
Intermittent Hypoxia Increased the Expression of DBH and PNMT in Neuroblastoma Cells via MicroRNA-375-Mediated Mechanism

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
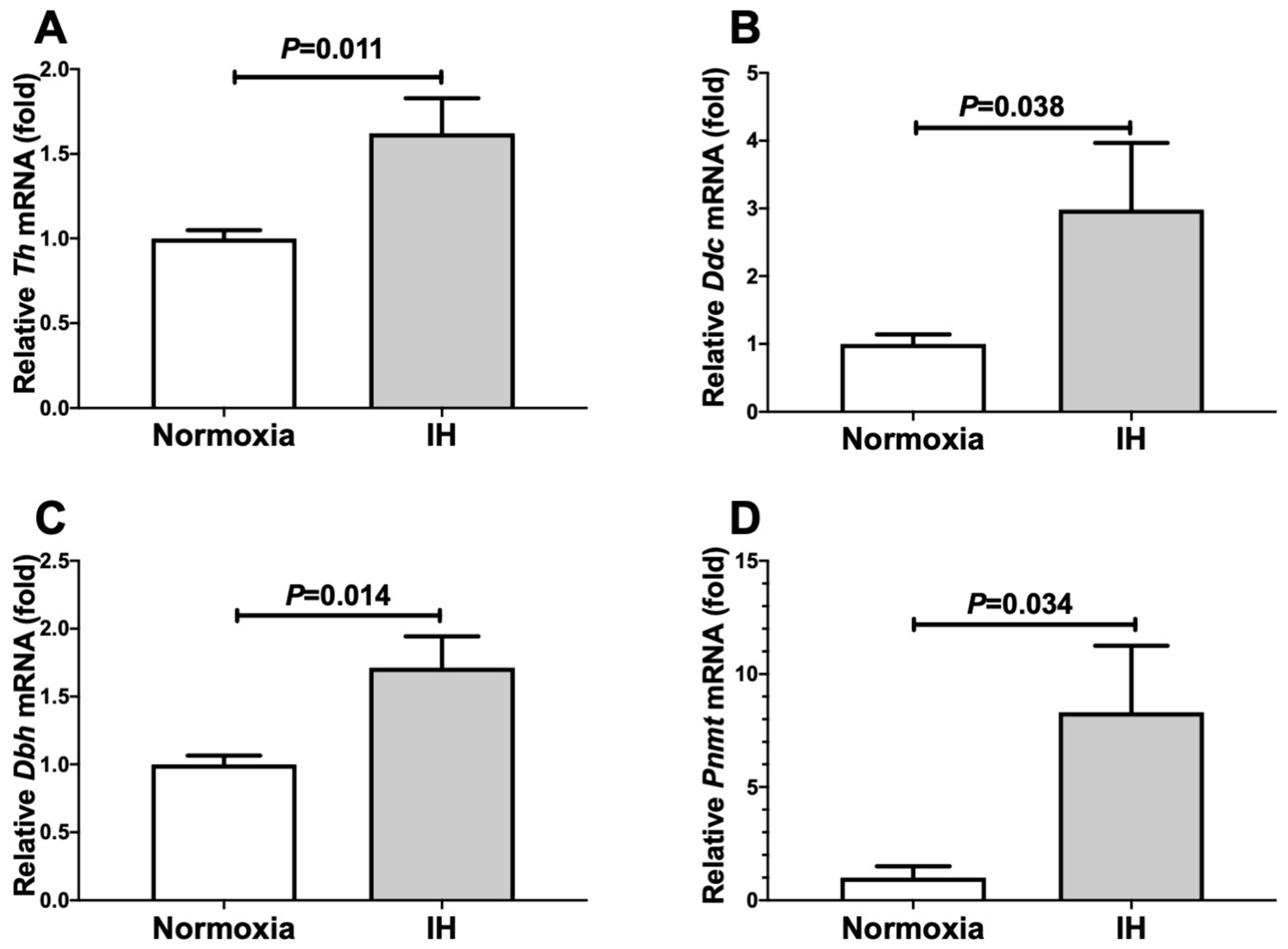
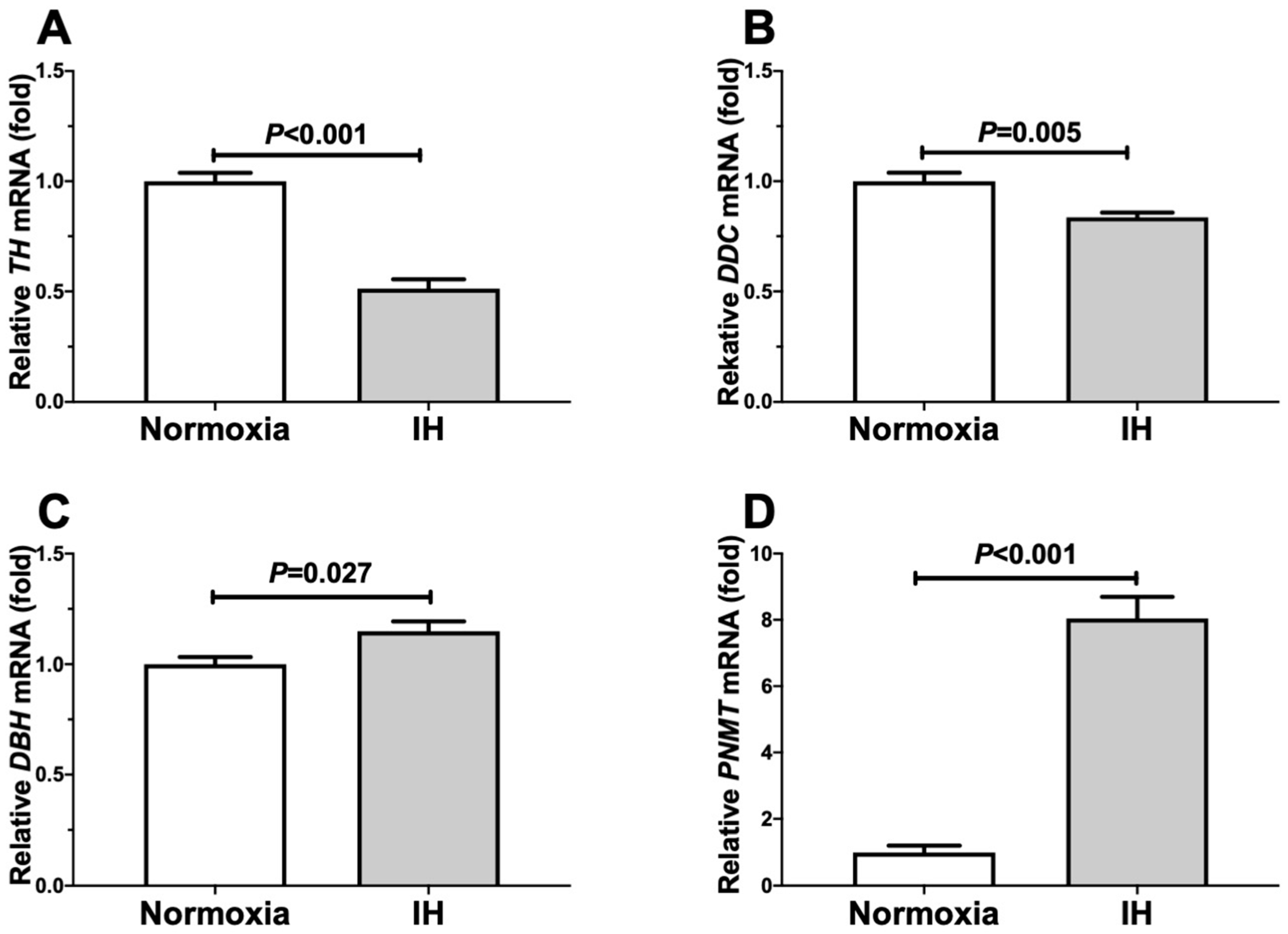
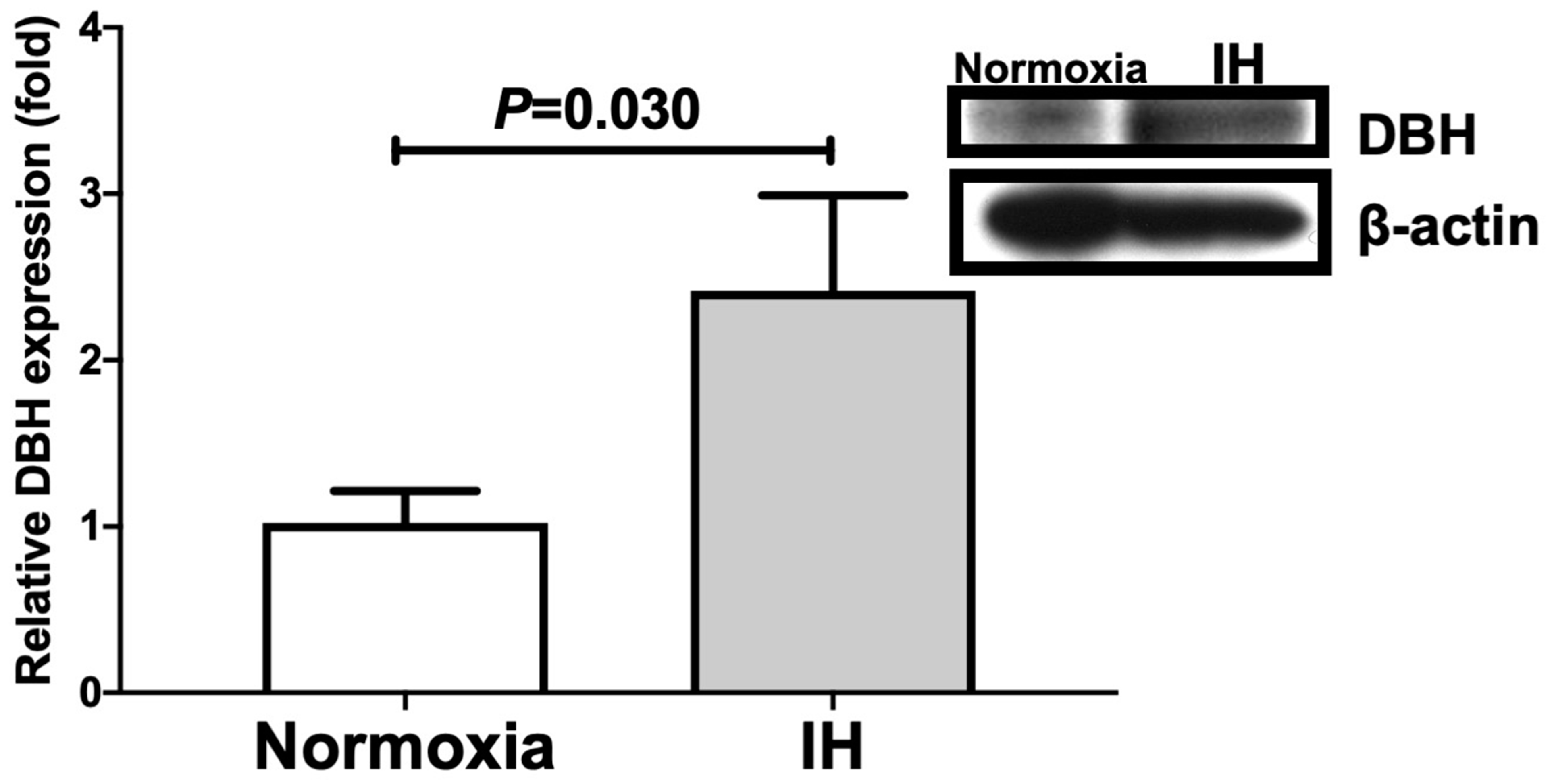
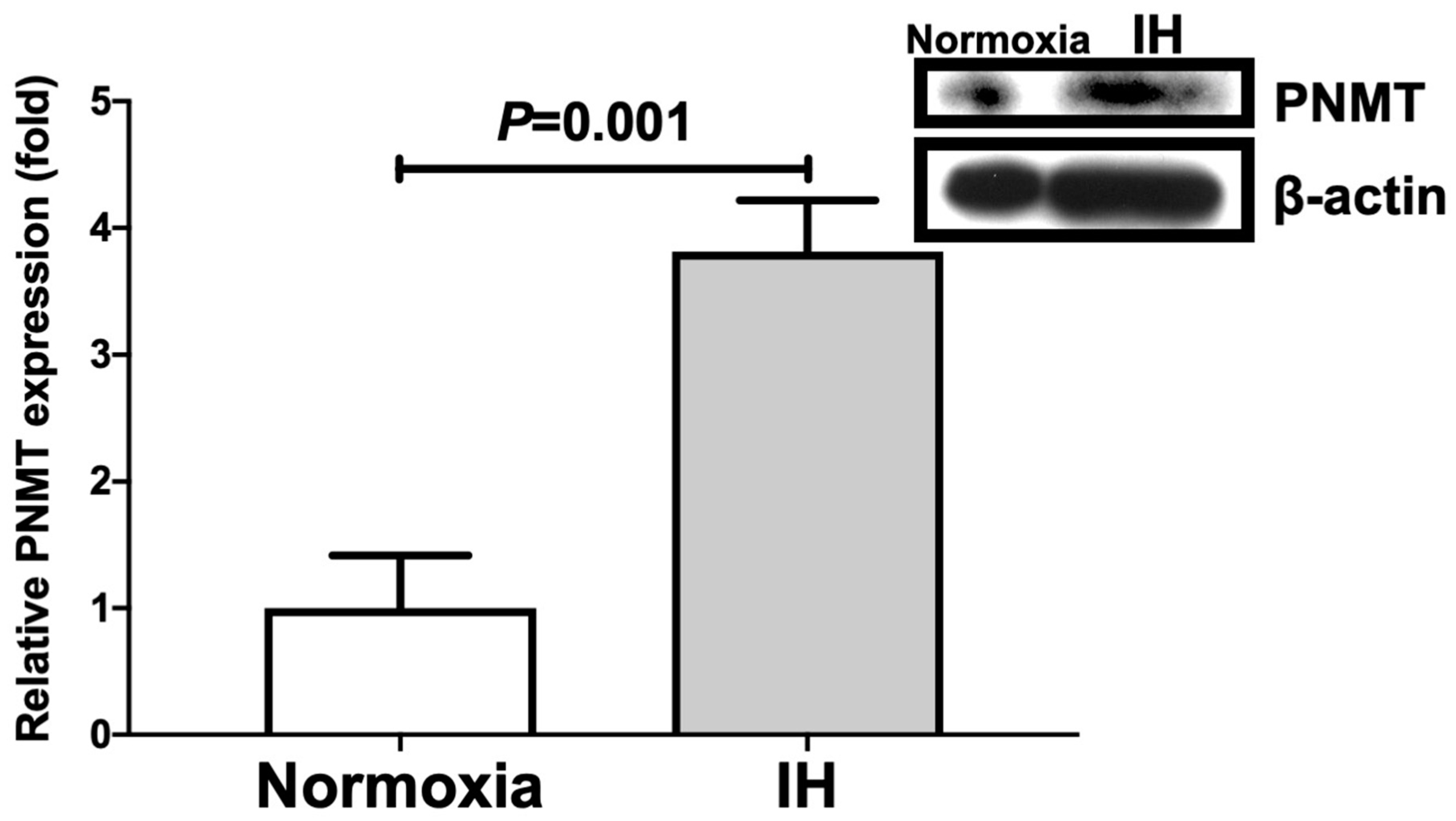
2.1. Gene Expression Levels of DBH and PNMT in Human and Mouse Neuroblastoma Cells Were Increased by IH
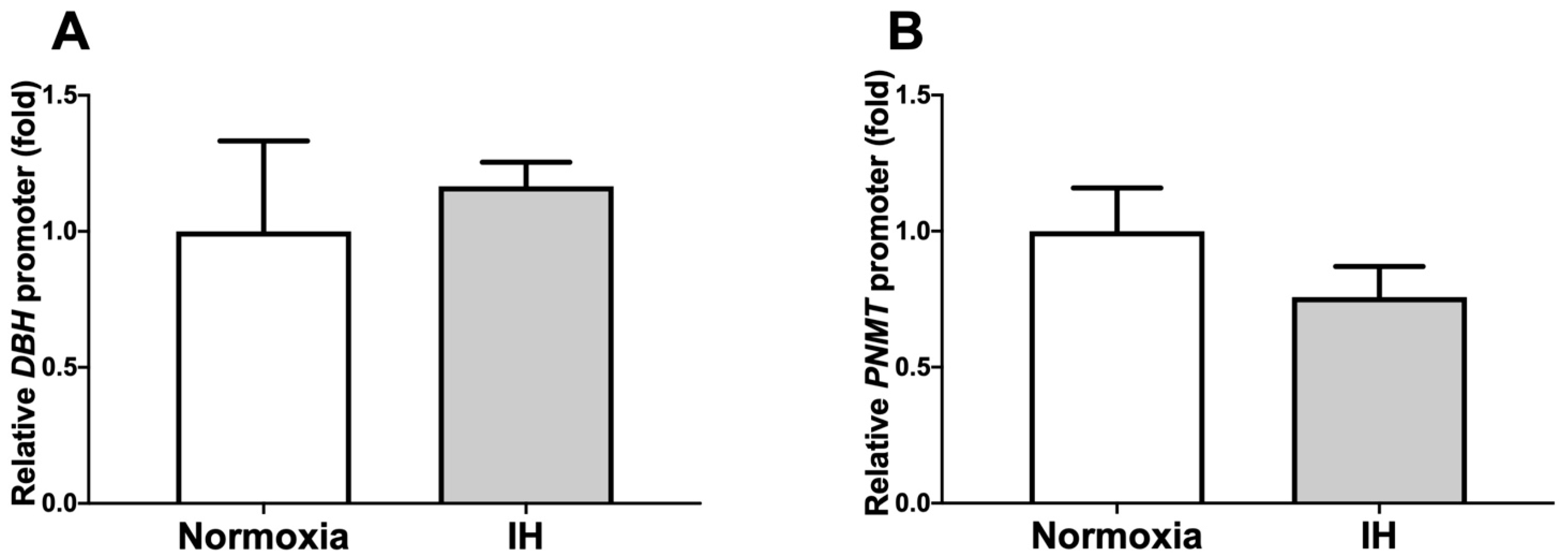
2.2. The Promoter Activities of DBH and PNMT Were Not Increased by IH
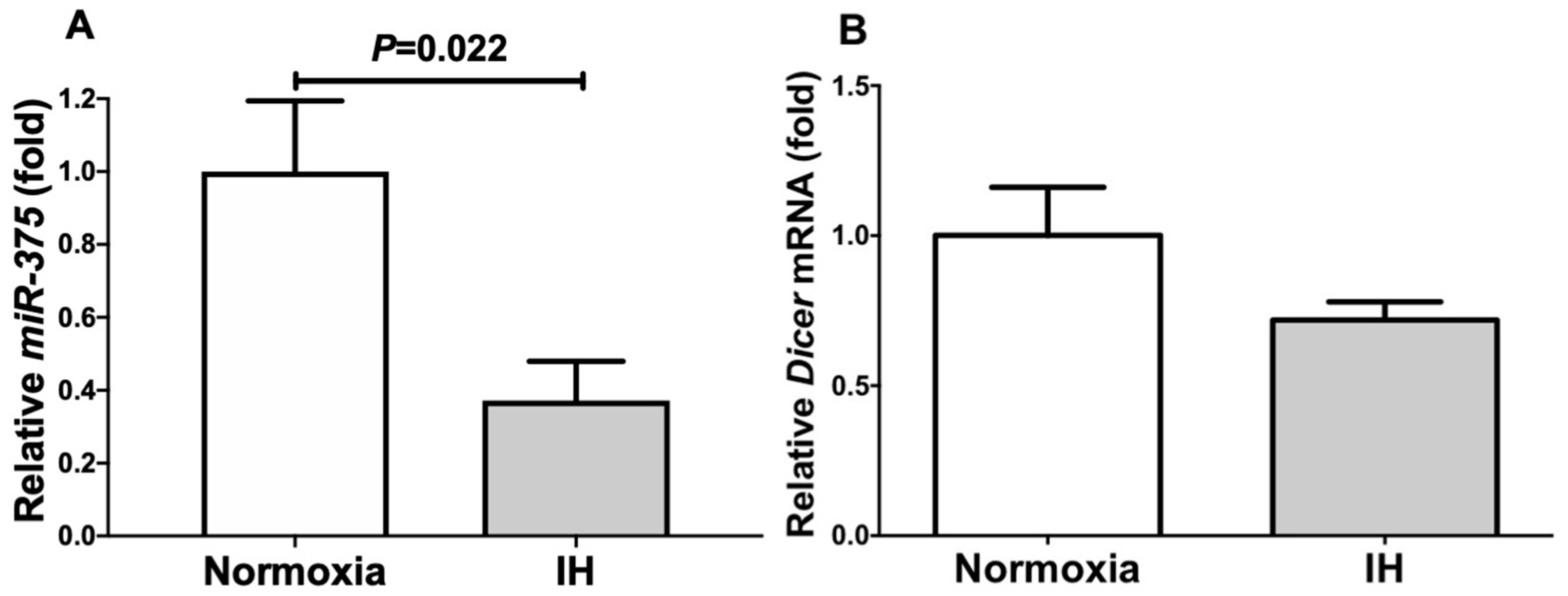
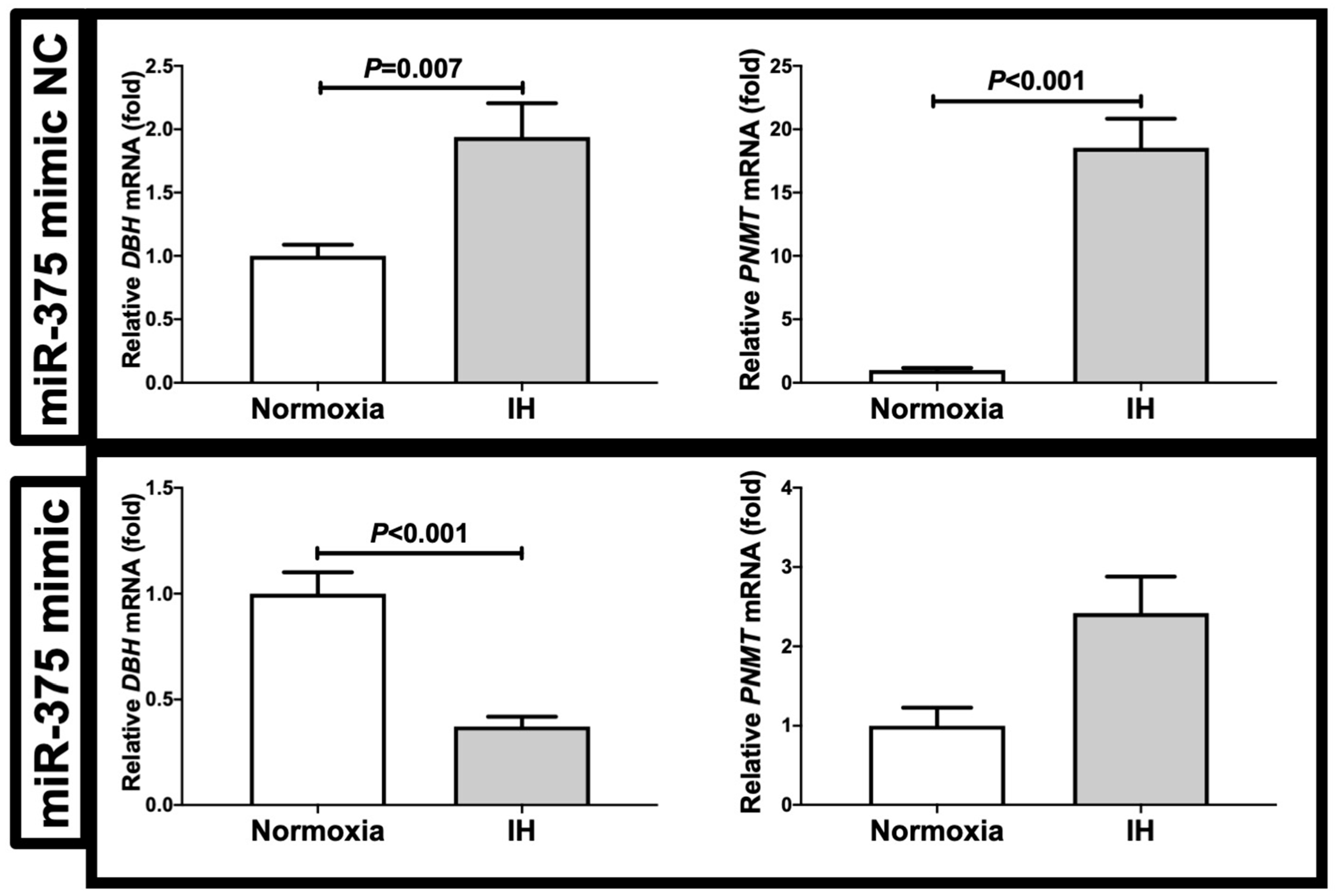
2.3. The MiR-375 Level Was Significantly Decreased by IH
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. RT-PCR
4.3. Construction of Reporter Plasmid and Luciferase Assay
4.4. Immunoblot Analysis
4.5. MiRNA Extraction, Reverse Transcription, and Real-Time Quantitative PCR
4.6. MiR-375 Mimic Transfection
4.7. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DBH | Dopamine β-hydroxylase |
| DDC | DOPA decarboxylase |
| DOPA | L-3,4-dihydroxyphenylalanine |
| Dicer | Endoribonuclease Dicer |
| FCS | Fetal calf serum |
| IH | Intermittent hypoxia |
| miRNA | MicroRNA |
| PNMT | Phenylethanolamine N-methyltransferase |
| Rig | Rat insulinoma gene |
| RpS15 | Ribosomal protein S15 |
| SAS | Sleep apnea syndrome |
| SHRs | Spontaneously hypertensive rats |
| TH | Tyrosine hydroxylase |
References
- Benjafield, A.V.; Ayas, N.T.; Eastwood, P.R.; Heinzer, R.; Ip, M.S.M.; Morrell, M.J.; Nunez, C.M.; Patel, S.R.; Penzel, T.; Pépin, J.-L.; et al. Estimation of the global prevalence and burden of obstructive sleep apnoes: A literature-based analysis. Lancet Respir. Med. 2019, 7, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, M.; Makati, D.; Akbar, S. Review of and updates on hypertension in obstructive sleep apnea. Int. J. Hypertens. 2017, 2017, 1848375. [Google Scholar] [CrossRef] [PubMed]
- Gislason, T.; Aberg, H.; Taube, A. Snoring and systemic hypertension—an epidemiological study. Acta Med. Scand. 1987, 222, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Marrone, O.; Bonsignore, M.R.; Fricano, L.; Lo Coco, R.; Cerasola, G.; Bonsignore, G. Gender and the systemic hypertension-snoring association: A questionnaire-based case-control study. Blood Press 1998, 7, 11–17. [Google Scholar] [CrossRef]
