
IL-17A Is the Critical Cytokine for Liver and Spleen Amyloidosis in Inflammatory Skin Disease

 , , ,
, , ,  , and
, and 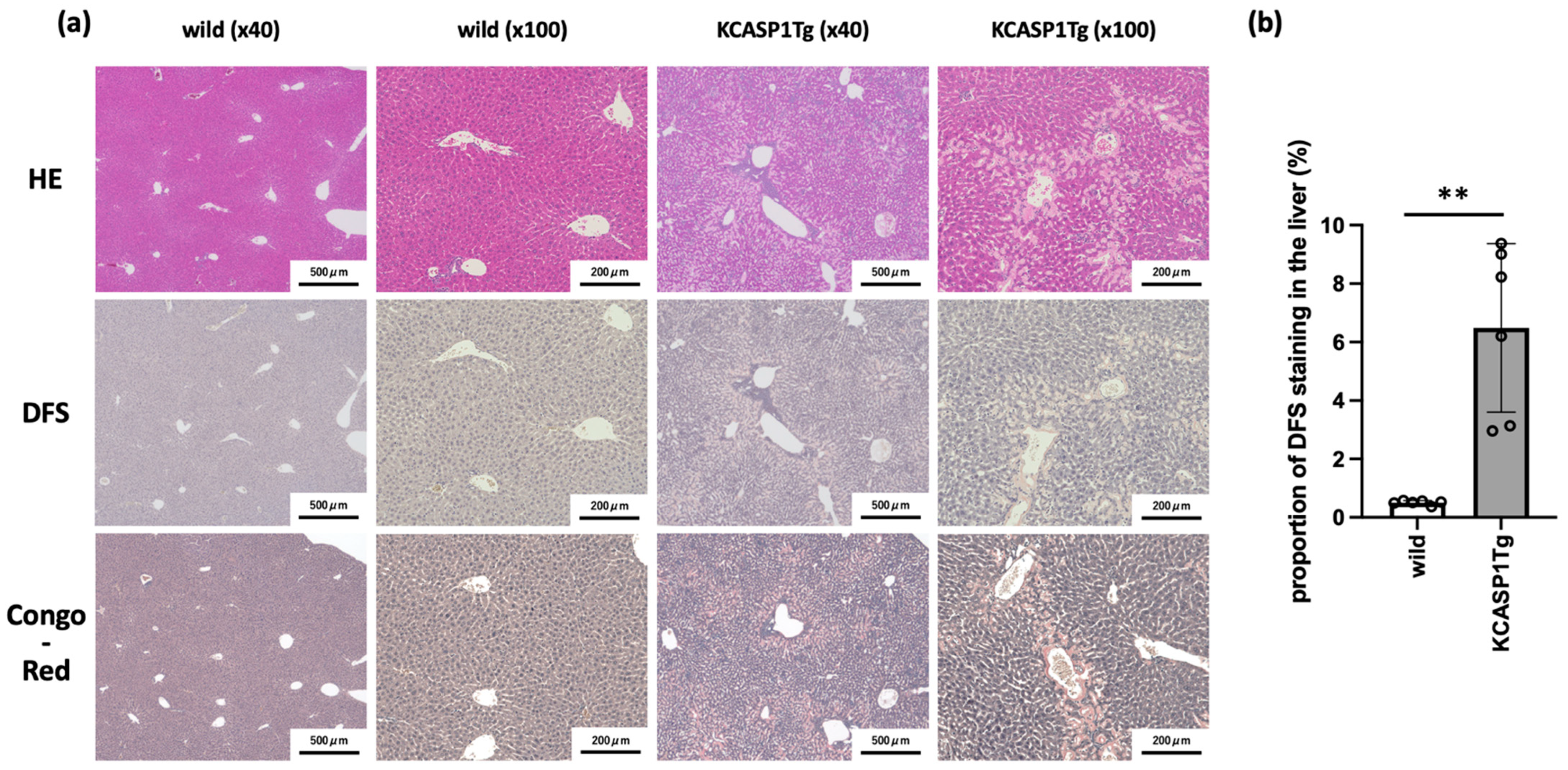{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Histological Analysis Showed the Amyloid Deposition in the Liver of KCASP1Tg Mice
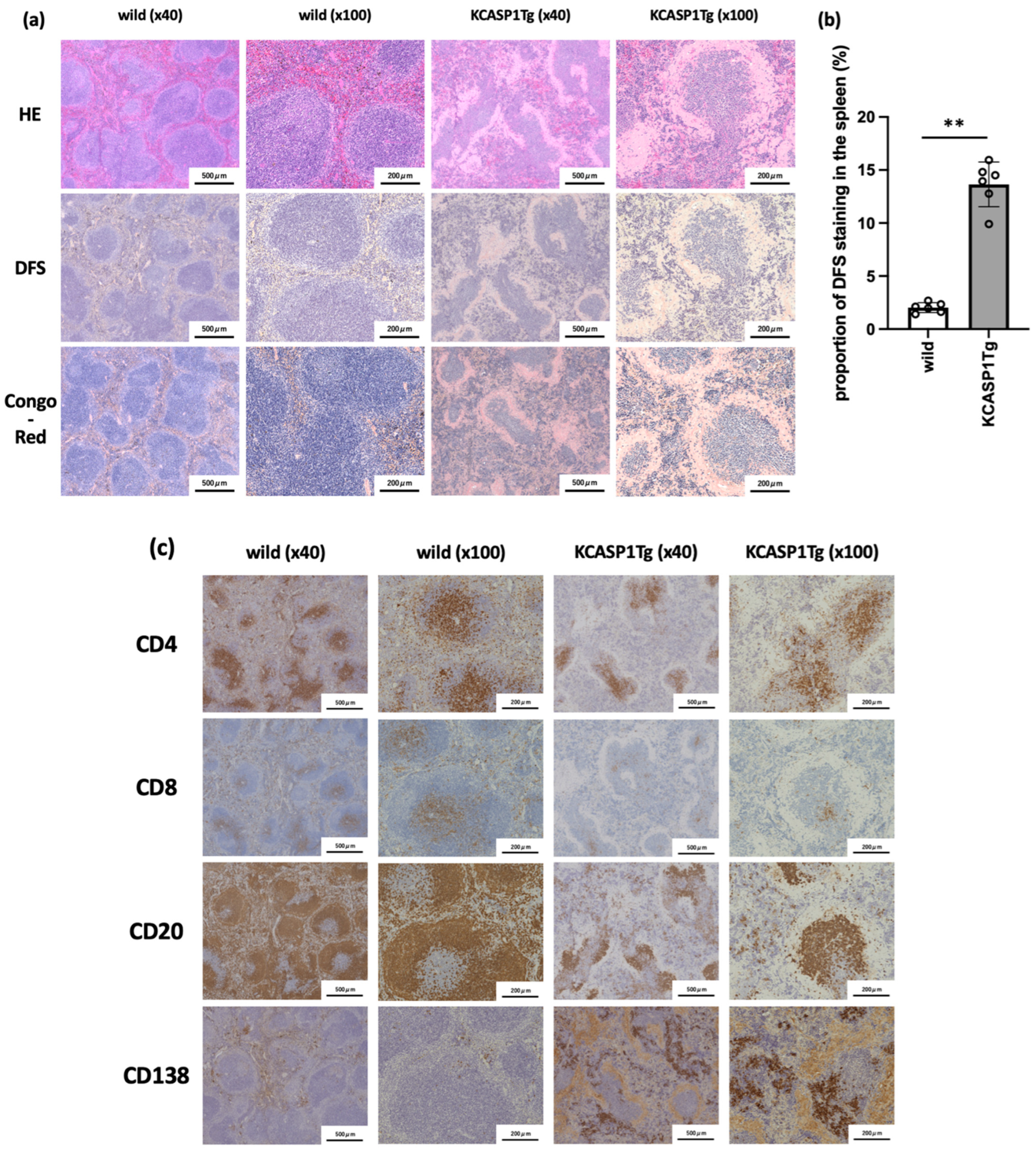
2.2. Histological Analysis of Spleen in KCASP1Tg Mice
2.3. KCASP1Tg Mice Showed Signs of Liver and Spleen Dysfunction
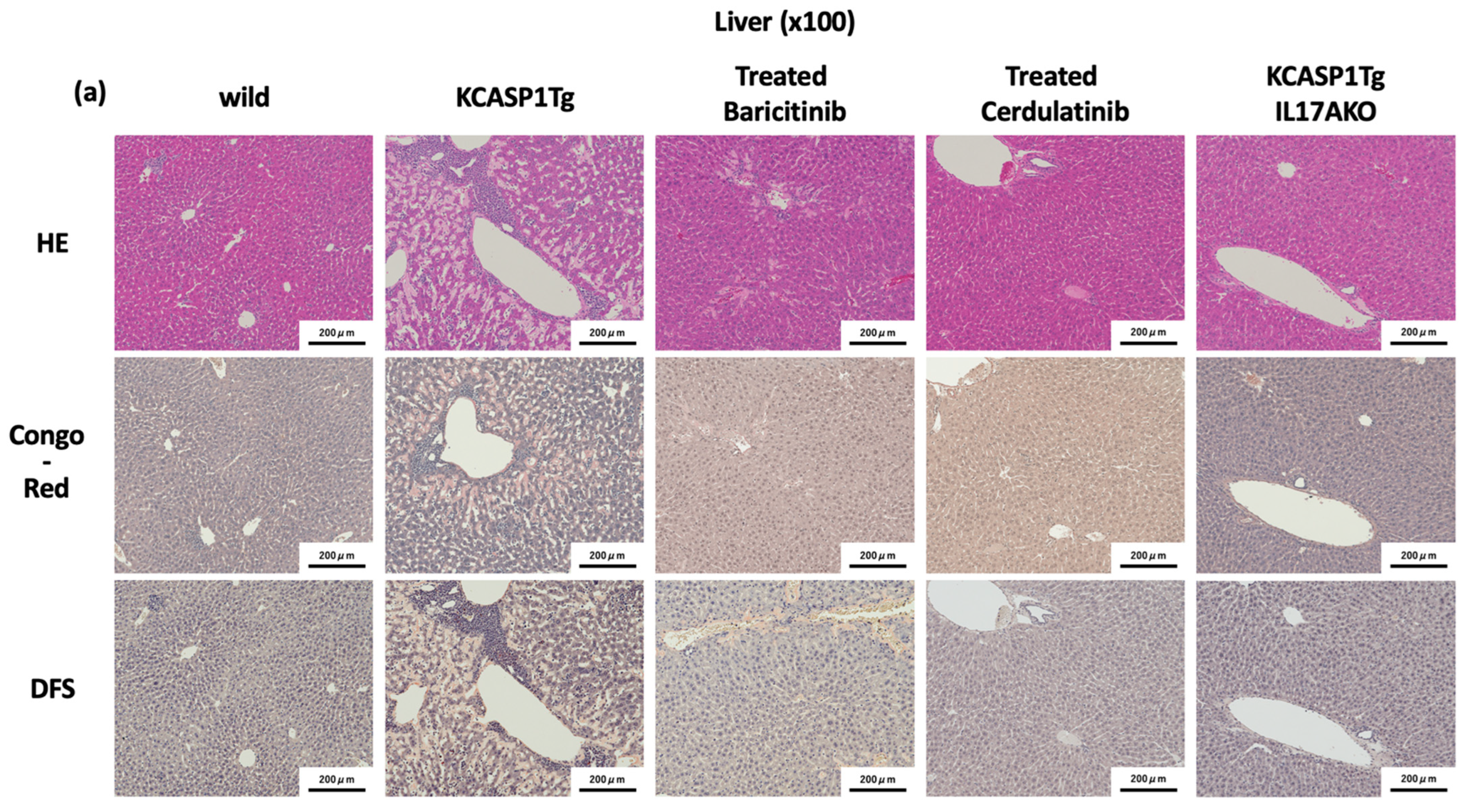
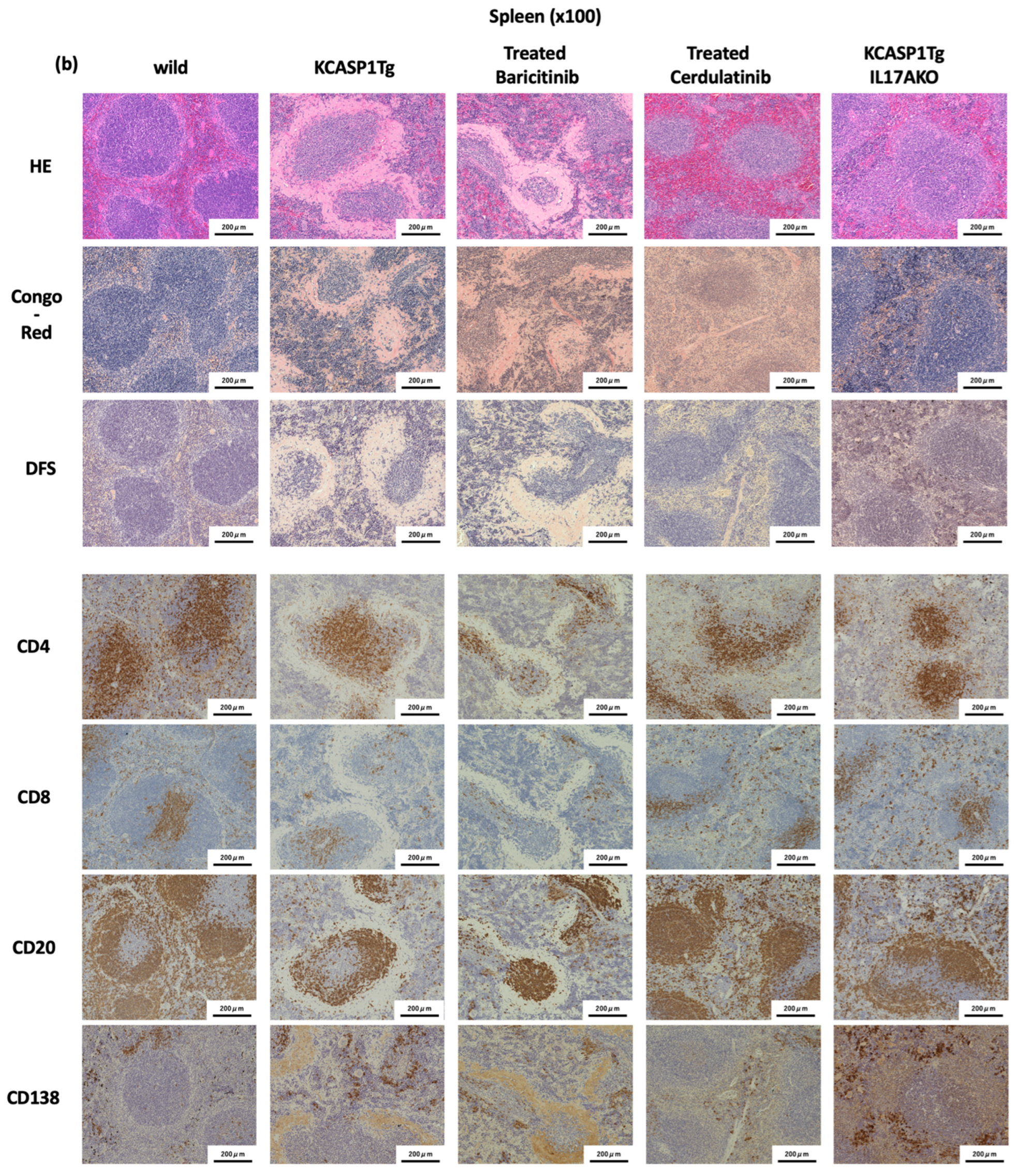
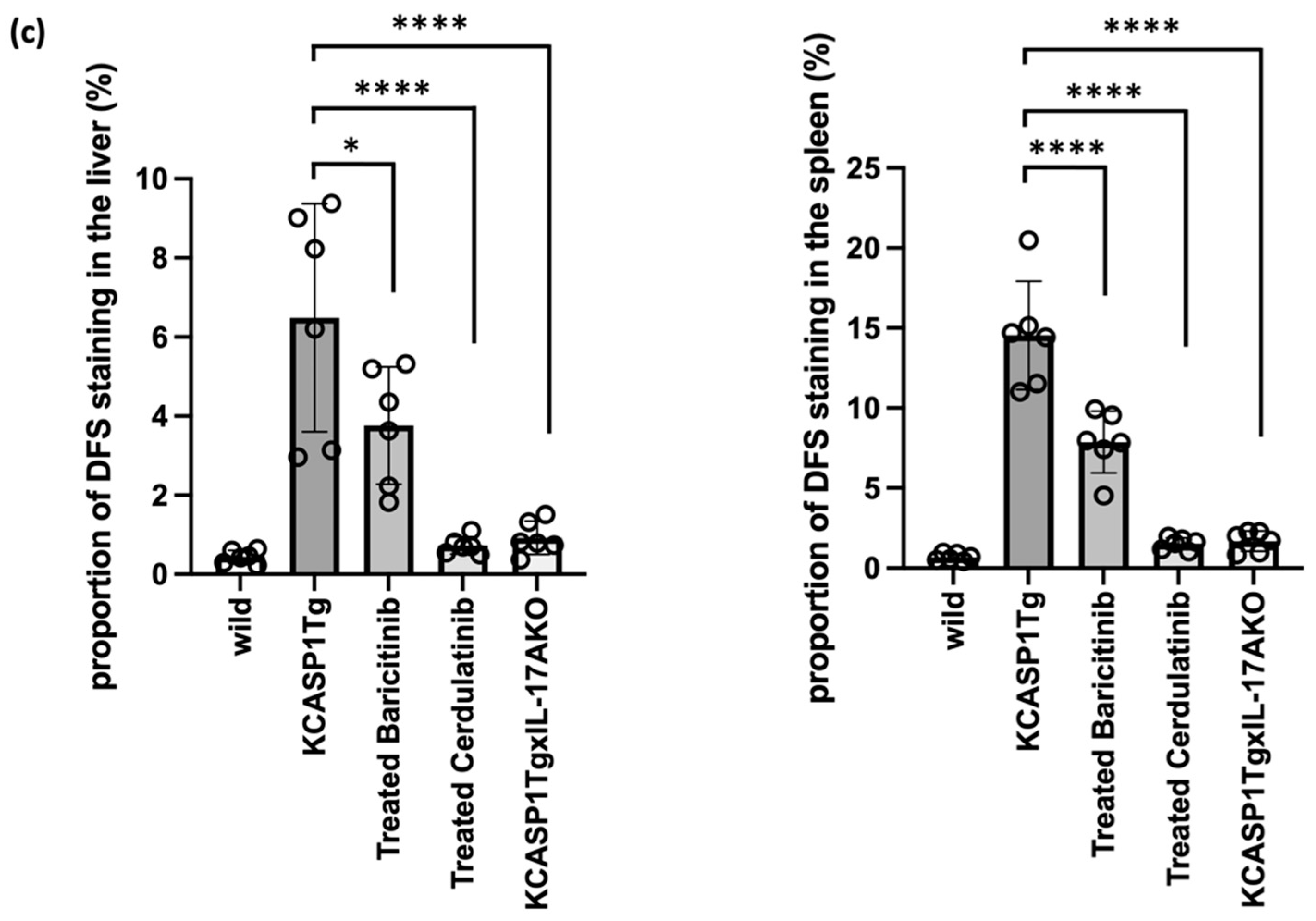
2.4. Effect of Administration of JAK Inhibitors and IL-17A Ablation
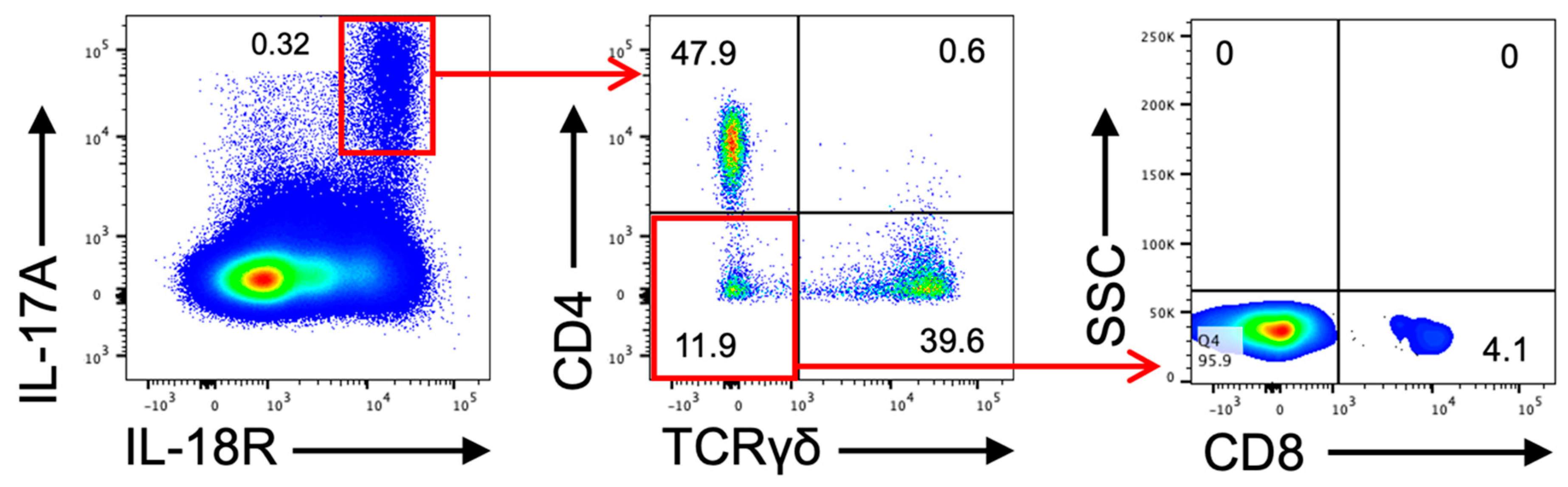
2.5. IL-17A-Producing Cell in Dermatitis Model Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Blood Sampling and Clinical Chemistry Parameters
4.3. Lymphocytes Staining in the Blood, Spleen, and Lymph Nodes
4.4. Amyloid and Infiltrating Cells Staining
4.5. Cytokine and SAA Analysis
4.6. Oral Administration of JAK Inhibitors
4.7. IL-17A-Producing Lymphatic Primary Cells
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Real de Asua, D.; Costa, R.; Galvan, J.M.; Filigheddu, M.T.; Trujillo, D.; Cadinanos, J. Systemic AA amyloidosis: Epidemiology, diagnosis, and management. Clin. Epidemiol. 