
Exploration of Multiverse Activities of Endocannabinoids in Biological Systems


 , and
, and 
Abstract
:
1. Introduction
1.1. A Brief History of Endocannabinoid System
1.2. Biochemistry, Production and Secretion of Endocannabinoids
1.3. Endocannabinoids Derived from Eicosapentaenoic and Docosahexaenoic Acids
1.4. Physiological Endocannabinoid-like Molecules
1.5. Uptake and Degradation of Endocannabinoids
1.6. Cannabinoid Receptors
1.7. Endocannabinoids and Their Pharmacology
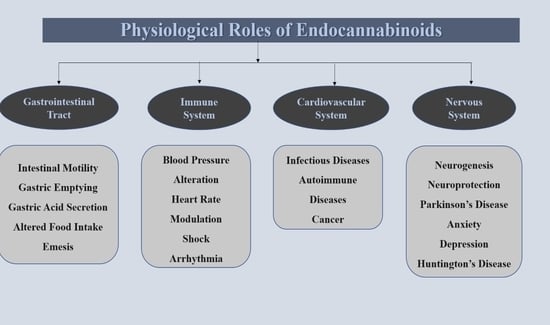
2. Physiological Role of Endocannabinoids in Human Gastrointestinal System
2.1. Motility
2.2. Gastric Emptying and Intestinal Motility
2.3. Secretion of Gastric Acid
2.4. Food Intake Alteration
2.5. Emesis
3. Physiological Role of Endocannabinoids in the Human Cardiovascular System
3.1. Endocannabinoids in Modulating Heart Rate and Blood Pressure
3.2. Endocannabinoids and Hypertension
3.3. Endocannabinoids and Shock
3.4. Endocannabinoids and Arrythmia
4. Physiological Role of Endocannabinoids in Immune System
4.1. Endocannabinoids and Infectious Diseases
4.2. Endocannabinoids and Cancer
4.3. Endocannabinoids and Autoimmune Diseases
5. Physiological Role of Endocannabinoids in Human Brain
5.1. Endocannabinoids and Neurogenesis
5.2. Endocannabinoids and Neuroprotection
5.3. Endocannabinoids and Parkinson’s Disease
5.4. Endocannabinoids and Anxiety
5.5. Endocannabinoids and Depression
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Fonseca, F.R.; del Arco, I.; Bermudez-Silva, F.J.; Bilbao, A.; Cippitelli, A.; Navarro, M. The Endocannabinoid System: Physiology and Pharmacology. Alcohol Alcohol. 2005, 40, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Battista, N.; di Tommaso, M.; Bari, M.; Maccarrone, M. The Endocannabinoid System: An Overview. Front. Behav. Neurosci. 2012, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bari, M.; Battista, N.; Pirazzi, V.; Maccarrone, M. The Manifold Actions of Endocannabinoids on Female and Male Reproductive Events. Front. Biosci. 2011, 16, 498–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beinfeld, M.C.; Connolly, K. Activation of CB1 Cannabinoid Receptors in Rat Hippocampal Slices Inhibits Potassium-Evoked Cholecystokinin Release, a Possible Mechanism Contributing to the Spatial Memory Defects Produced by Cannabinoids. Neurosci. Lett. 2001, 301, 69–71. [Google Scholar] [CrossRef]
- Mouslech, Z.; Valla, V. Endocannabinoid System: An Overview of Its Potential in Current Medical Practice. Neuroendocrinol. Lett. 2009, 30, 153–179. [Google Scholar] [PubMed]
- Pacher, P.; Bátkai, S.; Kunos, G. The Endocannabinoid System as an Emerging Target of Pharmacotherapy. Pharmacol. Rev. 2006, 58, 389–462. [Google Scholar] [CrossRef] [Green Version]
- Gabri, A.C.; Galanti, M.R.; Orsini, N.; Magnusson, C. Changes in Cannabis Policy and Prevalence of Recreational Cannabis Use among Adolescents and Young Adults in Europe—An Interrupted Time-Series Analysis. PLoS ONE 2022, 17, e0261885. [Google Scholar] [CrossRef]
- de Petrocellis, L.; di Marzo, V. An Introduction to the Endocannabinoid System: From the Early to the Latest Concepts. Best Pract. Res. Clin. Endocrinol. Metab. 2009, 23, 1–15. [Google Scholar] [CrossRef]
- Crocq, M.A. History of Cannabis and the Endocannabinoid System. Dialogues Clin. Neurosci. 2020, 22, 223–228. [Google Scholar] [CrossRef]