- Takeda, Y.; Itaya-Hironaka, A.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Ota, H.; Kawaguchi, R.; Takasawa, S. Intermittent hypoxia upregulates the renin and Cd38 mRNAs in renin-producing cells via the downregulation of miR-203. Int. J. Mol. Sci. 2021, 22, 10127. [Google Scholar] [CrossRef]
- Ota, H.; Fujita, Y.; Yamauchi, M.; Muro, S.; Kimura, H.; Takasawa, S. Relationship between intermittent hypoxia and Type 2 diabetes in sleep apnea syndrome. Int. J. Mol. Sci. 2019, 20, 4756. [Google Scholar] [CrossRef] [Green Version]
- Kimura, H.; Ota, H.; Kimura, Y.; Takasawa, S. Effects of intermittent hypoxia on pulmonary vascular and systemic diseases. Int. J. Environ. Res. Public Health 2019, 16, 3101. [Google Scholar] [CrossRef] [Green Version]
- Francia, S.; Michelini, F.; Saxena, A.; Tang, D.; de Hoon, M.; Anelli, V.; Mione, M.; Carninci, P.; d’Adda di Fagagna, F. Site-specific DICER and DROSHA RNA products control the DNA-damage response. Nature 2012, 488, 231–235. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.-H.; Wang, C.-X.; Liu, B.; Fan, X.-S.; Wen, J.-J.; Shi, Q.-L.; Zhou, X.-J. Dysregulation of microRNA biosynthesis enzyme Dicer plays an important role in gastric cancer progression. Int. J. Clin. Exp. Pathol. 2014, 7, 1702–1707. [Google Scholar] [PubMed]
- Baylor, P.; Mouton, A.; Shamoon, H.H.; Goebel, P. Increased norepinephrine variability in patients with sleep apnea syndrome. Am. J. Med. 1995, 99, 611–615. [Google Scholar] [CrossRef]
- Ziegler, M.G.; Nelesen, R.; Mills, P.; Ancoli-Israel, S.; Kennedy, B.; Dimsdale, J.E. Sleep apnea, norepinephrine-release rate, and daytime hypertension. Sleep 1997, 20, 224–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L.; Prabhakar, N.R. Neural regulation of hypoxia-inducible factors and redox state drives the pathogenesis of hypertension in a rodent model of sleep apnea. J. Appl. Physiol. 2015, 119, 1152–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khurana, S.; Peng, S.; McDonald, E.; Yates, W.; Venkataraman, K.; Tai, T.C. Phenylethanolamine N-methyltransferase gene expression in PC12 cells exposed to intermittent hypoxia. Neurosci. Lett. 2018, 666, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Nurse, C.A.; Salman, S.; Scott, A.L. Hypoxia-regulated catecholamine secretion in chromaffin cells. Cell Tissue Res. 2018, 372, 433–441. [Google Scholar] [CrossRef]