2014, 6, 369–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuda, Y. AA amyloidosis—Benefits and prospects of IL-6 inhibitors. Mod. Rheumatol. 2019, 29, 268–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanaka, K.; Nakanishi, T.; Saito, H.; Maruyama, J.; Isoda, K.; Yokochi, A.; Imanaka-Yoshida, K.; Tsuda, K.; Kakeda, M.; Okamoto, R.; et al. Persistent release of IL-1s from skin is associated with systemic cardio-vascular disease, emaciation and systemic amyloidosis: The potential of anti-IL-1 therapy for systemic inflammatory diseases. PLoS ONE 2014, 9, e104479. [Google Scholar] [CrossRef]
- Nakanishi, T.; Mizutani, K.; Iida, S.; Matsushima, Y.; Umaoka, A.; Kondo, M.; Habe, K.; Yamanaka, K. Janus Kinase Inhibitors Ameliorated Gastrointestinal Amyloidosis and Hypoalbuminemia in Persistent Dermatitis Mouse Model. Int. J. Mol. Sci. 2021, 23, 28. [Google Scholar] [CrossRef]
- Matsushima, Y.; Mizutani, K.; Goto, H.; Nakanishi, T.; Kondo, M.; Habe, K.; Isoda, K.; Mizutani, H.; Yamanaka, K. Emaciation, Congestive Heart Failure, and Systemic Amyloidosis in Severe Recessive Dystrophic Epidermolysis Bullosa: Possible Internal Complications Due to Skin-Derived Inflammatory Cytokines Derived from the Injured Skin. Dermatopathology 2020, 7, 41–47. [Google Scholar] [CrossRef]
