
A Single Injection of NTG-101 Reduces the Expression of Pain-Related Neurotrophins in a Canine Model of Degenerative Disc Disease

 and
and 
Abstract
:1. Introduction
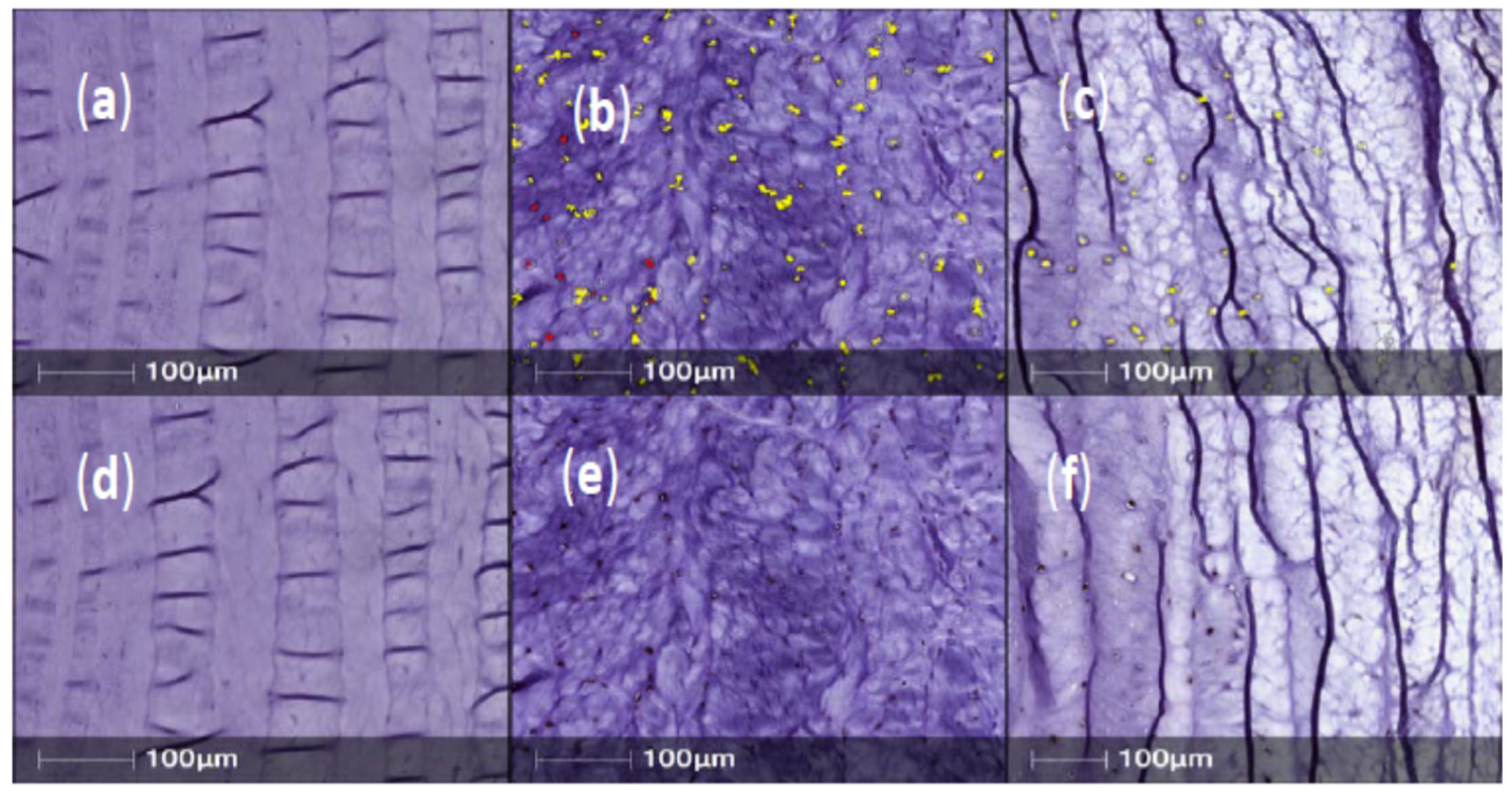
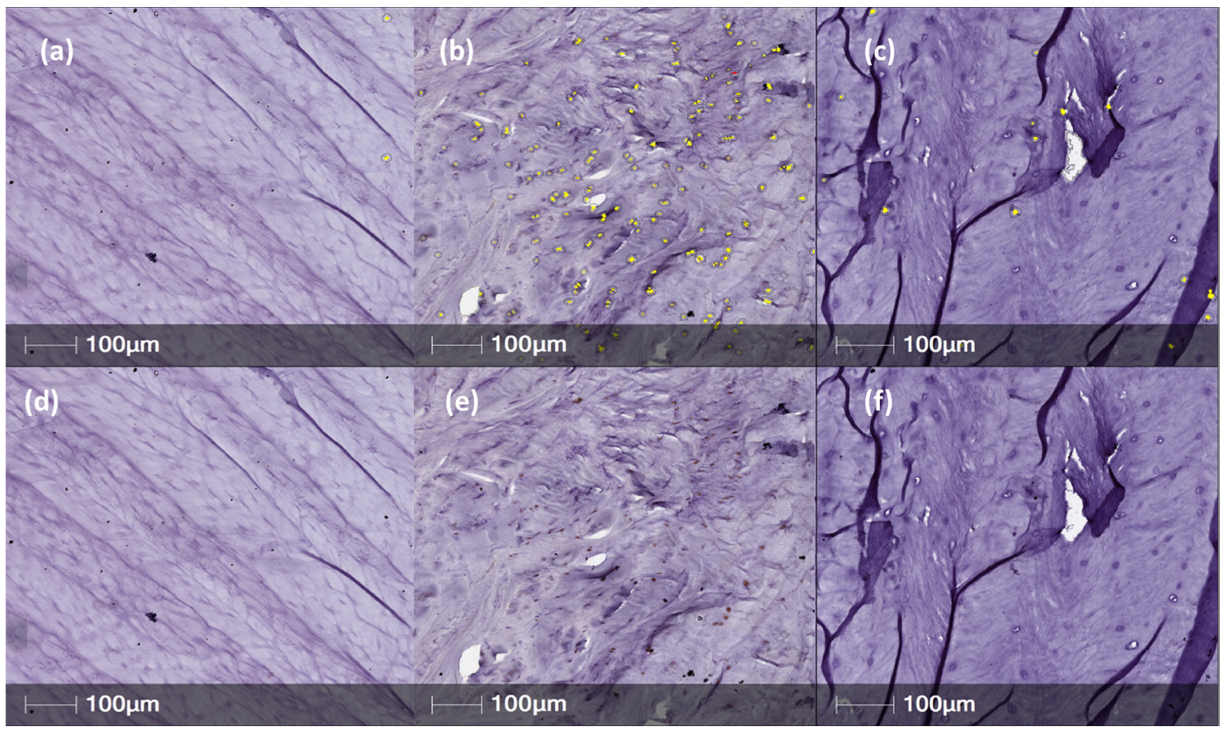
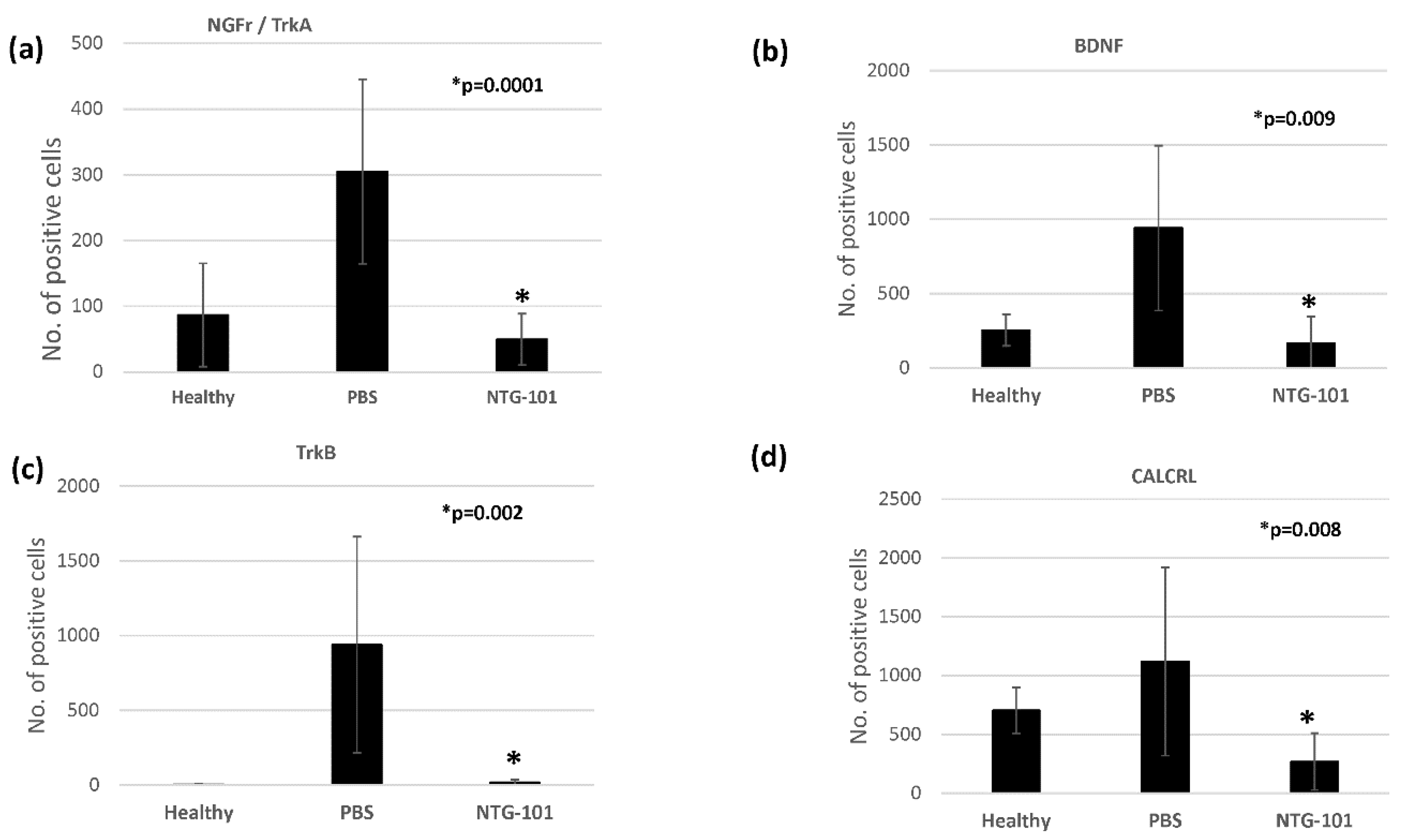
2. Results
In Vivo Expression of Neurotrophins/Pain Associated Proteins
3. Discussion
4. Materials and Methods
4.1. Induced Degenerative Disc Disease (DDD) in Chondrodystrophic Canines
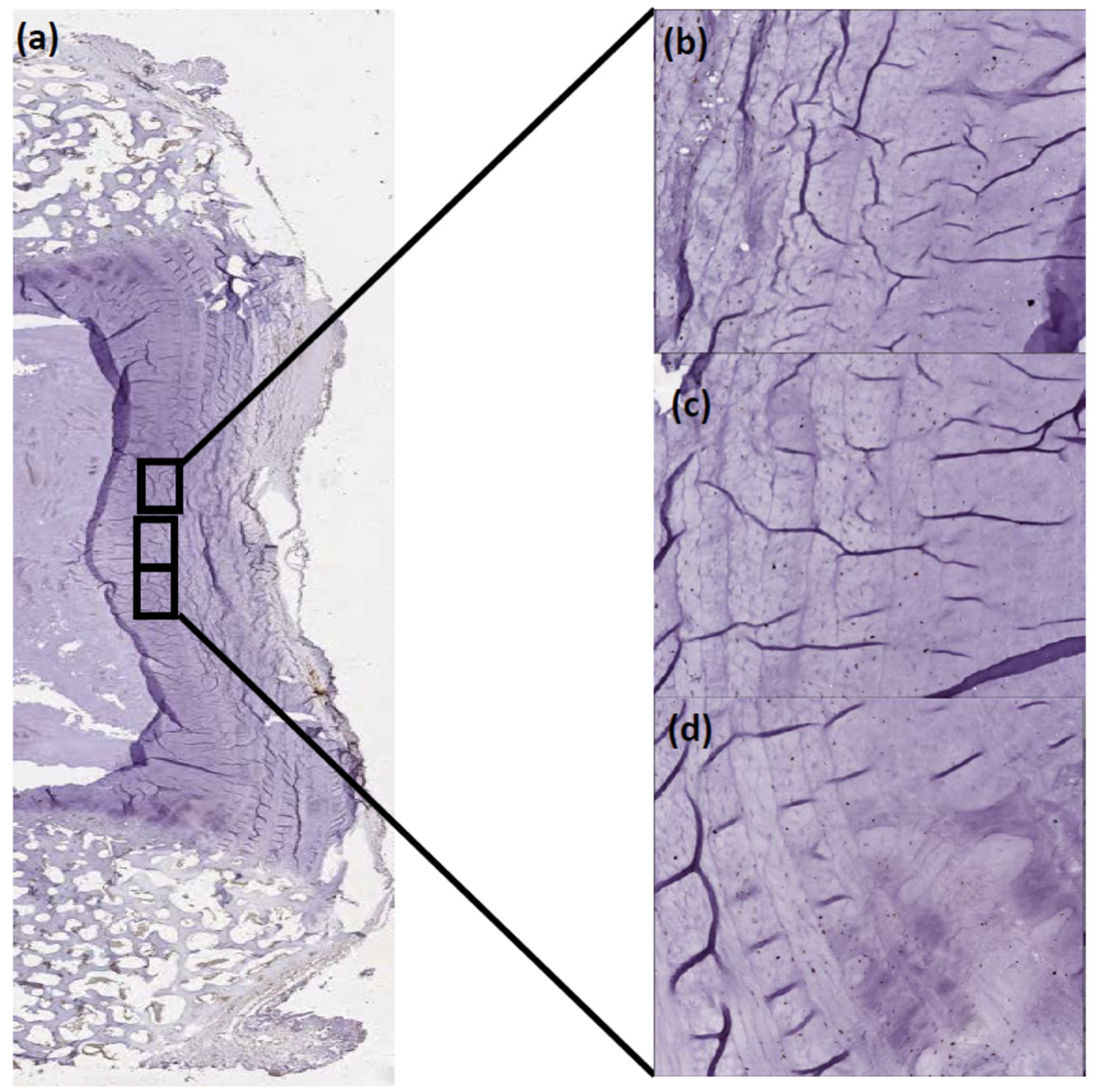
4.2. Tissue Preparation
Immunohistochemistry (IHC)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BDNF | Brain Derived Nerve Factor |
| CGRP | Calcitonin Gene Related Peptide |
| CALCRL | Calcitonin Receptor Like Receptor |
| CTGF | Connective Tissue Growth Factor |
| DDD | Degenerative Disc Disease |
| DRG | Dorsal Root Ganglion |
| GDNF | Glia Cell Line-Derived Neurotrophic Factor |
| HRP | Horse Radish Peroxidases |
| IL-1β | Interleukin- 1 beta |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| IVD | Intervertebral Disc |
| NGFr | Nerve Growth Factor receptor |
| NTC | No Treatment Controls |
| NFkB | Nuclear Factor Kappa B |
| PBS | Phosphate Buffered Saline |
| P38MAPK | P38 Mitogen Activated Protein Kinase |
| TGF-β1 | Transforming Growth Factor Beta-1 |
| TBS-T | Tris-Buffered Saline and Tween 20 |
| TrkA | Tyrosine Kinase Receptor A |
| TrkB | Tyrosine Kinase Receptor B |
| TNF-α | Tumor Necrosis Factor alpha |
| rh | Recombinant Human |
| ROI | Regions of Interest |
Appendix A

References
