular transcriptosomes in autopsied Alzheimer’s disease subjects and relevant animal models. Neurobiol. Aging 2006, 27, 1060–1077. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H.; Mani, G.; Park, B.S.; Jacques, J.; Murdoch, G.; Whetsell, W., Jr.; Kaye, J.; Manczak, M. Differential loss of synaptic proteins in Alzheimer’s disease: Implications for synaptic dysfunction. J. Alzheimers Dis. 2005, 7, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Manczak, M.; Calkins, M.J.; Reddy, P.H. Impaired mitochondrial dynamics and abnormal interaction of amyloid beta with mitochondrial protein Drp1 in neurons from patients with Alzheimer’s disease: Implications for neuronal damage. Hum. Mol. Genet. 2011, 20, 2495–2509. [Google Scholar] [CrossRef]
- Manczak, M.; Reddy, P.H. Abnormal interaction of VDAC1 with amyloid beta and phosphorylated tau causes mitochondrial dysfunction in Alzheimer’s disease. Hum. Mol. Genet. 2012, 21, 5131–5146. [Google Scholar] [CrossRef]
- Kandimalla, R.; Manczak, M.; Fry, D.; Suneetha, Y.; Sesaki, H.; Reddy, P.H. Reduced dynamin-related protein 1 protects against phosphorylated Tau-induced mitochondrial dysfunction and synaptic damage in Alzheimer’s disease. Hum. Mol. Genet. 2016, 25, 4881–4897. [Google Scholar] [CrossRef] [Green Version]
- Manczak, M.; Kandimalla, R.; Fry, D.; Sesaki, H.; Reddy, P.H. Protective effects of reduced dynamin-related protein 1 against amyloid beta-induced mitochondrial dysfunction and synaptic damage in Alzheimer’s disease. Hum. Mol. Genet. 2016, 25, 5148–5166. [Google Scholar] [CrossRef] [Green Version]
- Manczak, M.; Kandimalla, R.; Yin, X.; Reddy, P.H. Hippocampal mutant APP and amyloid beta-induced cognitive decline, dendritic spine loss, defective autophagy, mitophagy and mitochondrial abnormalities in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2018, 27, 1332–1342. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.H.; Yin, X.; Manczak, M.; Kumar, S.; Pradeepkiran, J.A.; Vijayan, M.; Reddy, A.P. Mutant APP and amyloid beta-induced defective autophagy, mitophagy, mitochondrial structural and functional changes and synaptic damage in hippocampal neurons from Alzheimer’s disease. Hum. Mol. Genet. 2018, 27, 2502–2516. [Google Scholar] [CrossRef]
- Fields, J.A.; Serger, E.; Campos, S.; Divakaruni, A.S.; Kim, C.; Smith, K.; Trejo, M.; Adame, A.; Spencer, B.; Rockenstein, E.; et al. HIV alters neuronal mitochondrial fission/fusion in the brain during HIV-associated neurocognitive disorders. Neurobiol. Dis. 2016, 86, 154–169. [Google Scholar] [CrossRef] [Green Version]
- Rozzi, S.J.; Avdoshina, V.; Fields, J.A.; Mocchetti, I. Human immunodeficiency virus Tat impairs mitochondrial fission in neurons. Cell Death Discov. 2018, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Teodorof-Diedrich, C.; Spector, S.A. Human Immunodeficiency Virus Type 1 gp120 and Tat Induce Mitochondrial Fragmentation and Incomplete Mitophagy in Human Neurons. J. Virol. 2018, 92, e00993-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benamar, K.; Geller, E.B.; Adler, M.W. Elevated level of the proinflammatory chemokine, RANTES/CCL5, in the periaqueductal grey causes hyperalgesia in rats. Eur. J. Pharmacol. 2008, 592, 93–95. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.A.F.K. The Mouse Brain in Streotaxic Coordinates, 2nd ed.; Academic Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Benamar, K.; Rawls, S.M.; Geller, E.B.; Adler, M.W. Intrahypothalamic injection of deltorphin-II alters body temperature in rats. Brain Res. 2004, 1019, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Benamar, K.; McMenamin, M.; Geller, E.B.; Chung, Y.G.; Pintar, J.E.; Adler, M.W. Unresponsiveness of mu-opioid receptor knockout mice to lipopolysaccharide-induced fever. Br. J. Pharmacol. 2005, 144, 1029–1031. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vijayan, M.; Yin, L.; Reddy, P.H.; Benamar, K. Behavioral Evidence for a Tau and HIV-gp120 Interaction. Int. J. Mol. Sci. 2022, 23, 5514. https://doi.org/10.3390/ijms23105514
Vijayan M, Yin L, Reddy PH, Benamar K. Behavioral Evidence for a Tau and HIV-gp120 Interaction. International Journal of Molecular Sciences. 2022; 23(10):5514. https://doi.org/10.3390/ijms23105514
Chicago/Turabian StyleVijayan, Murali, Linda Yin, P. Hemachandra Reddy, and Khalid Benamar. 2022. "Behavioral Evidence for a Tau and HIV-gp120 Interaction" International Journal of Molecular Sciences 23, no. 10: 5514. https://doi.org/10.3390/ijms23105514
APA StyleVijayan, M., Yin, L., Reddy, P. H., & Benamar, K. (2022). Behavioral Evidence for a Tau and HIV-gp120 Interaction. International Journal of Molecular Sciences, 23(10), 5514. https://doi.org/10.3390/ijms23105514