- di Marzo, V. A Brief History of Cannabinoid and Endocannabinoid Pharmacology as Inspired by the Work of British Scientists. Trends Pharmacol. Sci. 2006, 27, 134–140. [Google Scholar] [CrossRef]
- Piomelli, D. The Endocannabinoid System: A Drug Discovery Perspective. Curr. Opin. Investig. Drugs 2005, 6, 672–679. [Google Scholar] [PubMed]
- Gómez-Ruiz, M.; Hernández, M.; de Miguel, R.; Ramos, J.A. An Overview on the Biochemistry of the Cannabinoid System. Mol. Neurobiol. 2007, 36, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Bisogno, T.; Ligresti, A.; di Marzo, V. The Endocannabinoid Signalling System: Biochemical Aspects. In Pharmacology Biochemistry and Behavior; Elsevier: Milano, Italy, 2005; Volume 81, pp. 224–238. [Google Scholar]
- di Marzo, V. Endocannabinoids: Synthesis and Degradation. Rev. Physiol. Biochem. Pharm. 2006, 160, 1–24. [Google Scholar] [CrossRef]
- Fonseca, B.M.; Costa, M.A.; Almada, M.; Correia-Da-Silva, G.; Teixeira, N.A. Endogenous Cannabinoids Revisited: A Biochemistry Perspective. Prostaglandins Other Lipid Mediat. 2013, 102–103, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.I.; Nicoll, R.A. Endocannabinoid Signaling in the Brain. Science 1979, 2002, 296. [Google Scholar] [CrossRef] [Green Version]
- Giuffrida, A.; Beltramo, M.; Piomelli, D. Mechanisms of Endocannabinoid Inactivation: Biochemistry and Pharmacology. J. Pharmacol. Exp. Ther. 2001, 298, 7–14. [Google Scholar]
- Naughton, S.S.; Mathai, M.L.; Hryciw, D.H.; McAinch, A.J. Fatty Acid Modulation of the Endocannabinoid System and the Effect on Food Intake and Metabolism. Int. J. Endocrinol. 2013, 2013, 361895. [Google Scholar] [CrossRef] [Green Version]
- Watson, J.E.; Kim, J.S.; Das, A. Emerging Class of Omega-3 Fatty Acid Endocannabinoids & Their Derivatives. Prostaglandin. Other Lipid Mediat. 2019, 143, 106337. [Google Scholar]
- Fezza, F.; Bari, M.; Florio, R.; Talamonti, E.; Feole, M.; Maccarrone, M. Endocannabinoids, Related Compounds and Their Metabolic Routes. Molecules 2014, 19, 17078–17106. [Google Scholar] [CrossRef]
- Ahn, K.; McKinney, M.K.; Cravatt, B.F. Enzymatic Pathways That Regulate Endocannabinoid Signaling in the Nervous System. Chem. Rev. 2008, 108, 1687–1707. [Google Scholar] [CrossRef] [Green Version]
- Vaughan, C.W.; Christie, M.J. Retrograde Signalling by Endocannabinoids. Handb. Exp. Pharmacol. 2005, 168, 367–383. [Google Scholar] [CrossRef]
- McIntosh, A.L.; Huang, H.; Landrock, D.; Martin, G.G.; Li, S.; Kier, A.B.; Schroeder, F. Impact of Fabp1 Gene Ablation on Uptake and Degradation of Endocannabinoids in Mouse Hepatocytes. Lipids 2018, 53, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Nicolussi, S.; Gertsch, J. Endocannabinoid Transport Revisited. In Vitamins and Hormones; Elsevier: Amsterdam, The Netherlands, 2015; Volume 98. [Google Scholar]
- Freund, T.F.; Hájos, N. Excitement Reduces Inhibition via Endocannabinoids. Neuron 2003, 38, 362–365. [Google Scholar] [CrossRef] [Green Version]
- Howlett, A.C. The Cannabinoid Receptors. Prostaglandins Other Lipid Mediat. 2002, 68–69, 619–631. [Google Scholar] [CrossRef]
- Mackie, K. Cannabinoid Receptors: Where They Are and What They Do. J. Neuroendocrinol. 2008, 20, 10–14. [Google Scholar] [CrossRef]
- Starowicz, K.; Malek, N.; Przewlocka, B. Cannabinoid Receptors and Pain. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2013, 2, 121–132. [Google Scholar] [CrossRef]