- Kim, L.H.; Vail, D.; Azad, T.D.; Bentley, J.P.; Zhang, Y.; Ho, A.L.; Fatemi, P.; Feng, A.; Varshneya, K.; Desai, M.; et al. Expenditures and Health Care Utilization Among Adults With Newly Diagnosed Low Back and Lower Extremity Pain. JAMA Netw. Open 2019, 2, e193676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geurts, J.W.; Willems, P.C.; Kallewaard, J.-W.; van Kleef, M.; Dirksen, C. The impact of chronic discogenic low back pain: Costs and patient’s burden. Pain Res. Manag. 2018, 2018, 4696180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung Kennetch, M.C.; Karppinen, J.; Chan, D.; Ho Daniel, W.H.; Song, Y.-Q.; Sham, P.; Cheah Kathryn, S.E.; Leong John, C.Y.; Luk Keith, D.K. Prevalence and pattern of lumbar magnetic resonance imaging changes in a population of one thousand forty-three individuals. Spine 2009, 34, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.A.; Roughley, P.J. What is intervertebral disc degeneration and what causes it? Spine 2006, 31, 2151–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freemont, A.J. The cellular pathobiology of the degenerate intervertebral disc and discogenic pain. Rheumatology 2009, 48, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Matta, A.; Karim, M.Z.; Isenman, D.E.; Erwin, W.M. Molecular therapy for degenerative disc disease: Clues from secretome analysis of the notochordal cell-rich nucleus pulposus. Sci. Rep. 2017, 7, 45623. [Google Scholar] [CrossRef] [Green Version]
- Matta, A.; Karim, M.Z.; Gerami, H.; Jun, P.; Funabashi, M.; Kawchuck, G.; Goldstein, A.; Foltz, W.; Sussman, M.; Eek Bjorn, C.; et al. NTG-101: A novel molecular therapy that halts the progression of degenerative disc disease. Sci. Rep. 2018, 8, 16809. [Google Scholar] [CrossRef]