- Johnson-Huang, L.M.; McNutt, N.S.; Krueger, J.G.; Lowes, M.A. Cytokine-producing dendritic cells in the pathogenesis of inflammatory skin diseases. J. Clin. Immunol. 2009, 29, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Kato, S.; Matsushima, Y.; Mizutani, K.; Kawakita, F.; Fujimoto, M.; Okada, K.; Kondo, M.; Habe, K.; Suzuki, H.; Mizutani, H.; et al. The Stenosis of Cerebral Arteries and Impaired Brain Glucose Uptake by Long-Lasting Inflammatory Cytokine Release from Dermatitis Is Rescued by Anti-IL-1 Therapy. J. Investig. Dermatol. 2018, 138, 2280–2283. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, K.; Tanaka, M.; Tsutsui, H.; Kupper, T.S.; Asahi, K.; Okamura, H.; Nakanishi, K.; Suzuki, M.; Kayagaki, N.; Black, R.A.; et al. Skin-specific caspase-1-transgenic mice show cutaneous apoptosis and pre-endotoxin shock condition with a high serum level of IL-18. J. Immunol. 2000, 165, 997–1003. [Google Scholar] [CrossRef] [Green Version]
- Konishi, H.; Tsutsui, H.; Murakami, T.; Yumikura-Futatsugi, S.; Yamanaka, K.; Tanaka, M.; Iwakura, Y.; Suzuki, N.; Takeda, K.; Akira, S.; et al. IL-18 contributes to the spontaneous development of atopic dermatitis-like inflammatory skin lesion independently of IgE/stat6 under specific pathogen-free conditions. Proc. Natl. Acad. Sci. USA 2002, 99, 11340–11345. [Google Scholar] [CrossRef] [Green Version]
