
The microRNA-202 as a Diagnostic Biomarker and a Potential Tumor Suppressor



Abstract
:1. Introduction
2. Circulating miR-202 as a Diagnostic Biomarker in Cancer Patients
3. Upregulation of miR-202 in Breast Cancer Is Correlated with Drug Resistance
4. miR-202 as a Novel Gastrointestinal Tract Tumor Suppressor
4.1. Downregulation of miR-202 in Gastric Cancer Is a Potential Biomarker for Tumor Progression
4.2. The Role of miR-202 in Pancreatic Cancer and Hepatocellular Carcinoma
4.3. miR-202 in Colorectal Tumors
5. Tumor Suppression Function of miR-202 in Non-Small Cell Lung Cancer
6. Tumor Suppressor Function of miR-202 in Different Reproductive System and Gynecological Cancers
7. miR-202’s Role in Osteosarcoma
8. Tumor Suppression Role of miR-202 in Further Types of Cancer
9. Mechanistic Pathways Involving miR-202 as a Tumor Suppressor
9.1. The lncRNA Metastasis-Associated Lung Adenocarcinoma Transcript 1 (MALAT1) Promotes Cancer Progression via Down-Regulating miR-202 Expression
9.2. The lncRNA NORAD Inhibits miR-202 to Promote Cancer Progression
9.3. miR-202 Suppresses Cancer via Targeting the Oncogene Metadherin and the Ras GTPase
9.4. Cyclin D1 Is a Downstream Target of miR-202
9.5. miR-202 Binds to B Cell-Activating Factor (BAFF) and Enhances Tumor Suppression
10. The miR-202/PTEN/AKT Axis in Tumor Progression
11. Micro-RNA-Based Therapeutic Approaches
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Z.; Liu, Z. The Emerging Role of MicroRNAs in Breast Cancer. J. Oncol. 2020, 2020, 9160905. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, E.A.; Ibrahim, H.M.; Khalil, H.E. Pinocembrin Reduces Arthritic Symptoms in Mouse Model via Targeting Sox4 Signalling Molecules. J. Med. Food 2021, 24, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Hanieh, H.; Ahmed, E.A.; Vishnubalaji, R.; Alajez, N.M. SOX4: Epigenetic regulation and role in tumorigenesis. Semin. Cancer Biol. 2020, 67, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Dou, D.; Shi, Y.F.; Liu, Q.; Luo, J.; Liu, J.X.; Liu, M.; Liu, Y.Y.; Li, Y.L.; Qiu, X.D.; Tan, H.Y. Hsa-miR-202-3p, up-regulated in type 1 gastric neuroendocrine neoplasms, may target DUSP1. World J. Gastroenterol. 2018, 24, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Qiu, X.; Shen, X.; Shi, W.; Wu, X.; Gu, G.; Zhu, B.; Ju, S. miR-202 expression concentration and its clinical significance in the serum of multiple myeloma patients. Ann. Clin. Biochem. 2014, 51, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Lai, C.H.; Chen, H.C.; Lin, C.Y.; Tsai, C.L.; Tang, Y.H.; Huang, H.J.; Lin, C.T.; Chen, M.Y.; Huang, K.G.; et al. Serum microRNAs in clear cell carcinoma of the ovary. Taiwan. J. Obstet. Gynecol. 2014, 53, 536–541. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Pu, Y. MicroRNA signatures in total peripheral blood of gallbladder cancer patients. Tumour Biol. 2015, 36, 6985–6990. [Google Scholar] [CrossRef]
- Monastirioti, A.; Papadaki, C.; Rounis, K.; Kalapanida, D.; Mavroudis, D.; Agelaki, S. A Prognostic Role for Circulating microRNAs Involved in Macrophage Polarization in Advanced Non-Small Cell Lung Cancer. Cells 2021, 10, 1988. [Google Scholar] [CrossRef]
- Schrauder, M.G.; Strick, R.; Schulz-Wendtland, R.; Strissel, P.L.; Kahmann, L.; Loehberg, C.R.; Lux, M.P.; Jud, S.M.; Hartmann, A.; Hein, A.; et al. Circulating micro-RNAs as potential blood-based markers for early stage breast cancer detection. PLoS ONE 2012, 7, e29770. [Google Scholar] [CrossRef]