- Matta, A.; Karim, M.Z.; Gerami, H.; Benigno, B.; Erwin, W.M. A comparative study of mesenchymal stem cell transplantation and NTG-101 molecular therapy to treat degenerative disc disease. Sci. Rep. 2021, 11, 14804. [Google Scholar] [CrossRef]
- Garcia-Cosamalon, J.; del Valle Miguel, E.; Calavia Marta, G.; Garcia-Suarez, O.; Lopez-Muniz, A.; Otero, J.; Vega Jose, A. Intervertebral disc, sensory nerves and neurotrophins: Who is who in discogenic pain? J. Anat. 2010, 217, 1–15. [Google Scholar] [CrossRef]
- Kartha, S.; Zeeman Martha, E.; Baig Hassam, A.; Guarino Benjamin, B.; Winkelstein Beth, A. Upregulation of BDNF and NGF in cervical intervertebral discs exposed to painful whole body vibration. Spine 2014, 1, 1542–1548. [Google Scholar] [CrossRef] [Green Version]
- Krock, E.; Rosenzweig Derek, H.; Chabot-Dore, A.-J.; Jarzem, P.; Weber, M.; Ouellet Jean, A.; Stone Laura, S.; Lisbet, H. Painful degenerating intervertebral discs up-regulate neurite sprouting and CGRP through nociceptive factors. J. Cell. Mol. Med. 2014, 18, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Huang Eric, J.; Reichardt Lous, F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezet, S.; McMahon, S.B. Neurotrophins: Mediators and Modulators of Pain. Annu. Rev. Neurosci. 2006, 29, 507–538. [Google Scholar] [CrossRef] [PubMed]
- Binch Abbie, L.A.; Cole Ashley, A.; Breakwell Lee, M.; Michael Anthony, L.R.; Chiverton, N.; Cross Alison, K.; Le Maitre Christine, L. Expression and regulation of neurotrophic and angiogenic factors during human intervertbral disc degeneration. Arthritis Res. Ther. 2014, 16, 4116. [Google Scholar]
- Freemont, A.J.; Peacock, T.E.; Goupille, P.; Hoyland, J.A.; O’Brien, J.; Jayson, M.I.V. Nerve Ingrowth into Diseased Intervertebral Disc in Chronic Back Pain. Lancet 1997, 350, 178–181. [Google Scholar] [CrossRef]