- Makarenko, V.V.; Peng, Y.J.; Khan, S.A.; Nanduri, J.; Fox, A.P.; Prabhakar, N.R. Long-term facilitation of catecholamine secretion from adrenal chromaffin cells of neonatal rats by chronic intermittent hypoxia. J. Neurophysiol. 2019, 122, 1874–1883. [Google Scholar] [CrossRef]
- Gridina, A.; Su, X.; Khan, S.A.; Peng, Y.J.; Wang, B.; Nanduri, J.; Fox, A.P.; Prabhakar, N.R. Gaseous transmitter regulation of hypoxia-evoked catecholamine secretion from murine adrenal chromaffin cells. J. Neurophysiol. 2021, 125, 1533–1542. [Google Scholar] [CrossRef]
- Grobecker, H.; Roizen, M.F.; Weise, V.; Saavedra, J.M.; Kopin, I.J. Sympathoadrenal medullary activity in young, spontaneously hypertensive rats. Nature 1975, 258, 267. [Google Scholar] [CrossRef]
- Grobecker, H.; Saavedra, J.M.; Weise, V.K. Biosynthetic enzyme activities and catecholamines in adrenal glands of genetic and experimental hypertensive rats. Circ. Res. 1982, 50, 742–746. [Google Scholar] [CrossRef] [Green Version]
- Moura, E.; Pinho Costa, P.M.; Moura, D.; Guimarães, S.; Vieira-Coelho, M.A. Decreased tyrosine hydroxylase activity in the adrenals of spontaneously hypertensive rats. Life Sci. 2005, 76, 2953–2964. [Google Scholar] [CrossRef]
- Nakamura, K.; Nakamura, K. Enhanced sympathetic activity in young spontaneously hypertensive rats is not the trigger mechanism for genetic hypertension. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1977, 299, 143–148. [Google Scholar] [CrossRef]
- Teitelman, G.; Ross, R.A.; Joh, T.H.; Reis, D.J. Differences in utero in activities of catecholamine biosynthetic enzymes in adrenals of spontaneously hypertensive rats. Clin. Sci. 1981, 61, 227s–230s. [Google Scholar] [CrossRef] [Green Version]
- Kumai, T.; Tanaka, M.; Watanabe, M.; Kobayashi, S. Elevated tyrosine hydroxylase mRNA levels in the adrenal medulla of spontaneously hypertensive rats. Jpn. J. Pharmacol. 1994, 65, 367–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, H.; De Jong, W.; Lovenberg, W. Further studies on catecholamine synthesis in the spontaneously hypertensive rat: Catecholamine synthesis in the central nervous system. Eur. J. Pharmacol. 1973, 22, 91–98. [Google Scholar] [CrossRef]
- Nagatsu, T.; Kato, T.; Numata, Y.; Ikuta, K.; Umezawa, H.; Matsuzaki, M.; Takeuchi, T. Serum dopamine β-hydroxylase activity in developing hypertensive rats. Nature 1974, 251, 630–631. [Google Scholar] [CrossRef] [PubMed]