- Joosse, S.A.; Muller, V.; Steinbach, B.; Pantel, K.; Schwarzenbach, H. Circulating cell-free cancer-testis MAGE-A RNA, BORIS RNA, let-7b and miR-202 in the blood of patients with breast cancer and benign breast diseases. Br. J. Cancer 2014, 111, 909–917. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Park, S.; Hwang, D.; Kim, S.I.; Lee, H. Diagnostic Value of Circulating miR-202 in Early-Stage Breast Cancer in South Korea. Medicina 2020, 56, 340. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ding, D.; Zhao, G. Reduced miR-202 levels enhanced oral cancer development via targeting Sp1. Exp. Ther. Med. 2019, 18, 489–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Guo, J.; Zhang, X. MiR-202-5p/PTEN mediates doxorubicin-resistance of breast cancer cells via PI3K/Akt signalling pathway. Cancer Biol. Ther. 2019, 20, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Du, C.; Zhang, L.; Wang, Y.; Zhang, Y.; Li, J. lncRNA GSEC Promotes the Progression of Triple Negative Breast Cancer (TNBC) by Targeting the miR-202-5p/AXL Axis. Onco Targets Ther. 2021, 14, 2747–2759. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Cao, C.; Dai, Q.; Chen, J.; Tu, J. miR-202 acts as a potential tumor suppressor in breast cancer. Oncol. Lett. 2018, 16, 1155–1162. [Google Scholar] [CrossRef]
- Harati, R.; Hafezi, S.; Mabondzo, A.; Tlili, A. Silencing miR-202-3p increases MMP-1 and promotes a brain invasive phenotype in metastatic breast cancer cells. PLoS ONE 2020, 15, e0239292. [Google Scholar] [CrossRef]
- Ibrahim, H.M.; Ismail, M.B.; Ammar, R.B.; Ahmed, E.A. Thidiazuron suppresses breast cancer via targeting miR-132 and dysregulation of the PI3K-Akt signalling pathway mediated by the miR-202-5p-PTEN axis. Biochem. Cell Biol. 2021, 99, 374–384. [Google Scholar] [CrossRef]
- Xu, F.; Li, H.; Hu, C. MiR-202 inhibits cell proliferation, invasion, and migration in breast cancer by targeting ROCK1 gene. J. Cell. Biochem. 2019, 120, 16008–16018. [Google Scholar] [CrossRef]
- Meng, X.; Chen, X.; Lu, P.; Ma, W.; Yue, D.; Song, L.; Fan, Q. MicroRNA-202 inhibits tumor progression by targeting LAMA1 in esophageal squamous cell carcinoma. Biochem. Biophys. Res. Commun. 2016, 473, 821–827. [Google Scholar] [CrossRef]
- Meng, X.; Chen, X.; Lu, P.; Ma, W.; Yue, D.; Song, L.; Fan, Q. miR-202 Promotes Cell Apoptosis in Esophageal Squamous Cell Carcinoma by Targeting HSF2. Oncol. Res. 2017, 25, 215–223. [Google Scholar] [CrossRef]
- Meng, X.R.; Lu, P.; Mei, J.Z.; Liu, G.J.; Fan, Q.X. Expression analysis of miRNA and target mRNAs in esophageal cancer. Braz. J. Med. Biol. Res. 2014, 47, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Li, C.; Wang, M.; Su, L.; Qu, Y.; Li, J.; Yu, B.; Yan, M.; Yu, Y.; Liu, B.; et al. Decrease of miR-202-3p expression, a novel tumor suppressor, in gastric cancer. PLoS ONE 2013, 8, e69756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, Z.; Li, M.J.; Guo, H.Y.; Jing, N.C. Long non-coding RNA metastasis-associated lung adenocarcinoma transcript 1 regulates the expression of Gli2 by miR-202 to strengthen gastric cancer progression. Biomed. Pharmacother. 2017, 85, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Tan, X.; Tang, H.; Liu, J. CCL21 activation of the MALAT1/SRSF1/mTOR axis underpins the development of gastric carcinoma. J. Transl. Med. 2021, 19, 210. [Google Scholar] [CrossRef] [PubMed]
- Farhana, L.; Dawson, M.I.; Fontana, J.A. Down regulation of miR-202 modulates Mxd1 and Sin3A repressor complexes to induce apoptosis of pancreatic cancer cells. Cancer Biol. Ther. 2015, 16, 115–124. [Google Scholar] [CrossRef]
- Mody, H.R.; Hung, S.W.; Pathak, R.K.; Griffin, J.; Cruz-Monserrate, Z.; Govindarajan, R. miR-202 Diminishes TGFbeta Receptors and Attenuates TGFbeta1-Induced EMT in Pancreatic Cancer. Mol. Cancer Res. 2017, 15, 1029–1039. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.J.; Li, X.D.; Wu, L.P.; Guo, P.; Feng, L.X.; Li, B. MicroRNA-202 suppresses glycolysis of pancreatic cancer by targeting hexokinase 2. J. Cancer 2021, 12, 1144–1153. [Google Scholar] [CrossRef]