- Freemont, A.J.; Watkins, A.; Le Maitre, C.; Baird, P.; Jeziorska, M.; Knight, M.T.N.; Ross, E.R.S.; O’Brien, J.P.; Hoyland, J.A. Nerve growth factor expression and innervation of the painful intervertebral disc. J. Pathol. 2002, 197, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, I.G.; Aoki, Y.; Ochiai, N.; Kishida, S.; Endo, M.; Yamashita, T.; Takahashi, K.; Ohtori, S.; Koshi, T.; Yamashita, M.; et al. Nerve Growth Factor of Cultured Medium Extracted From Human Degenerative Nucleus Pulposus Promotes Sensory Nerve Growth and Induces Substance P In Vitro. Spine 2009, 34, 2263–2269. [Google Scholar] [CrossRef]
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.B.; et al. Neuropathic pain. Nat. Rev. Dis. Primers 2017, 3, 17002. [Google Scholar] [CrossRef] [Green Version]
- Gruber, H.E.; Hoelschher, G.L.; Bethea, S.; Hanley, E.N. Interleukin 1-beta upregulates brain-derived neurotrophic factor, neurotrophin 3 and neuropilin 2 gene expression and NGF production in annulus cells. Biotech. Histochem. 2012, 87, 506–511. [Google Scholar] [CrossRef]
- Sugiura, A.; Ohtori, S.; Yamashita, M.; Yamauchi, K.; Inoue, G.; Susuki, M.; Norimoto, M.; Orita, S.; Eguchi, Y.; Kuniyoshi, K.; et al. Effect of applying p75NTR saporin to a punctured intervertebral disc on calcitonin gene-related peptide expression in rat dorsal root ganglions. J. Orthop. Sci. 2010, 15, 407–413. [Google Scholar] [CrossRef]