- Nagatsu, T.; Ikuta, K.; Numata, Y.; Kato, T.; Sano, M. Vascular and brain dopamine β-hydroxylase activity in young spontaneously hypertensive rats. Science 1976, 191, 290–291. [Google Scholar] [CrossRef]
- Ciaranello, R.D.; Hoffman, H.J.; Shire, J.G.; Axelrod, J. Genetic regulation of the catecholamine biosynthetic enzymes. II. Inheritance of tyrosine hydroxylase, Dopamine-β-hydroxylase, and phenylethanolamine N-methyltransferase. J. Biol. Chem. 1974, 249, 4528–4536. [Google Scholar] [CrossRef]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Poy, M.N.; Hausser, J.; Trajkovski, M.; Braun, M.; Collins, S.; Rorsman, P.; Zavolan, M.; Stoffel, M. miR-375 maintains normal pancreatic α- and β-cell mass. Proc. Natl. Acad. Sci. USA 2009, 106, 5813–5818. [Google Scholar] [CrossRef] [Green Version]
- Frank, A.-C.; Ebersberger, S.; Fink, A.F.; Lampe, S.; Weigert, A.; Schmid, T.; Ebersberger, I.; Syed, S.N.; Brüne, B. Apoptotic tumor cell-derived microRNA-375 uses CD36 to alter the tumor-associated macrophage phenotype. Nat. Commun. 2019, 10, 1135. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Gao, J.; Zhang, D.; Liu, H.; Gou, K.; Cui, S. miR-375 acts as a novel factor modulating pituitary prolactin synthesis through Rasd1 and Esr1. J. Endocrinol. 2021, 250, 25–35. [Google Scholar] [CrossRef]
- Yang, X.; Nanayakkara, J.; Claypool, D.; Saghafinia, S.; Wong, J.J.M.; Xu, M.; Wang, X.; Nicol, C.J.B.; Michael, I.P.; Hafner, M.; et al. A miR-375/YAP axis regulates neuroendocrine differentiation and tumorigenesis in lung carcinoid cells. Sci. Rep. 2021, 11, 10455. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-Y.; Song, W.; Lu, F.-F.; Dai, Z.-J.; Cao, L.-H.; Lin, S. miR-375 inhibits the proliferation and invasion of nasopharyngeal carcinoma cells by suppressing PDK1. Biomed. Res. Int. 2020, 2020, 9704245. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhang, Y.; Li, X.; Wang, X.; Yuan, Y. miR-375 inhibits the proliferation, migration and invasion of esophageal squamouse cell carcinoma by targeting XPR1. Curr. Gene Ther. 2021, 21, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-de-la-Torre, M.; Khalyfa, A.; Sánchez-de-la-Torre, A.; Martinez-Alonso, M.; Martinez-García, M.Á.; Barceló, A.; Lloberes, P.; Campos-Rodriguez, F.; Capote, F.; Diaz-de-Atauri, M.J.; et al. Precision medicine in patients with resistant hypertension and obstructive sleep apnea: Blood pressure response to continuous positive airway pressure treatment. J. Am. Coll. Cardiol. 2015, 66, 1023–1032. [Google Scholar] [CrossRef] [Green Version]