- Ma, Y.S.; Yang, X.L.; Liu, Y.S.; Ding, H.; Wu, J.J.; Shi, Y.; Jia, C.Y.; Lu, G.X.; Zhang, D.D.; Wang, H.M.; et al. Long non-coding RNA NORAD promotes pancreatic cancer stem cell proliferation and self-renewal by blocking microRNA-202-5p-mediated ANP32E inhibition. J. Transl. Med. 2021, 19, 400. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, D.; Xiong, Y.; Xue, C.; Chen, G.; Yan, B.; Ye, Q. miR-202 suppresses cell proliferation in human hepatocellular carcinoma by downregulating LRP6 post-transcriptionally. FEBS Lett. 2014, 588, 1913–1920. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Cai, J.B.; Peng, R.; Wei, C.Y.; Lu, J.C.; Gao, C.; Shen, Z.Z.; Zhang, P.F.; Huang, X.Y.; Ke, A.W.; et al. The long noncoding RNA NORAD enhances the TGF-beta pathway to promote hepatocellular carcinoma progression by targeting miR-202-5p. J. Cell Physiol. 2019, 234, 12051–12060. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, J.; Sun, F.; Wang, Z.; Xu, W.; Yu, Y.; Ding, F.; Shen, H. miR-202 functions as a tumor suppressor in hepatocellular carcinoma by targeting HK2. Oncol. Lett. 2020, 19, 2265–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, S.B.; Qiu, H.; Chen, J.M.; Shi, W.; Chen, Y.S. MicroRNA-202-5p functions as a tumor suppressor in colorectal carcinoma by directly targeting SMARCC1. Gene 2018, 676, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Chen, Z.; Lin, S.; Zheng, Y.; Liu, Y.; Gao, J.; Chen, S. MiR-202 inhibits the proliferation and invasion of colorectal cancer by targeting UHRF1. Acta Biochim. Biophys. Sin. 2019, 51, 1305–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Huang, Z.; Guo, W.; Ni, S.; Xiao, X.; Wang, L.; Huang, D.; Tan, C.; Xu, Q.; Zha, R.; et al. microRNA-202-3p inhibits cell proliferation by targeting ADP-ribosylation factor-like 5A in human colorectal carcinoma. Clin. Cancer Res. 2014, 20, 1146–1157. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, X.Y.; Hu, P.; Ding, Y.S. lncRNA NORAD Contributes to Colorectal Cancer Progression by Inhibition of miR-202-5p. Oncol. Res. 2018, 26, 1411–1418. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, Y.; Xu, Y.; Xie, Q.; Wu, S.; Dong, Y.; Chen, B.; Xia, Y.; Guo, L.; Li, Q.; et al. MiRNA-202-5p promotes Colorectal Carcinogenesis through suppression of PTEN. J. Cancer 2021, 12, 3154–3163. [Google Scholar] [CrossRef]
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Global Cancer Observatory: Cancer Today; International Agency for Research on Cancer: Lyon, France, 2020. [Google Scholar]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Zhuang, D.; Liang, L.; Zhang, H.; Feng, X. miR-202 Suppresses Hepatocellular Carcinoma Progression via Downregulating BCL2 Expression. Oncol. Res. 2020, 28, 399–408. [Google Scholar] [CrossRef]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Herbst, R.S.; Boshoff, C. Toward personalized treatment approaches for non-small-cell lung cancer. Nat. Med. 2021, 27, 1345–1356. [Google Scholar] [CrossRef] [PubMed]
- Nymark, P.; Guled, M.; Borze, I.; Faisal, A.; Lahti, L.; Salmenkivi, K.; Kettunen, E.; Anttila, S.; Knuutila, S. Integrative analysis of microRNA, mRNA and aCGH data reveals asbestos- and histology-related changes in lung cancer. Genes Chromosom. Cancer 2011, 50, 585–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Huang, J.; Wang, X.R.; Quan, Y.H. MicroRNA-202 induces cell cycle arrest and apoptosis in lung cancer cells through targeting cyclin D1. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2278–2284. [Google Scholar]
- Zhao, Z.; Lv, B.; Zhang, L.; Zhao, N.; Lv, Y. miR-202 functions as a tumor suppressor in non-small cell lung cancer by targeting STAT3. Mol. Med. Rep. 2017, 16, 2281–2289. [Google Scholar] [CrossRef]
- Sun, W.; Ping, W.; Tian, Y.; Zou, W.; Liu, J.; Zu, Y. miR-202 Enhances the Anti-Tumor Effect of Cisplatin on Non-Small Cell Lung Cancer by Targeting the Ras/MAPK Pathway. Cell. Physiol. Biochem. 2018, 51, 2160–2171. [Google Scholar] [CrossRef]
- Peng, J.; Chen, X.L.; Cheng, H.Z.; Xu, Z.Y.; Wang, H.; Shi, Z.Z.; Liu, J.; Ning, X.G.; Peng, H. Silencing of KCNK15-AS1 inhibits lung cancer cell proliferation via upregulation of miR-202 and miR-370. Oncol. Lett. 2019, 18, 5968–5976. [Google Scholar] [CrossRef] [Green Version]
- Tiansheng, G.; Junming, H.; Xiaoyun, W.; Peixi, C.; Shaoshan, D.; Qianping, C. lncRNA Metastasis-Associated Lung Adenocarcinoma Transcript 1 Promotes Proliferation and Invasion of Non-Small Cell Lung Cancer Cells via Down-Regulating miR-202 Expression. Cell J. 2020, 22, 375–385. [Google Scholar] [CrossRef]