- Le Fur, G.; Rinaldi-Carmona, M.; Barth, F.; Alexander, J. Pharmacology of Cannabinoid Receptors. Bull. L’academie Natl. Med. 2007, 191, 933–940. [Google Scholar] [CrossRef]
- Lu, H.C.; MacKie, K. An Introduction to the Endogenous Cannabinoid System. Biol. Psychiatry 2016, 79, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Di Marzo, V.; Melck, D.; Bisogno, T.; de Petrocellis, L. Endocannabinoids: Endogenous Cannabinoid Receptor Ligands with Neuromodulatory Action. Trends Neurosci. 1998, 21, 80. [Google Scholar] [CrossRef]
- Sugiura, T.; Kondo, S.; Sukagawa, A.; Nakane, S.; Shinoda, A.; Itoh, K.; Yamashita, A.; Waku, K. 2-Arachidonoylglycerol: A Possible Endogenous Cannabinoid Receptor Ligand in Brain. Biochem. Biophys. Res. Commun. 1995, 215, 89–97. [Google Scholar] [CrossRef]
- Howlett, A.C. Cannabinoid Receptor Signaling. Handb. Exp. Pharmacol. 2005, 168, 53–79. [Google Scholar] [CrossRef]
- Mackie, K. Cannabinoid Receptors as Therapeutic Targets. Annu. Rev. Pharmacol. Toxicol. 2006, 46, 101–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez De Fonseca, F.; del Arco, I.; Martín-Calderón, J.L.; Gorriti, M.A.; Navarro, M. Role of the Endogenous Cannabinoid System in the Regulation of Motor Activity. Neurobiol. Dis. 1998, 5, 483–501. [Google Scholar] [CrossRef] [Green Version]
- Merritt, L.L.; Martin, B.R.; Walters, C.; Lichtman, A.H.; Damaj, M.I. The Endogenous Cannabinoid System Modulates Nicotine Reward and Dependence. J. Pharmacol. Exp. Ther. 2008, 326, 483–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piomelli, D.; Giuffrida, A.; Calignano, A.; Rodríguez De Fonseca, F. The Endocannabinoid System as a Target for Therapeutic Drugs. Trends Pharmacol. Sci. 2000, 21, 218–242. [Google Scholar] [CrossRef] [Green Version]
- Pertwee, R.G. Cannabinoid Pharmacology: The First 66 Years. Br. J. Pharmacol. 2006, 147, S163–S171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Moreno, J.A.; González-Cuevas, G.; Moreno, G.; Navarro, M. The Pharmacology of the Endocannabinoid System: Functional and Structural Interactions with Other Neurotransmitter Systems and Their Repercussions in Behavioral Addiction. Addict. Biol. 2008, 13, 160–187. [Google Scholar] [CrossRef]
- Romero, J.; Lastres-Becker, I.; de Miguel, R.; Berrendero, F.; Ramos, J.A.; Fernández-Ruiz, J. The Endogenous Cannabinoid System and the Basal Ganglia: Biochemical, Pharmacological, and Therapeutic Aspects. Pharmacol. Ther. 2002, 95, 137–152. [Google Scholar] [CrossRef]
- Craft, R.M.; Marusich, J.A.; Wiley, J.L. Sex Differences in Cannabinoid Pharmacology: A Reflection of Differences in the Endocannabinoid System? Life Sci. 2013, 92, 476–481. [Google Scholar] [CrossRef] [Green Version]
- Urquhart, P.; Nicolaou, A.; Woodward, D.F. Endocannabinoids and Their Oxygenation by Cyclo-Oxygenases, Lipoxygenases and Other Oxygenases. Biochim. Et Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 366–376. [Google Scholar] [CrossRef]
- Sanger, G.J. Endocannabinoids and the Gastrointestinal Tract: What Are the Key Questions? Br. J. Pharmacol. 2007, 152, 663–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Jo, J.; Chung, H.Y.; Pothoulakis, C.; Im, E. Endocannabinoids in the Gastrointestinal Tract. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G655–G666. [Google Scholar] [CrossRef] [PubMed]