- Yoshimoto, T.; Mizutani, H.; Tsutsui, H.; Noben-Trauth, N.; Yamanaka, K.; Tanaka, M.; Izumi, S.; Okamura, H.; Paul, W.E.; Nakanishi, K. IL-18 induction of IgE: Dependence on CD4+ T cells, IL-4 and STAT6. Nat. Immunol. 2000, 1, 132–137. [Google Scholar] [CrossRef]
- Blancas-Mejia, L.M.; Ramirez-Alvarado, M. Systemic amyloidoses. Annu. Rev. Biochem. 2013, 82, 745–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanaka, K.; Okada, K.; Nakanishi, T.; Mizutani, K.; Matsushima, Y.; Kondo, M.; Habe, K.; Mizutani, H.; Seo, N. Skin inflammation leads immunoglobulin G aggregation and deposition in multiple organs. J. Dermatol. Sci 2017, 88, 146–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizutani, K.; Shirakami, E.; Ichishi, M.; Matsushima, Y.; Umaoka, A.; Okada, K.; Yamaguchi, Y.; Watanabe, M.; Morita, E.; Yamanaka, K. Systemic Dermatitis Model Mice Exhibit Atrophy of Visceral Adipose Tissue and Increase Stromal Cells via Skin-Derived Inflammatory Cytokines. Int. J. Mol. Sci. 2020, 21, 3367. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Mizutani, H. “Inflammatory skin march”: IL-1-mediated skin inflammation, atopic dermatitis, and psoriasis to cardiovascular events. J. Allergy Clin. Immunol. 2015, 136, 823–824. [Google Scholar] [CrossRef] [PubMed]