- Shen, J.G.; Xu, S.N.; Yin, L.G. LncRNA NORAD/miR-202-5p regulates the drug resistance of A549/DDP to cisplatin by targeting P-gp. Gen. Physiol. Biophys. 2020, 39, 481–489. [Google Scholar] [CrossRef]
- Wang, R.; Chen, X.F.; Shu, Y.Q. Prediction of non-small cell lung cancer metastasis-associated microRNAs using bioinformatics. Am. J. Cancer Res. 2015, 5, 32–51. [Google Scholar]
- Li, Y.; Huang, H.; Ye, X.; Huang, Z.; Chen, X.; Wu, F.; Lin, T. miR-202-3p negatively regulates MMP-1 to inhibit the proliferation, migration and invasion of lung adenocarcinoma cells. Cell Cycle 2021, 20, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Wang, Q.; Wang, Y.; Hu, B.; Dong, X.; Zhang, H.; Wang, W. Long non-coding RNA metastasis-associated lung adenocarcinoma transcript 1/microRNA-202-3p/periostin axis modulates invasion and epithelial-mesenchymal transition in human cervical cancer. J. Cell. Physiol. 2019, 234, 14170–14180. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Li, H.; Lv, Q.; Wu, K.; Zhang, W.; Zhang, J.; Zhu, D.; Liu, Q.; Zhang, W. miR-202 inhibits the progression of human cervical cancer through inhibition of cyclin D1. Oncotarget 2016, 7, 72067–72075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Xie, L.; Xu, S.; Yan, W.; Zhang, H.; Meng, Y.; Liu, J.; Wei, X. Effects of miR-202-5p silencing PIK3CA gene expression on proliferation, invasion, and epithelial-mesenchymal transition of cervical cancer SiHa cells through inhibiting PI3K/Akt/mTOR signalling pathway activation. Mol. Cell. Biochem. 2021, 476, 4031–4044. [Google Scholar] [CrossRef]
- Causin, R.L.; da Silva, L.S.; Evangelista, A.F.; Leal, L.F.; Souza, K.C.B.; Pessoa-Pereira, D.; Matsushita, G.M.; Reis, R.M.; Fregnani, J.; Marques, M.M.C. MicroRNA Biomarkers of High-Grade Cervical Intraepithelial Neoplasia in Liquid Biopsy. Biomed. Res. Int. 2021, 2021, 6650966. [Google Scholar] [CrossRef]
- Yu, H.Y.; Pan, S.S. MiR-202-5p suppressed cell proliferation, migration and invasion in ovarian cancer via regulating HOXB2. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2256–2263. [Google Scholar] [CrossRef]
- Chang, R.K.; Li, X.; Mu, N.; Hrydziuszko, O.; Garcia-Majano, B.; Larsson, C.; Lui, W.O. MicroRNA expression profiles in non-epithelial ovarian tumors. Int. J. Oncol. 2018, 52, 55–66. [Google Scholar] [CrossRef]
- Deng, X.; Hou, C.; Liang, Z.; Wang, H.; Zhu, L.; Xu, H. miR-202 Suppresses Cell Proliferation by Targeting FOXR2 in Endometrial Adenocarcinoma. Dis. Markers 2017, 2017, 2827435. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Xing, T.; Wang, Q.; Liu, A.; Liu, H.; Hu, Y.; Ji, Y.; Song, Y.; Wang, D. MicroRNA-202 inhibits cell migration and invasion through targeting FGF2 and inactivating Wnt/beta-catenin signalling in endometrial carcinoma. Biosci. Rep. 2019, 39, BSR20190680. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Zhai, J.; Fu, Y. Overexpression of Nuclear Enriched Autosomal Transcript 1 Facilitates Cell Proliferation, Migration Invasion, and Suppresses Apoptosis in Endometrial Cancer by Targeting MicroRNA-202-3p/T Cell Immunoglobulin and Mucin Domain 4 Axis. Cancer Biother. Radiopharm. 2020. [Google Scholar] [CrossRef]
- De Almeida, B.C.; Garcia, N.; Maffazioli, G.; dos Anjos, L.G.; Baracat, E.C.; Carvalho, K.C. Oncomirs Expression Profiling in Uterine Leiomyosarcoma Cells. Int. J. Mol. Sci. 2017, 19, 52. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Cai, J.; Xie, W.; Luo, H.; Yang, F. miR-202 suppresses prostate cancer growth and metastasis by targeting PIK3CA. Exp. Ther. Med. 2018, 16, 1499–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, A.C.; Vira, M.; Walter, V.; Shen, J.; Raman, J.D.; Sanda, M.G.; Patil, D.; Taioli, E. Circulating microRNAs in plasma among men with low-grade and high-grade prostate cancer at prostate biopsy. Prostate 2019, 79, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Brooks, N.; Pouniotis, D.S. Immunomodulation in endometrial cancer. Int. J. Gynecol. Cancer 2009, 19, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yin, J.; Liu, J.; Zhu, R.X.; Zheng, Y.; Wang, X.L. MiR-202-3p functions as a tumor suppressor and reduces cell migration and invasion in papillary thyroid carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1145–1150. [Google Scholar] [CrossRef] [PubMed]