- Massa, F.; Storr, M.; Lutz, B. The Endocannabinoid System in the Physiology and Pathophysiology of the Gastrointestinal Tract. J. Mol. Med. 2005, 83, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Dipatrizio, N.V. Endocannabinoids in the Gut. Cannabis Cannabinoid Res. 2016, 1, 67–77. [Google Scholar] [CrossRef]
- Pinto, L.; Capasso, R.; di Carlo, G.; Izzo, A.A. Endocannabinoids and the Gut. Prostaglandins Leukot. Essent. Fat. Acids 2002, 66, 333–341. [Google Scholar] [CrossRef]
- Izzo, A.A.; Camilleri, M. Emerging Role of Cannabinoids in Gastrointestinal and Liver Diseases: Basic and Clinical Aspects. Gut 2008, 57, 1140–1155. [Google Scholar] [CrossRef]
- Camilleri, M.; Carlson, P.; McKinzie, S.; Grudell, A.; Busciglio, I.; Burton, D.; Baxter, K.; Ryks, M.; Zinsmeister, A.R. Genetic Variation in Endocannabinoid Metabolism, Gastrointestinal Motility, and Sensation. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 294, G13–G19. [Google Scholar] [CrossRef]
- Aviello, G.; Romano, B.; Izzo, A.A. Cannabinoids and Gastrointestinal Motility: Animal and Human Studies. Eur. Rev. Med. Pharmacol. Sci. 2008, 12, 81–93. [Google Scholar]
- Di Marzo, V.; Capasso, R.; Matias, I.; Aviello, G.; Petrosino, S.; Borrelli, F.; Romano, B.; Orlando, P.; Capasso, F.; Izzo, A.A. The Role of Endocannabinoids in the Regulation of Gastric Emptying: Alterations in Mice Fed a High-Fat Diet. Br. J. Pharmacol. 2008, 153, 1272–7280. [Google Scholar] [CrossRef] [Green Version]
- Hillard, C.J. Biochemistry and Pharmacology of the Endocannabinoids Arachidonylethanolamide and 2-Arachidonylglycerol. Prostaglandin. Other Lipid Mediat. 2000, 61, 3–18. [Google Scholar] [CrossRef]
- Rahman, S.M.K.; Uyama, T.; Hussain, Z.; Ueda, N. Roles of Endocannabinoids and Endocannabinoid-Like Molecules in Energy Homeostasis and Metabolic Regulation: A Nutritional Perspective. Annu. Rev. Nutr. 2021, 41, 177–202. [Google Scholar] [CrossRef] [PubMed]
- Gyires, K.; Zádori, Z.S. Role of Cannabinoids in Gastrointestinal Mucosal Defense and Inflammation. Curr. Neuropharmacol. 2016, 14, 935–951. [Google Scholar] [CrossRef] [PubMed]
- Rajaraman, G.; Simcocks, A.; Hryciw, D.H.; Hutchinson, D.S.; McAinch, A.J. G Protein Coupled Receptor 18: A Potential Role for Endocannabinoid Signaling in Metabolic Dysfunction. Mol. Nutr. Food Res. 2016, 60, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Quarta, C.; Mazza, R.; Obici, S.; Pasquali, R.; Pagotto, U. Energy Balance Regulation by Endocannabinoids at Central and Peripheral Levels. Trends Mol. Med. 2011, 17, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Bisogno, T.; Maccarrone, M. Endocannabinoid Signaling and Its Regulation by Nutrients. BioFactors 2014, 40, 373–380. [Google Scholar] [CrossRef]
- Alizadeh, A.; Zendehdel, M.; Babapour, V.; Charkhkar, S.; Hassanpour, S. Role of Cannabinoidergic System on Food Intake in Neonatal Layer-Type Chicken. Vet. Res. Commun. 2015, 39, 151–157. [Google Scholar] [CrossRef]
- Sharkey, K.A.; Darmani, N.A.; Parker, L.A. Regulation of Nausea and Vomiting by Cannabinoids and the Endocannabinoid System. Eur. J. Pharmacol. 2014, 722, 134–136. [Google Scholar] [CrossRef] [Green Version]
- Malik, Z.; Baik, D.; Schey, R. The Role of Cannabinoids in Regulation of Nausea and Vomiting, and Visceral Pain. Curr. Gastroenterol. Rep. 2015, 17, 429. [Google Scholar] [CrossRef]