- Andersen, Y.M.F.; Egeberg, A.; Gislason, G.H.; Hansen, P.R.; Skov, L.; Thyssen, J.P. Risk of myocardial infarction, ischemic stroke, and cardiovascular death in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2016, 138, 310–312.e313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ascott, A.; Mulick, A.; Yu, A.M.; Prieto-Merino, D.; Schmidt, M.; Abuabara, K.; Smeeth, L.; Roberts, A.; Langan, S.M. Atopic eczema and major cardiovascular outcomes: A systematic review and meta-analysis of population-based studies. J. Allergy Clin. Immunol. 2019, 143, 1821–1829. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.C.; Lan, C.E. Psoriasis and Cardiovascular Comorbidities: Focusing on Severe Vascular Events, Cardiovascular Risk Factors and Implications for Treatment. Int. J. Mol. Sci. 2017, 18, 2211. [Google Scholar] [CrossRef] [Green Version]
- Gelfand, J.M.; Dommasch, E.D.; Shin, D.B.; Azfar, R.S.; Kurd, S.K.; Wang, X.; Troxel, A.B. The risk of stroke in patients with psoriasis. J. Investig. Dermatol. 2009, 129, 2411–2418. [Google Scholar] [CrossRef] [Green Version]
- Mizutani, K.; Isono, K.; Matsushima, Y.; Okada, K.; Umaoka, A.; Iida, S.; Habe, K.; Hagimori, K.; Yamazaki, H.; Yamanaka, K. Inflammatory Skin-Derived Cytokines Accelerate Osteoporosis in Mice with Persistent Skin Inflammation. Int. J. Mol. Sci. 2020, 21, 3620. [Google Scholar] [CrossRef]