- Bouça, B.; Bogalho, P.; Rizzo, M.; Silva-Nunes, J. The role of metabolome and non-coding RNA on pheochromocytomas and paragangliomas: An update. Metabolites 2022, 12, 131. [Google Scholar] [CrossRef]
- Chen, X.; Li, B.; Luo, R.; Cai, S.; Zhang, C.; Cao, X. Analysis of the function of microRNA-375 in humans using bioinformatics. Biomed. Rep. 2017, 6, 561–566. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, J.; Cheng, P.; Lv, F. Correlation between miRNA target site polymorphisms in the 3′ UTR of AVPR1A and the risk of hypertension in the Chinese Han population. Biosci. Rep. 2019, 39, BSR20182232. [Google Scholar] [CrossRef] [Green Version]
- Nakazawa, T.; Takasawa, S.; Noguchi, N.; Nata, K.; Tohgo, A.; Mori, M.; Nakagawara, K.; Akiyama, T.; Ikeda, T.; Yamauchi, A.; et al. Genomic organization, chromosomal localization, and promoter of human gene for FK506-binding protein 12.6. Gene 2005, 360, 55–64. [Google Scholar] [CrossRef]
- Takasawa, S.; Kuroki, M.; Nata, K.; Noguchi, N.; Ikeda, T.; Yamauchi, A.; Ota, H.; Itaya-Hironaka, A.; Sakuramoto-Tsuchida, S.; Takahashi, I.; et al. A novel ryanodine receptor expressed in pancreatic islets by alternative splicing from type 2 ryanodine receptor gene. Biochem. Biophys. Res. Commun. 2010, 397, 140–145. [Google Scholar] [CrossRef]
- Shobatake, R.; Takasawa, K.; Ota, H.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Uchiyama, T.; Makino, M.; Sugie, K.; Takasawa, S.; et al. Up-regulation of POMC and CART mRNAs by intermittent hypoxia via GATA transcription factors in human neuronal cells. Int. J. Biochem. Cell Biol. 2018, 95, 100–107. [Google Scholar] [CrossRef]
- Ota, H.; Tamaki, S.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Morioka, T.; Takasawa, S.; Kimura, H. Attenuation of glucose-induced insulin secretion by intermittent hypoxia via down-regulation of CD38. Life Sci. 2012, 90, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Ota, H.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Miyaoka, T.; Fujimura, T.; Tsujinaka, H.; Yoshimoto, K.; Nakagawara, K.; Tamaki, S.; et al. Pancreatic β cell proliferation by intermittent hypoxia via up-regulation of Reg family genes and HGF gene. Life Sci. 2013, 93, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, T.; Ota, H.; Itaya-Hironaka, A.; Shobatake, R.