- Van Sickle, M.D.; Oland, L.D.; Ho, W.; Hillard, C.J.; Mackie, K.; Davison, J.S.; Sharkey, K.A. Cannabinoids Inhibit Emesis through CB1 Receptors in the Brainstem of the Ferret. Gastroenterology 2001, 121, 767–774. [Google Scholar] [CrossRef]
- Whiting, P.F.; Wolff, R.F.; Deshpande, S.; di Nisio, M.; Duffy, S.; Hernandez, A.v.; Keurentjes, J.C.; Lang, S.; Misso, K.; Ryder, S.; et al. Cannabinoids for Medical Use: A Systematic Review and Meta-Analysis. JAMA 2015, 313, 2456–2473. [Google Scholar] [CrossRef]
- Sierra, S.; Luquin, N.; Navarro-Otano, J. The Endocannabinoid System in Cardiovascular Function: Novel Insights and Clinical Implications. Clin. Auton. Res. 2018, 28, 35–52. [Google Scholar] [CrossRef]
- O’Sullivan, S.E. Endocannabinoids and the Cardiovascular System in Health and Disease. In Endocannabinoids; Springer: Berlin, Germany, 2015. [Google Scholar]
- Montecucco, F.; di Marzo, V. At the Heart of the Matter: The Endocannabinoid System in Cardiovascular Function and Dysfunction. Trends Pharm. Sci. 2012, 33, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Zubrzycki, M.; Liebold, A.; Janecka, A.; Zubrzycka, M. A New Face of Endocannabinoids in Pharmacotherapy. Part I: Protective Role of Endocannabinoids in Hypertension and Myocardial Infarction. J. Physiol. Pharmacol. 2014, 65, 171–181. [Google Scholar] [PubMed]
- Ros, J.; Clària, J.; To-Figueras, J.; Planagumà, A.; Cejudo-Martín, P.; Fernández-Varo, G.; Martín-Ruiz, R.; Arroyo, V.; Rivera, F.; Rodüs, J.; et al. Endogenous Cannabinoids: A New System Involved in the Homeostasis of Arterial Pressure in Experimental Cirrhosis in the Rat. Gastroenterology 2002, 122, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Kunos, G.; Járai, Z.; Bátkai, S.; Goparaju, S.K.; Ishac, E.J.N.; Liu, J.; Wang, L.; Wagner, J.A. Endocannabinoids as Cardiovascular Modulators. Chem. Phys. Lipids 2000, 108, 159–168. [Google Scholar] [CrossRef]
- Mourtakos, S.; Vassiliou, G.; Kontoangelos, K.; Philippou, A.; Tzavellas, E.; Tornero-Aguilera, J.F.; Clemente-Suárez, V.J.; Papageorgiou, C.; Sidossis, L.S.; Papageorgiou, C. Endocannabinoids and Heart Rate Variability Alterations after Exposure to Prolonged Intensive Physical Exercise of the Hellenic Navy Seals. Int. J. Environ. Res. Public Health 2022, 19, 28. [Google Scholar] [CrossRef]
- Jones, R.T. Cardiovascular System Effects of Marijuana. J. Clin. Pharmacol. 2002, 42, 48S–63S. [Google Scholar] [CrossRef]
- Bátkai, S.; Pacher, P.; Osei-Hyiaman, D.; Radaeva, S.; Liu, J.; Harvey-White, J.; Offertáler, L.; Mackie, K.; Rudd, M.A.; Bukoski, R.D.; et al. Endocannabinoids Acting at Cannabinoid-1 Receptors Regulate Cardiovascular Function in Hypertension. Circulation 2004, 110, 1996–2002. [Google Scholar] [CrossRef] [Green Version]
- Szekeres, M.; Nádasy, G.L.; Turu, G.; Soltész-Katona, E.; Benyó, Z.; Offermanns, S.; Ruisanchez, É.; Szabó, E.; Takáts, Z.; Bátkai, S.; et al. Endocannabinoid-Mediated Modulation of Gq/11 Protein-Coupled Receptor Signaling-Induced Vasoconstriction and Hypertension. Mol. Cell. Endocrinol. 2015, 403, 46–56. [Google Scholar] [CrossRef]
- Cunha, P.; Romão, A.M.; Mascarenhas-Melo, F.; Teixeira, H.M.; Reis, F. Endocannabinoid System in Cardiovascular Disorders-New Pharmacotherapeutic Opportunities. J. Pharm. Bioallied Sci. 2011, 3, 350–360. [Google Scholar]
- Malinowska, B.; Lupinski, S.; Godlewski, G.; Baranowska, U.; Schlicker, E. Role of Endocannabinoids in Cardiovascular Shock. J. Physiol. Pharmacol. 2008, 59, 91–107. [Google Scholar]