- Umaoka, A.; Takeuchi, H.; Mizutani, K.; Seo, N.; Matsushima, Y.; Habe, K.; Hagimori, K.; Yamaguchi, Y.; Ikeda, T.; Yamanaka, K. Skin Inflammation and Testicular Function: Dermatitis Causes Male Infertility via Skin-Derived Cytokines. Biomedicines 2020, 8, 293. [Google Scholar] [CrossRef]
- Zhang, N.; Ahsan, M.H.; Purchio, A.F.; West, D.B. Serum amyloid A-luciferase transgenic mice: Response to sepsis, acute arthritis, and contact hypersensitivity and the effects of proteasome inhibition. J. Immunol. 2005, 174, 8125–8134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlar, C.M.; Whitehead, A.S. Serum amyloid A, the major vertebrate acute-phase reactant. Eur. J. Biochem. 1999, 265, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Palm, A.E.; Kleinau, S. Marginal zone B cells: From housekeeping function to autoimmunity? J. Autoimmun. 2021, 119, 102627. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Shelef, M.; Calame, K. Regulation of plasma-cell development. Nat. Rev. Immunol. 2005, 5, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Calame, K.L.; Lin, K.I.; Tunyaplin, C. Regulatory mechanisms that determine the development and function of plasma cells. Annu. Rev. Immunol. 2003, 21, 205–230. [Google Scholar] [CrossRef] [PubMed]