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Makino, M.; Kimura, H.; Takeda, M.; Ohbayashi, C.; et al. Up-regulation of selenoprotein P and HIP/PAP mRNAs in hepatocytes by intermittent hypoxia via down-regulation of miR-203. Biochem. Biophys. Rep. 2017, 11, 130–137. [Google Scholar] [CrossRef]
- Kyotani, Y.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Makino, M.; Takasawa, S.; Yoshizumi, M. Intermittent hypoxia-induced epiregulin expression by IL-6 production in human coronary artery smooth muscle cells. FEBS Open Bio 2018, 8, 868–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchiyama, T.; Itaya-Hironaka, A.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Shobatake, R.; Ota, H.; Takeda, M.; Ohbayashi, C.; Takasawa, S. Intermittent hypoxia up-regulates CCL2, RETN, and TNFα mRNAs in adipocytes via down-regulation of miR-452. Int. J. Mol. Sci. 2019, 20, 1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shobatake, R.; Itaya-Hironaka, A.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Uchiyama, T.; Ota, H.; Takahashi, N.; Ueno, S.; Sugie, K.; et al. Intermittent hypoxia up-regulates gene expressions of peptide YY (PYY), glucagon-like peptide-1 (GLP-1), and neurotensin (NTS) in enteroendocrine cells. Int. J. Mol. Sci. 2019, 20, 1849. [Google Scholar] [CrossRef] [Green Version]
- Murphy, A.M.; Thomas, A.; Crinion, S.J.; Kent, B.D.; Tambuwala, M.M.; Fabre, A.; Pepin, J.-L.; Roche, H.M.; Arnaud, C.; Ryan, S. Intermittent hypoxia in obstructive sleep apnoea mediates insulin resistance through adipose tissue inflammation. Eur. Respir. J. 2017, 49, 1601731. [Google Scholar] [CrossRef] [Green Version]
- Ota, H.; Takasawa, S.; Yamauchi, M.; Yoshikawa, M.; Tomoda, K.; Kimura, H. Intermittent hypoxia in pancreatic beta cells. Pancreat. Disord. Ther. 2015, 5, S5-004. [Google Scholar] [CrossRef] [Green Version]
- Tohma, Y.; Dohi, Y.; Shobatake, R.; Uchiyama, T.; Takeda, M.; Takasawa, S.; Tanaka, Y.; Ohgushi, H. Reg gene expression in periosteum after fracture and its in vitro induction triggered by IL-6. Int. J. Mol. Sci. 2017, 18, 2257. [Google Scholar] [CrossRef] [Green Version]
- Takasawa, S.; Tsuchida, C.; Sakuramoto-Tsuchida, S.; Takeda, M.; Itaya-Hironaka, A.; Yamauchi, A.; Misu, M.; Shobatake, R.; Uchiyama, T.; Makino, M.; et al. Expression of human REG family genes in inflammatory bowel disease and their molecular mechanism. Immunol. Res. 2018, 66, 800–805. [Google Scholar] [CrossRef]