- Sadykova, L.R.; Ntekim, A.I.; Muyangwa-Semenova, M.; Rutland, C.S.; Jeyapalan, J.N.; Blatt, N.; Rizvanov, A.A. Epidemiology and Risk Factors of Osteosarcoma. Cancer Investig. 2020, 38, 259–269. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, T.; Hong, H.; Liu, Q.; Zhang, H. miR-202 suppresses proliferation and induces apoptosis of osteosarcoma cells by downregulating Gli2. Mol. Cell. Biochem. 2014, 397, 277–283. [Google Scholar] [CrossRef]
- Li, C.; Ma, D.; Yang, J.; Lin, X.; Chen, B. miR-202-5p inhibits the migration and invasion of osteosarcoma cells by targeting ROCK1. Oncol. Lett. 2018, 16, 829–834. [Google Scholar] [CrossRef] [Green Version]
- Diao, C.Y.; Guo, H.B.; Ouyang, Y.R.; Zhang, H.C.; Liu, L.H.; Bu, J.; Wang, Z.H.; Xiao, T. Screening for metastatic osteosarcoma biomarkers with a DNA microarray. Asian Pac. J. Cancer Prev. 2014, 15, 1817–1822. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Piao, C.D.; Ding, J.; Li, Z.W. LncRNA MALAT1 facilitates lung metastasis of osteosarcomas through miR-202 sponging. Sci. Rep. 2020, 10, 12757. [Google Scholar] [CrossRef]
- Lin, Z.; Song, D.; Wei, H.; Yang, X.; Liu, T.; Yan, W.; Xiao, J. TGF-beta1-induced miR-202 mediates drug resistance by inhibiting apoptosis in human osteosarcoma. J. Cancer Res. Clin. Oncol. 2016, 142, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Tang, J.; Hu, F.; Liao, Y.; Li, R.; Zhou, Y.; Yao, Z.; Geng, Z.; Yang, Z.; Zhang, X.; et al. Effects of different levels of TGF-β expression and tumor cell necrosis rates in osteosarcoma on the chemotherapy resistance of osteosarcoma. J. Bone Oncol. 2020, 23, 100299. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Yang, H.; Liu, D.; Pei, R. LncRNA NORAD promotes thyroid carcinoma progression through targeting miR-202-5p. Am. J. Transl. Res. 2019, 11, 290–299. [Google Scholar] [PubMed]
- Zhang, Y.; Shen, X.J.; Wu, X.H.; Cong, H.; Ni, H.B.; Ju, S.Q.; Su, J.Y. miR-202 contributes to sensitizing MM cells to drug significantly via activing JNK/SAPK signalling pathway. Zhonghua Xue Ye Xue Za Zhi 2016, 37, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.J.; Shen, X.J.; Wang, X.D.; Ju, S.Q. Effect of miR-202 on the growth of multiple myeloma cells via regulating B cell-activating factor and the underlying mechanism. Zhonghua Zhong Liu Za Zhi 2013, 35, 886–891. [Google Scholar]
- Musolino, C.; Oteri, G.; Allegra, A.; Mania, M.; D’Ascola, A.; Avenoso, A.; Innao, V.; Allegra, A.G.; Campo, S. Altered microRNA expression profile in the peripheral lymphoid compartment of multiple myeloma patients with bisphosphonate-induced osteonecrosis of the jaw. Ann. Hematol. 2018, 97, 1259–1269. [Google Scholar] [CrossRef]
- Shen, X.; Guo, Y.; Qi, J.; Shi, W.; Wu, X.; Ni, H.; Ju, S. Study on the Association Between miRNA-202 Expression and Drug Sensitivity in Multiple Myeloma Cells. Pathol. Oncol. Res. 2016, 22, 531–539. [Google Scholar] [CrossRef]
- Yang, J.; Fan, B.; Zhao, Y.; Fang, J. MicroRNA-202 inhibits cell proliferation, migration and invasion of glioma by directly targeting metadherin. Oncol. Rep. 2017, 38, 1670–1678. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, A.E.; Liu, R.; Fu, A.; Zheng, T.; Slack, F.; Zhu, Y. Targetome profiling, pathway analysis and genetic association study implicate miR-202 in lymphomagenesis. Cancer Epidemiol. Biomark. Prev. 2013, 22, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Nie, Z.Y.; Yao, M.; Yang, Z.; Yang, L.; Liu, X.J.; Yu, J.; Ma, Y.; Zhang, N.; Zhang, X.Y.; Liu, M.H.; et al. De-regulated STAT5A/miR-202-5p/USP15/Caspase-6 regulatory axis suppresses CML cell apoptosis and contributes to Imatinib resistance. J. Exp. Clin. Cancer Res. 2020, 39, 17. [Google Scholar] [CrossRef]