- Moser, K.; Tokoyoda, K.; Radbruch, A.; MacLennan, I.; Manz, R.A. Stromal niches, plasma cell differentiation and survival. Curr Opin. Immunol. 2006, 18, 265–270. [Google Scholar] [CrossRef]
- Lalor, S.J.; Dungan, L.S.; Sutton, C.E.; Basdeo, S.A.; Fletcher, J.M.; Mills, K.H. Caspase-1-processed cytokines IL-1beta and IL-18 promote IL-17 production by gammadelta and CD4 T cells that mediate autoimmunity. J. Immunol. 2011, 186, 5738–5748. [Google Scholar] [CrossRef] [Green Version]
- Couderc, E.; Morel, F.; Levillain, P.; Buffiere-Morgado, A.; Camus, M.; Paquier, C.; Bodet, C.; Jegou, J.F.; Pohin, M.; Favot, L.; et al. Interleukin-17A-induced production of acute serum amyloid A by keratinocytes contributes to psoriasis pathogenesis. PLoS ONE 2017, 12, e0181486. [Google Scholar] [CrossRef]
- Lee, J.Y.; Hall, J.A.; Kroehling, L.; Wu, L.; Najar, T.; Nguyen, H.H.; Lin, W.Y.; Yeung, S.T.; Silva, H.M.; Li, D.; et al. Serum Amyloid A Proteins Induce Pathogenic Th17 Cells and Promote Inflammatory Disease. Cell 2020, 180, 79–91.e16. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iida, S.; Nakanishi, T.; Momose, F.; Ichishi, M.; Mizutani, K.; Matsushima, Y.; Umaoka, A.; Kondo, M.; Habe, K.; Hirokawa, Y.; et al. IL-17A Is the Critical Cytokine for Liver and Spleen Amyloidosis in Inflammatory Skin Disease. Int. J. Mol. Sci. 2022, 23, 5726. https://doi.org/10.3390/ijms23105726