- Navone Stefania, E.; Marfia, G.; Canzi, L.; Ciusani, E.; Canazza, A.; Visintini, S.; Campanella, R.; Parati Eugenio, A. Expression of neural and neurotrophic markers in nucleus pulposus cells isolated from degenerated intervertebral disc. J. Orthop. Res. 2012, 30, 1470–1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purmessur, D.; Freemont, A.J.; Hoyland, J.A. Expression and regulation of neurotrophins in the nondegenerate and degenerate human intervertebral disc. Arthritis Res. Ther. 2008, 10, R99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber Helen, E.; Ingram Jane, A.; Hoelscher, G.; Zinchenko, N.; Norton, H.J.; Hanley Edward, N., Jr. Brain-derived neurotrophic factor and its receptor in the human and the sand rat intervertebral disc. Arthritis Res. Ther. 2008, 10, R82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deuis Jennifer, R.; Dvorakova, L.S.; Irina, V. Methods Used to Evaluate Pain Behavior in Rodents. Front. Mol. Neurosci. 2017, 10, 204. [Google Scholar] [CrossRef] [Green Version]
- Burke, J.G.; Watson, R.W.G.; McCormack, D.; Dowling, F.E.; Walsh, M.G.; Fitzpatrick, J.M. Intervertebral discs which cause low back pain secrete high levels of proinflammatory mediators. J. Bone Jt. Surg. 2002, 84-B, 196–201. [Google Scholar] [CrossRef]
- Bogduk, N.; Tynan, W.; Wilson, A.S. The nerve supply to the human lumbar intervertebral discs. J. Anat. 1981, 132, 39–56. [Google Scholar]
- Fagan, A.; Moore, R.; Vernon Roberts, B.; Blumbergs, P.; Fraser, R. ISSLS prize winner: The innervation of the intervertebral disc: A quantitative analysis. Spine 2003, 28, 2570–2576. [Google Scholar] [CrossRef]
- Lyu, F.-J.; Cui, H.; Pah, H.; Cheung, K.M.C.; Cao, X.; Iatridis, J.C.; Zheng, Z. Painful intervertebral disc degeneration and inflammation: From laboratory evidence to clinical interventions. Bone Res. 2021, 9, 7. [Google Scholar] [CrossRef]