- Deng, Y.; Li, X.; Feng, J.; Zhang, X. Overexpression of miR-202 resensitizes imatinib resistant chronic myeloid leukemia cells through targetting Hexokinase 2. Biosci. Rep. 2018, 38, BSR20171383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruzgielewicz, A.; Osuch-Wojcikiewicz, E.; Niemczyk, K.; Sieniawska-Buccella, O.; Siwak, M.; Walczak, A.; Nowak, A.; Majsterek, I. Altered Expression of miRNAs Is Related to Larynx Cancer TNM Stage and Patients’ Smoking Status. DNA Cell Biol. 2017, 36, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Guo, Y.; Yu, J.; Qi, J.; Shi, W.; Wu, X.; Ni, H.; Ju, S. miRNA-202 in bone marrow stromal cells affects the growth and adhesion of multiple myeloma cells by regulating B cell-activating factor. Clin. Exp. Med. 2016, 16, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Kang, J.H.; Yoo, K.C.; Kang, S.G.; Lee, H.J.; Lee, S.J. K-RAS Acts as a Critical Regulator of CD44 to Promote the Invasiveness and Stemness of GBM in Response to Ionizing Radiation. Int. J. Mol. Sci. 2021, 22, 10923. [Google Scholar] [CrossRef]
- Buechner, J.; Tomte, E.; Haug, B.H.; Henriksen, J.R.; Lokke, C.; Flaegstad, T.; Einvik, C. Tumour-suppressor microRNAs let-7 and mir-101 target the proto-oncogene MYCN and inhibit cell proliferation in MYCN-amplified neuroblastoma. Br. J. Cancer 2011, 105, 296–303. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Kong, L.B.; Li, J.T.; Guo, Z.Y.; Xue, Q.; Yang, T.; Meng, Y.L.; Jin, B.Q.; Wen, W.H.; Yang, A.G. MiR-568 inhibits the activation and function of CD4(+) T cells and Treg cells by targeting NFAT5. Int. Immunol. 2014, 26, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xu, J.; Yang, G.; Li, H.; Guo, X. miR-202 Inhibits Cell Proliferation, Migration, and Invasion by Targeting Epidermal Growth Factor Receptor in Human Bladder Cancer. Oncol. Res. 2018, 26, 949–957. [Google Scholar] [CrossRef]
- Su, Y.; Lu, S.; Li, J.; Deng, L. Shikonin-mediated up-regulation of miR-34a and miR-202 inhibits retinoblastoma proliferation. Toxicol. Res. (Camb) 2018, 7, 907–912. [Google Scholar] [CrossRef] [Green Version]
- Ji, P.; Diederichs, S.; Wang, W.; Boing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Ding, M.; Wang, B.; Cai, Y.; Guo, C.; Yuan, C. Molecular Mechanism of the Canonical Oncogenic lncRNA MALAT1 in Gastric Cancer. Curr. Med. Chem. 2021, 28, 8800–8809. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, L.; Guo, H.L.; Zhang, Z.W.; Wang, C.; Chian, R.C.; Zhang, Z.F. MicroRNA202 inhibits endometrial stromal cell migration and invasion by suppressing the KRas/Raf1/MEK/ERK signalling pathway. Int. J. Mol. Med. 2020, 46, 2078–2088. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.S.; Whitty, E.; Yoo, B.; Moore, A.; Sempere, L.F.; Medarova, Z. Clinical Applications of Short Non-Coding RNA-Based Therapies in the Era of Precision Medicine. Cancers 2022, 14, 1588. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.S.; Kang, Y.K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.L.; Kim, T.Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]


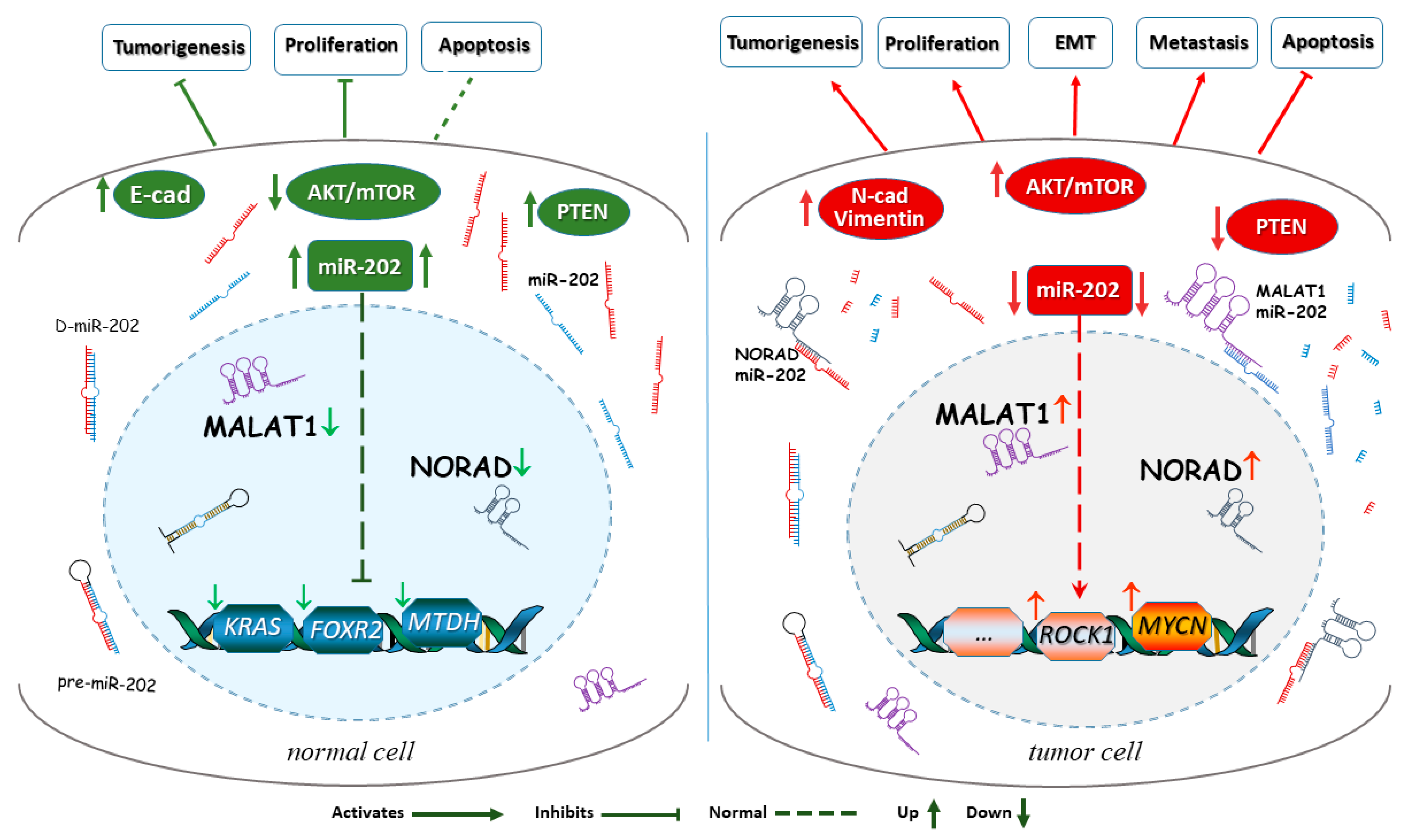{kind=link}
| mir-202 Function | Regulation (Up/Down) | Patients’ Samples, Cell Lines, Animal Model | Involved Downstream/Upstream Molecules | Ref. |
|---|---|---|---|---|
| pro-tumorigenic | up | 23 DOX-resistant, 39 DOX-sensitive BC cancer samples, cells lines, xenografted nude mice | PTEN/PI3k/Akt | [13] |
| pro-tumorigenic | up | BC cell lines | PTEN/PI3k/Akt | [17] |
| pro-tumorigenic | up | 45 pairs of BC cancer and adjacent normal tissue, cell lines | lncRNA GSEC, AXL | [14] |
| pro-tumorigenic | up (early stage) | Plasma samples of 30 BC patients (stages I–III), 30 control samples | n.d. | [11] |
| pro-tumorigenic | up | Sera of 102 BC patients and 26 with benign breast diseases, blood samples of 37 healthy controls | n.d. | [10] |
| pro-tumorigenic | up (early stages) | Blood samples of 48 early-stage BC and 57 controls | n.d. | [9] |
| suppressor | down | 27 BC tissue samples, cell lines | ROCK1, E-cadherin, Twist, N-cadherin, and MMP2 | [18] |
| suppressor | down | BC cell lines | MMP-1, claudin-5, ZO-1 and ß-catenin | [16] |
| suppressor | down | 30 BC tissues | KRAS | [15] |