- Hiley, C.R.; Ford, W.R. Endocannabinoids as Mediators in the Heart: A Potential Target for Therapy of Remodelling after Myocardial Infarction? Br. J. Pharmacol. 2003, 138, 1183–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozturk, H.M.; Yetkin, E.; Ozturk, S. Synthetic Cannabinoids and Cardiac Arrhythmia Risk: Review of the Literature. Cardiovasc. Toxicol. 2019, 19, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Durst, R.; Lotan, C. The Potential for Clinical Use of Cannabinoids in Treatment of Cardiovascular Diseases. Cardiovasc. Ther. 2011, 29, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Rahaman, O.; Ganguly, D. Endocannabinoids in Immune Regulation and Immunopathologies. Immunology 2021, 164, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Almogi-Hazan, O.; Or, R. Cannabis, the Endocannabinoid System and Immunity—the Journey from the Bedside to the Bench and Back. Int. J. Mol. Sci. 2020, 21, 4448. [Google Scholar] [CrossRef]
- Salzet, M.; Breton, C.; Bisogno, T.; di Marzo, V. Comparative Biology of the Endocannabinoid System: Possible Role in the Immune Response. Eur. J. Biochem. 2000, 267, 4917–4927. [Google Scholar] [CrossRef]
- Tanasescu, R.; Constantinescu, C.S. Cannabinoids and the Immune System: An Overview. Immunobiology 2010, 215, 588–597. [Google Scholar] [CrossRef]
- Van Breemen, R.B.; Muchiri, R.N.; Bates, T.A.; Weinstein, J.B.; Leier, H.C.; Farley, S.; Tafesse, F.G. Cannabinoids Block Cellular Entry of SARS-CoV-2 and the Emerging Variants. J. Nat. Prod. 2022, 85, 176–184. [Google Scholar] [CrossRef]
- Hernández-Cervantes, R.; Méndez-DÍaz, M.; Prospéro-García, Ó.; Morales-Montor, J. Immunoregulatory Role of Cannabinoids during Infectious Disease. NeuroImmunoModulation 2018, 24, 183–199. [Google Scholar] [CrossRef]
- Wang, B.; Li, D.; Fiselier, A.; Kovalchuk, I.; Kovalchuk, O. New AKT-Dependent Mechanisms of Anti-COVID-19 Action of High-CBD Cannabis Sativa Extracts. Cell Death Discov. 2022, 8, 110. [Google Scholar] [CrossRef]
- Croxford, J.L.; Yamamura, T. Cannabinoids and the Immune System: Potential for the Treatment of Inflammatory Diseases? J. Neuroimmunol. 2005, 166, 3–18. [Google Scholar] [CrossRef]
- Hermanson, D.J.; Marnett, L.J. Cannabinoids, Endocannabinoids, and Cancer. Cancer Metastasis Rev. 2011, 30, 599–612. [Google Scholar] [CrossRef]
- De Petrocellis, L.; Melck, D.; Bisogno, T.; di Marzo, V. Endocannabinoids and Fatty Acid Amides in Cancer, Inflammation and Related Disorders. Chem. Phys. Lipids 2000, 108, 191–209. [Google Scholar] [CrossRef]
- Velasco, G.; Sánchez, C.; Guzmán, M. Endocannabinoids and Cancer. Endocannabinoids 2015, 449–472. [Google Scholar] [CrossRef]
- Moreno, E.; Cavic, M.; Krivokuca, A.; Casadó, V.; Canela, E. The Endocannabinoid System as a Target in Cancer Diseases: Are We There Yet? Front. Pharmacol. 2019, 10, 339. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.; Pearson, R.G.; Kurian, N.; Latif, M.L.; Garle, M.J.; Barrett, D.A.; Kendall, D.A.; Scammell, B.E.; Reeve, A.J.; Chapman, V. Characterisation of the Cannabinoid Receptor System in Synovial Tissue and Fluid in Patients with Osteoarthritis and Rheumatoid Arthritis. Arthritis Res. Ther. 2008, 10, R43. [Google Scholar] [CrossRef] [Green Version]
- Katchan, V.; David, P.; Shoenfeld, Y. Cannabinoids and Autoimmune Diseases: A Systematic Review. Autoimmun. Rev. 2016, 15, 513–528. [Google Scholar] [CrossRef]