- Akasaka, J.; Naruse, K.; Sado, T.; Uchiyama, T.; Makino, M.; Yamauchi, A.; Ota, H.; Sakuramoto-Tsuchida, S.; Itaya-Hironaka, A.; Takasawa, S.; et al. Involvement of receptor for advanced glycation endproducts in hypertensive disorders of pregnancy. Int. J. Mol. Sci. 2019, 20, 5462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takasawa, S.; Tohgo, A.; Noguchi, N.; Koguma, T.; Nata, K.; Sugimoto, T.; Yonekura, H.; Okamoto, H. Synthesis and hydrolysis of cyclic ADP-ribose by human leukocyte antigen CD38 and inhibition of the hydrolysis by ATP. J. Biol. Chem. 1993, 268, 26052–26054. [Google Scholar] [CrossRef]
- Yoshimoto, K.; Fujimoto, T.; Itaya-Hironaka, A.; Miyaoka, T.; Sakuramoto-Tsuchida, S.; Yamauchi, A.; Takeda, M.; Kasai, T.; Nakagawara, K.; Nonomura, A.; et al. Involvement of autoimmunity to REG, a regeneration factor, in patients with primary Sjögren’s syndrome. Clin. Exp. Immunol. 2013, 174, 1–9. [Google Scholar] [CrossRef]
- Kato, I.; Yamamoto, Y.; Fujimura, M.; Noguchi, N.; Takasawa, S.; Okamoto, H. CD38 disruption impairs glucose-induced increases in cyclic ADP-ribose, [Ca2+]i, and insulin secretion. J. Biol. Chem. 1999, 274, 1869–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikehata, F.; Satoh, J.; Nata, K.; Tohgo, A.; Nakazawa, T.; Kato, I.; Kobayashi, S.; Akiyama, T.; Takasawa, S.; Toyota, T.; et al. Autoantibodies against CD38 (ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase) that impair glucose-induced insulin secretion in noninsulin-dependent diabetes patients. J. Clin. Investig. 1998, 102, 395–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shervani, N.J.; Takasawa, S.; Uchigata, Y.; Akiyama, T.; Nakagawa, K.; Noguchi, N.; Takada, H.; Takahashi, I.; Yamauchi, A.; Ikeda, T.; et al. Autoantibodies to REG, a beta-cell regeneration factor, in diabetic patients. Eur. J. Clin. Investig. 2004, 34, 752–758. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target mRNA | Primer Sequence |
|---|---|
| Th (NM_009377) | 5′-GCCAAGGACAAGCTCAGGAAC-3′ |
| 5′-ATCAATGGCCAGGGTGTACG-3′ | |
| Ddc (NM_001190448) | 5′-CTGCAGGCTTACATCCGAA-3′ |
| 5′-TTGATCTCTGAAGCAGCT-3′ | |
| Dbh (NM_138942) | 5′-GACTCAACTACTGCCGGCACGT-3′ |
| 5′-CTGGGTGCACTTGTCTGTGCAGT-3′ | |
| Pnmt (NM_008890) | 5′-GTGAAGCGAGTCCTGCCTATC-3′ |