| mir-202 Function | Regulation | Samples, Cell Lines, Patient Material, Animal Model | Involved Downstream/Upstream Targets | Ref. |
|---|---|---|---|---|
| (Up/Down) | ||||
| Oral Cancer | ||||
| suppressor | down | 73 oral cancer tissue, 48 normal tissues, blood samples, cell lines | Sp1, protein kinase B | [12] |
| Esophageal squamous cell carcinoma (ESCC) | ||||
| suppressor | down | cell lines | HSF2/Hsp70 | [20] |
| down | 76 esophageal cancers (44 ESCC, 32 EAC) with adjacent normal tissues | n.d. | [21] | |
| suppressor | down | 30 primary ESCC tissues and adjacent noncancerous tissues, cell lines | p-FAK, p-Akt, LAMA1 | [19] |
| Gastric Cancer | ||||
| suppressor | down | 60 GC tissues and adjacent normal tissues | MALAT1 | [23] |
| suppressor | down | 150 GC tissues and adjacent normal tissues, mouse xenografts | Gli1, γ-catenin, BCL-2 | [22] |
| suppressor | down | 115 GC tissues with normal tissue samples, cell lines, mouse xenografts | MALAT1, SRSF1, CCL21, mTOR | [24] |
| Hepatocellular carcinoma | ||||
| suppressor | down | 56 HCC samples, cell lines | hexokinase 2, glycolysis | [31] |
| suppressor | down | Eight pairs of snap-frozen HCC tumor, cell lines | LRP6 | [29] |
| suppressor | down | Tissues from 95 patients with HCC who underwent tumor, cell lines, Tumor xenograft | NORAD, NORAD/miR-202-5p/TGF-β axis | [30] |
| Gall bladder cancer | ||||
| pro-tumorigenic | up | Blood samples of 40 GBC patients | n.d. | [7] |
| Pancreatic cancer | ||||
| suppressor | down | PC cell lines & orthotopic mouse xenografts | TGFβ1, EMT | [26] |
| suppressor | down | cell lines | hexokinase 2, inhibited glycolysis | [27] |
| suppressor | down | cell lines | Mxd1, SAP18 | [25] |
| suppressor | down | 28 PC cancer and adjacent normal tissue samples, cell lines, mouse xenografts | lncRNA NORAD and ANP32E | [28] |
| Colorectal cancer | ||||
| suppressor | down | 68 cases of CRC and 66 adjacent normal tissues, cell lines and xenografted nude mice | UHRF1 | [33] |
| suppressor | down | 68 CRC tissues, cell lines | 1SMARCC1 | [32] |
| suppressor | down | 47 pairs of CRC tissues and adjacent normal tissues, cell lines and xenografted nude mice | NORAD | [35] |
| suppressor | down | 98 primary CRC cancer tissues and adjacent normal tissues, cell lines, mouse xenografts | ADP-ribosylation factor-like 5A | [34] |
| pro-tumorigenic | up | 25 CRC tumors and adjacent normal tissues, cell lines | PTEN, c-Myc AKT | [36] |
| mir-202 Function | Regulation | Samples, Cell Lines, Patient Material, Animal Model | Involved Downstream/Upstream Molecules | References |
|---|---|---|---|---|
| suppressor | down | 56 patient tissues and corresponding normal tissues, cell lines | STAT3 | [46] |
| suppressor | down | 40 lung adenocarcinoma and adjacent normal tissue samples | KCNK15-AS1, EGFR, miR-202/miR-370/EGFR axis | [48] |
| suppressor | down | 40 NSCLC tissues, cell lines | MALAT1 | [49] |
| suppressor | down | 25 NSCLC and adjacent paratumor tissues, cell lines, mouse xenografts | KRAS | [47] |
| suppressor | down | cell lines | NORAD | [50] |
| suppressor | down | 20 lung cancer and adjacent normal tissues | cyclin D1 | [45] |
| suppressor | down | 26 tumor and corresponding normal lung tissue samples | n.d. | [44] |
| suppressor | down | Nude mice, cell lines | [34] | |
| pro-tumorigenic | up | 125 plasma samples first-line chemotherapy | Macrophage Polarization | [8] |
| miR-202 Function | Regulation | Samples, Cell Lines, Patient Material, Animal Model | Involved Downstream/Upstream Molecules | Ref. |
|---|---|---|---|---|
| Cervical cancer | ||||
| suppressor | down | 105 patient samples, cell lines | PIK3CA/PI3K/Akt/mTOR pathway | [55] |
| suppressor | down | 100 CC and control tissues, cell lines | cyclin D1 | [54] |
| suppressor | down | 23 CC tissues, cell lines | MALAT1/miR-202-3p/periostin, EMT, Akt/mTOR signaling | [53] |
| Endometrial cancer | ||||
| suppressor | down | 90 tumor & 40 corresponding. normal tissues, cell lines, xenografted nude mice | FOXR2 oncogene | [59] |