Iida S, Nakanishi T, Momose F, Ichishi M, Mizutani K, Matsushima Y, Umaoka A, Kondo M, Habe K, Hirokawa Y, et al. IL-17A Is the Critical Cytokine for Liver and Spleen Amyloidosis in Inflammatory Skin Disease. International Journal of Molecular Sciences. 2022; 23(10):5726. https://doi.org/10.3390/ijms23105726
Chicago/Turabian StyleIida, Shohei, Takehisa Nakanishi, Fumiyasu Momose, Masako Ichishi, Kento Mizutani, Yoshiaki Matsushima, Ai Umaoka, Makoto Kondo, Koji Habe, Yoshifumi Hirokawa, and et al. 2022. "IL-17A Is the Critical Cytokine for Liver and Spleen Amyloidosis in Inflammatory Skin Disease" International Journal of Molecular Sciences 23, no. 10: 5726. https://doi.org/10.3390/ijms23105726
APA StyleIida, S., Nakanishi, T., Momose, F., Ichishi, M., Mizutani, K., Matsushima, Y., Umaoka, A., Kondo, M., Habe, K., Hirokawa, Y., Watanabe, M., Iwakura, Y., Miyahara, Y., Imai, Y., & Yamanaka, K. (2022). IL-17A Is the Critical Cytokine for Liver and Spleen Amyloidosis in Inflammatory Skin Disease. International Journal of Molecular Sciences, 23(10), 5726. https://doi.org/10.3390/ijms23105726