- Sido, J.M.; Nagarkatti, P.S.; Nagarkatti, M. Role of Endocannabinoid Activation of Peripheral CB1 Receptors in the Regulation of Autoimmune Disease. Int. Rev. Immunol. 2015, 34, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Hill, K.P.; Gold, M.S.; Nemeroff, C.B.; McDonald, W.; Grzenda, A.; Widge, A.S.; Rodriguez, C.; Kraguljac, N.v.; Krystal, J.H.; Carpenter, L.L. Risks and Benefits of Cannabis and Cannabinoids in Psychiatry. Am. J. Psychiatry 2022, 179, 98–109. [Google Scholar] [CrossRef]
- Prenderville, J.A.; Kelly, Á.M.; Downer, E.J. The Role of Cannabinoids in Adult Neurogenesis. Br. J. Pharmacol. 2015, 172, 3950–3963. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Vasilyev, D.V.; Goncalves, M.B.; Howell, F.V.; Hobbs, C.; Reisenberg, M.; Shen, R.; Zhang, M.Y.; Strassle, B.W.; Lu, P.; et al. Loss of Retrograde Endocannabinoid Signaling and Reduced Adult Neurogenesis in Diacylglycerol Lipase Knock-out Mice. J. Neurosci. 2010, 30, 2017–2024. [Google Scholar] [CrossRef]
- Oddi, S.; Scipioni, L.; Maccarrone, M. Endocannabinoid System and Adult Neurogenesis: A Focused Review. Curr. Opin. Pharmacol. 2020, 50, 25–32. [Google Scholar] [CrossRef]
- Molina-Holgado, E.; Molina-Holgado, F. Mending the Broken Brain: Neuroimmune Interactions in Neurogenesis: Review. J. Neurochem. 2010, 114, 1277–1290. [Google Scholar]
- Mechoulam, R.; Spatz, M.; Shohami, E. Endocannabinoids and Neuroprotection. Sci. STKE 2002, 2002, re5. [Google Scholar] [CrossRef]
- Xu, J.Y.; Chen, C. Endocannabinoids in Synaptic Plasticity and Neuroprotection. Neuroscientist 2015, 21, 152–168. [Google Scholar] [CrossRef] [Green Version]
- Paloczi, J.; Varga, Z.v.; Hasko, G.; Pacher, P. Neuroprotection in Oxidative Stress-Related Neurodegenerative Diseases: Role of Endocannabinoid System Modulation. Antioxid. Redox Signal. 2018, 29, 75–108. [Google Scholar] [CrossRef]
- Sarne, Y.; Mechoulam, R. Cannabinoids: Between Neuroprotection and Neurotoxicity. Curr. Drug Targets CNS Neurol. Disord. 2005, 4, 677–684. [Google Scholar] [CrossRef]
- Micale, V.; Mazzola, C.; Drago, F. Endocannabinoids and Neurodegenerative Diseases. Pharmacol. Res. 2007, 56, 382–392. [Google Scholar] [CrossRef]
- Stampanoni Bassi, M.; Sancesario, A.; Morace, R.; Centonze, D.; Iezzi, E. Cannabinoids in Parkinson’s Disease. Cannabis Cannabinoid Res. 2017, 2, 21–29. [Google Scholar] [CrossRef]
- Di Filippo, M.; Picconi, B.; Tozzi, A.; Ghiglieri, V.; Rossi, A.; Calabresi, P. The Endocannabinoid System in Parkinsons Disease. Curr. Pharm. Des. 2008, 14, 2337–2346. [Google Scholar] [CrossRef] [PubMed]
- Behl, T.; Kaur, G.; Bungau, S.; Jhanji, R.; Kumar, A.; Mehta, V.; Zengin, G.; Brata, R.; Hassan, S.S.U.; Fratila, O. Distinctive Evidence Involved in the Role of Endocannabinoid Signalling in Parkinson’s Disease: A Perspective on Associated Therapeutic Interventions. Int. J. Mol. Sci. 2020, 21, 6235. [Google Scholar] [CrossRef] [PubMed]
- Shonesy, B.C.; Bluett, R.J.; Ramikie, T.S.; Báldi, R.; Hermanson, D.J.; Kingsley, P.J.; Marnett, L.J.; Winder, D.G.; Colbran, R.J.; Patel, S. Genetic Disruption of 2-Arachidonoylglycerol Synthesis Reveals a Key Role for Endocannabinoid Signaling in Anxiety Modulation. Cell Rep. 2014, 9, 1644–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viveros, M.P.; Marco, E.M.; File, S.E. Endocannabinoid System and Stress and Anxiety Responses. Pharmacol. Biochem. Behav. 2005, 81, 331–342. [Google Scholar] [CrossRef]