| suppressor | down | 76 EC tissue samples | FGF2 and Wnt/β-catenin | [60] |
| suppressor | down | 20 EC and adjacent normal tissue samples, cell lines | NEAT1, TIMD4 | [61] |
| Uterine Leiomyosarcoma | ||||
| pro-tumorigenic | up | 39 paraffin UL tumor samples, cell lines | MYCN | [62] |
| Ovarian cancer | ||||
| pro-tumorigenic | up | 21 patient serum samples | n.d. | [6] |
| suppressor | down | 55 paired OC and para-tumor tissues, cell lines | HOXB2 | [57] |
| pro-tumorigenic | up | 23 non-epithelial ovarian germ cell tumors, 16 ovarian, 7 sex cord stromal tumors | n.d. | [58] |
| Prostate cancer | ||||
| suppressor | down | 54 PC tissues and 11 adjacent normal prostate tissues, cell lines and mouse xenografts | PIK3CA | [63] |
| inconclusive | up in high grade PC | 66 sera of patients with low- and high-grade PC | n.d. | [64] |
| mir-202 Function | Regulation | Samples, Cell Lines, Patient Material, Animal Model | Involved Downstream/Upstream Molecules | Ref. |
|---|---|---|---|---|
| suppressor | down | 36 tumor specimens and normal adjacent tissues, cell lines | ROCK1 | [70] |
| pro-tumorigenic | up | 8 paired tumor and normal adjacent tissues, cell lines | PDCD4 | [73] |
| suppressor | down | 16 tumor tissues and matched adjacent normal tissues, cell lines | Gli2 | [69] |
| suppressor | down | 6 metastatic OS and non-metastatic OS | CALD1, STX1A | [71] |
| suppressor | down | 32 OS without metastasis and 24 w lung metastases, 30 controls, cell lines | MALAT1 | [72] |
| mir-202 Function | Regulation | Samples, Cell Lines, Patient Material, Animal Model | Involved Downstream/Upstream Molecules | Ref. |
|---|---|---|---|---|
| Chronic myeloid leukemia | ||||
| pro-tumorigenic | up | 30 patient samples, cells lines, xenografts of nude mice | STAT5A/miR-202-5p/USP15/Caspase-6 regulatory axis | [82] |
| suppressor | down | 15 CML samples, cell lines | HK2 | [83] |
| Follicular lymphoma | ||||
| suppressor | down | cell lines | HAS2, FAM135A | [81] |
| Glioma | ||||
| suppressor | down | 43 glioma samples and adjacent normal tissues, cell lines | MTDH, PI3K/Akt, Wnt/β-catenin pathways | [80] |
| Laryngeal cancer | ||||
| pro-tumorigenic | up | 48 laryngeal primary tumors | n.d. | [84] |
| Multiple myeloma | ||||
| pro-tumorigenic | up | cell lines | BAFF, JNK/SAPK signaling pathway | [76] |
| pro-tumorigenic | up | 40 MM patient sera, 30 healthy controls | BAFF | [5] |
| suppressor | down | cell lines | BAFF, JNK/SAPK signaling pathway | [77,79] |
| suppressor | down | 7 MM patient samples, cell lines | BAFF | [85] |
| Thyroid carcinoma | ||||
| suppressor | down | 40 PTC and adjacent normal tissues | lncRNA NORAD, EMT | [75] |
| suppressor | down | 96 pairs of PTC and adjacent normal tissue, cell lines | β-catenin, WNT signaling | [67] |
| Glioblastoma multiforme | ||||
| suppressor | down | cell lines | IR-induced K-RAS/ERK signaling, CD44 | [86] |
| Neuroblastoma | ||||
| suppressor | down | cell lines | MYCN | [87] |
| suppressor | down | cell lines | MYKN, E2F1 | [88] |
| Urinary Bladder cancer | ||||
| suppressor | down | 50 cancer and corresponding tissues, cell lines | EGFR | [89] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, E.A.; Rajendran, P.; Scherthan, H. The microRNA-202 as a Diagnostic Biomarker and a Potential Tumor Suppressor. Int. J. Mol. Sci. 2022, 23, 5870. https://doi.org/10.3390/ijms23115870
Ahmed EA, Rajendran P, Scherthan H. The microRNA-202 as a Diagnostic Biomarker and a Potential Tumor Suppressor. International Journal of Molecular Sciences. 2022; 23(11):5870. https://doi.org/10.3390/ijms23115870
Chicago/Turabian StyleAhmed, Emad A., Peramaiyan Rajendran, and Harry Scherthan. 2022. "The microRNA-202 as a Diagnostic Biomarker and a Potential Tumor Suppressor" International Journal of Molecular Sciences 23, no. 11: 5870. https://doi.org/10.3390/ijms23115870
APA StyleAhmed, E. A., Rajendran, P., & Scherthan, H. (2022). The microRNA-202 as a Diagnostic Biomarker and a Potential Tumor Suppressor. International Journal of Molecular Sciences, 23(11), 5870. https://doi.org/10.3390/ijms23115870