- Aoki, Y.; Ohtori, S.; Kazuhisa, T.; Ino, H.; Yakahashi, Y.; Chiba, T.; Moriya, H. Innervation of the lumbar intervertebral disc by nerve growth factor-dependent neurons related to inflammatory pain. Spine 2004, 29, 1077–1081. [Google Scholar] [CrossRef]
- Matta, A.; Erwin, W.M. Current Status of the Instructional Cues Provided by Notochordal Cells in Novel Disc Repair Strategies. Int. J. Mol. Sci. 2022, 23, 427. [Google Scholar] [CrossRef]
- Matta, A.; Karim, M.Z.; Isenman, D.; Erwin, W.M. Regenerative therapy for degenerative disc disease: Clues from secretome analysis of notochordal cells. In Proceedings of the the Ninth Annual Meeting of the Lumbar Spine Research Society, Chicago, IL, USA, 14–15 April 2016. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Age (Years) | Gender | Spinal Level | Pfirmann Grade | Modic Type | NGFr * | BDNF * | TrkB * | CALCRL * |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 51 | F | L5-S1 | III | 0 | 1 | 1 | 1 | 1 |
| 2 | 59 | F | L4-L5 | II | 0 | 1 | 1 | 1 | 1 |
| 3 | 79 | F | L5-S1 | IV | 0 | 1 | 1 | 1 | 1 |
| 4 | 67 | F | L5-S1 | III | 1 | 1 | 1 | 1 | 1 |
| 5 | 57 | F | L5-S1 | III | 1 | 1 | 1 | 1 | 1 |
| 6 | 66 | F | L4–5 | IV | 0 | 1 | 1 | 1 | 1 |
| 7 | 78 | F | L2–3 | III | 3 | 1 | 1 | 1 | 1 |
| 8 | 32 | F | L5-S1 | IV | 1 | 1 | 1 | 0 | 1 |
| 9 | 30 | F | L5-S1 | IV | 0 | 1 | 1 | 1 | 1 |
| 10 | 71 | M | L5-S1 | II | 0 | 1 | 0 | 1 | 1 |
| 11 | 47 | M | L5-S1 | III | 0 | 1 | 1 | 1 | 1 |
| 12 | 54 | F | L4–5 | III | 0 | 0 | 0 | 1 | 1 |
| 13 | 78 | M | L2–3 | V | 1 | 1 | 1 | 1 | 1 |
| 14 | 72 | M | L4–5 | III | 2 | 0 | 0 | 1 | 1 |
| 15 | 27 | M | L4–5 | III | 0 | 1 | 1 | 1 | 1 |
| S. No. | IHC Protein | Antibody Source Company | Primary Antibody Cat# and Dilution Used |