- Rana, T.; Behl, T.; Sehgal, A.; Mehta, V.; Singh, S.; Kumar, R.; Bungau, S. Integrating Endocannabinoid Signalling In Depression. J. Mol. Neurosci. 2021, 71, 2022–2034. [Google Scholar] [CrossRef]
- Gallego-Landin, I.; García-Baos, A.; Castro-Zavala, A.; Valverde, O. Reviewing the Role of the Endocannabinoid System in the Pathophysiology of Depression. Front. Pharmacol. 2021, 12, 762738. [Google Scholar] [CrossRef]
- Sharafi, A.; Pakkhesal, S.; Fakhari, A.; Khajehnasiri, N.; Ahmadalipour, A. Rapid Treatments for Depression: Endocannabinoid System as a Therapeutic Target. Neurosci. Biobehav. Rev. 2022, 137, 104635. [Google Scholar] [CrossRef]
- Kreitzer, A.C.; Malenka, R.C. Dopamine Modulation of State-Dependent Endocannabinoid Release and Long-Term Depression in the Striatum. J. Neurosci. 2005, 25, 10537–10545. [Google Scholar] [CrossRef]
- Scherma, M.; Lisa Muntoni, A.; Riedel, G.; Fratta, W.; Fadda, P. Cannabinoids and Their Therapeutic Applications in Mental Disorders. Dialogues Clin. Neurosci. 2020, 22, 271–279. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Name of Compound | Activity/Mechanism of Action |
|---|---|---|
| 1. | HU-210 | CB1 Receptor Agonist |
| 2. | Δ9-THC | Partial Agonist of CB1 and CB2 Receptors |
| 3. | HU-308 | Selective CB2 Receptor |
| 4. | CP-55940 | Potent and Complete Agonist of CB1 and CB2 Receptors |
| 5. | R-(+)-WIN-55, 2/2-2 | Complete CB1 and CB2 Agonist; Affinity Higher towards CB2 Receptor |
| 6. | Anandamide | Partial CB1 and CB2 Agonist |
| 7. | 2-AG | Complete CB1 and CB2 Agonist |
| 8. | Arachidonyl-2′-chloroethylamide (ACEA) | CB1 Agonist |
| 9. | SR 141716A | CB1 Receptor Antagonist |
| 10. | SR 144528 | CB2 Receptor Antagonist |
| 11. | LY320135 | CB1 Receptor Antagonist |
| 12. | AM 630 | CB2 Receptor Antagonist; Low Affinity CB1 Partial Agonist |
| 13. | LH-21 | CB1 Receptor Antagonist |
| 14. | AM 404 | Transport Inhibitor |
| 15. | VCM 707 | Transport Inhibitor |
| 16. | AM 1172 | FAAH-Resistant Transport Inhibitor |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Behl, T.; Makkar, R.; Sehgal, A.; Singh, S.; Makeen, H.A.; Albratty, M.; Alhazmi, H.A.; Meraya, A.M.; Bungau, S. Exploration of Multiverse Activities of Endocannabinoids in Biological Systems. Int. J. Mol. Sci. 2022, 23, 5734. https://doi.org/10.3390/ijms23105734
Behl T, Makkar R, Sehgal A, Singh S, Makeen HA, Albratty M, Alhazmi HA, Meraya AM, Bungau S. Exploration of Multiverse Activities of Endocannabinoids in Biological Systems. International Journal of Molecular Sciences. 2022; 23(10):5734. https://doi.org/10.3390/ijms23105734
Chicago/Turabian StyleBehl, Tapan, Rashita Makkar, Aayush Sehgal, Sukhbir Singh, Hafiz A. Makeen, Mohammed Albratty, Hassan A. Alhazmi, Abdulkarim M. Meraya, and Simona Bungau. 2022. "Exploration of Multiverse Activities of Endocannabinoids in Biological Systems" International Journal of Molecular Sciences 23, no. 10: 5734. https://doi.org/10.3390/ijms23105734
APA StyleBehl, T., Makkar, R., Sehgal, A., Singh, S., Makeen, H. A., Albratty, M., Alhazmi, H. A., Meraya, A. M., & Bungau, S. (2022). Exploration of Multiverse Activities of Endocannabinoids in Biological Systems. International Journal of Molecular Sciences, 23(10), 5734. https://doi.org/10.3390/ijms23105734