| 5′-AAGATGCCTTTGACATCATCTACC-3′ | |
| Rig/RpS15 (NM_009091) | 5′-ACGGCAAGACCTTCAACCAG-3′ |
| 5′-ATGGAGAACTCGCCCAGGTAG-3′ |
| Target mRNA/MiR | Primer Sequence |
|---|---|
| TH (NM_199292) | 5′-GGAGTTCGGGCTGTGTAAGCA-3′ |
| 5′-GACTGGTACGTCTGGTCTTGGTAGG-3′ | |
| DDC (NM_001082971) | 5′-GAACAGACTTAACGGGAGCCTTT-3′ |
| 5′-AATGCCGGTAGTCAGTGATAAGC-3′ | |
| DBH (NM_000787) | 5′-GTGCTACATTAAGGAGCTTCCAAAG-3’ |
| 5′-GGCCTCATTGCCCTTGGT-3′ | |
| PNMT (NM_002686) | 5′-CTGACTCGGCCCCGGGCCAG-3′ |
| 5′-GGCCTCCCCAGCCAGGTACC-3′ | |
| Dicer (NM_177438) | 5′-GAGCTGTCCTATCAGATCAGGG-3′ |
| 5′-ACTTGTTGAGCAACCTGGTTT-3′ | |
| β-actin (NM_001101) | 5′-GCGAGAAGATGACCCAGA-3′ |
| 5′-CAGAGGCGTACAGGGATA-3′ | |
| miR-375 (NR_029867) | 5′-AGCCGTTTGTTCGTTCGGCT-3′ |
| 5′-GTGCAGGGTCCGAGGT-3′ | |
| U6 (NR_004394) | 5′-CTCGCTTCGGCAGCACA-3′ |
| 5′-AACGCTTCACGAATTTGCGT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takasawa, S.; Shobatake, R.; Takeda, Y.; Uchiyama, T.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Asai, K.; Ota, H.; Itaya-Hironaka, A. Intermittent Hypoxia Increased the Expression of DBH and PNMT in Neuroblastoma Cells via MicroRNA-375-Mediated Mechanism. Int. J. Mol. Sci. 2022, 23, 5868. https://doi.org/10.3390/ijms23115868
Takasawa S, Shobatake R, Takeda Y, Uchiyama T, Yamauchi A, Makino M, Sakuramoto-Tsuchida S, Asai K, Ota H, Itaya-Hironaka A. Intermittent Hypoxia Increased the Expression of DBH and PNMT in Neuroblastoma Cells via MicroRNA-375-Mediated Mechanism. International Journal of Molecular Sciences. 2022; 23(11):5868. https://doi.org/10.3390/ijms23115868
Chicago/Turabian StyleTakasawa, Shin, Ryogo Shobatake, Yoshinori Takeda, Tomoko Uchiyama, Akiyo Yamauchi, Mai Makino, Sumiyo Sakuramoto-Tsuchida, Keito Asai, Hiroyo Ota, and Asako Itaya-Hironaka. 2022. "Intermittent Hypoxia Increased the Expression of DBH and PNMT in Neuroblastoma Cells via MicroRNA-375-Mediated Mechanism" International Journal of Molecular Sciences 23, no. 11: 5868. https://doi.org/10.3390/ijms23115868
APA StyleTakasawa, S., Shobatake, R., Takeda, Y., Uchiyama, T., Yamauchi, A., Makino, M., Sakuramoto-Tsuchida, S., Asai, K., Ota, H., & Itaya-Hironaka, A. (2022). Intermittent Hypoxia Increased the Expression of DBH and PNMT in Neuroblastoma Cells via MicroRNA-375-Mediated Mechanism. International Journal of Molecular Sciences, 23(11), 5868. https://doi.org/10.3390/ijms23115868