|---|---|---|---|
| 1. | NGFr/TrkA | R&D Systems | MAB367 (Mouse Monoclonal, 1:50 dilution) |
| 2. | BDNF | Biorbyt | Orb251616 (Rabbit pAb, 1:100 dilution). |
| 3. | TrkB | Biorbyt | Orb214339 (Rabbit pAb, 1:100 dilution) |
| 4. | CALCRL | Antibodies Online | ABIN1048377 (Rabbit Polyclonal, 1:200 dilution) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matta, A.; Karim, M.Z.; Gerami, H.; Benigno, B.Z.; Cheng, I.; Mehrkens, A.; Erwin, W.M. A Single Injection of NTG-101 Reduces the Expression of Pain-Related Neurotrophins in a Canine Model of Degenerative Disc Disease. Int. J. Mol. Sci. 2022, 23, 5717. https://doi.org/10.3390/ijms23105717
Matta A, Karim MZ, Gerami H, Benigno BZ, Cheng I, Mehrkens A, Erwin WM. A Single Injection of NTG-101 Reduces the Expression of Pain-Related Neurotrophins in a Canine Model of Degenerative Disc Disease. International Journal of Molecular Sciences. 2022; 23(10):5717. https://doi.org/10.3390/ijms23105717
Chicago/Turabian StyleMatta, Ajay, Muhammad Zia Karim, Hoda Gerami, Bettina Zoe Benigno, Ivan Cheng, Arne Mehrkens, and William Mark Erwin. 2022. "A Single Injection of NTG-101 Reduces the Expression of Pain-Related Neurotrophins in a Canine Model of Degenerative Disc Disease" International Journal of Molecular Sciences 23, no. 10: 5717. https://doi.org/10.3390/ijms23105717
APA StyleMatta, A., Karim, M. Z., Gerami, H., Benigno, B. Z., Cheng, I., Mehrkens, A., & Erwin, W. M. (2022). A Single Injection of NTG-101 Reduces the Expression of Pain-Related Neurotrophins in a Canine Model of Degenerative Disc Disease. International Journal of Molecular Sciences, 23(10), 5717. https://doi.org/10.3390/ijms23105717