
Light Induces Carotenoid Biosynthesis-Related Gene Expression, Accumulation of Pigment Content, and Expression of the Small Heat Shock Protein in Apple Fruit



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
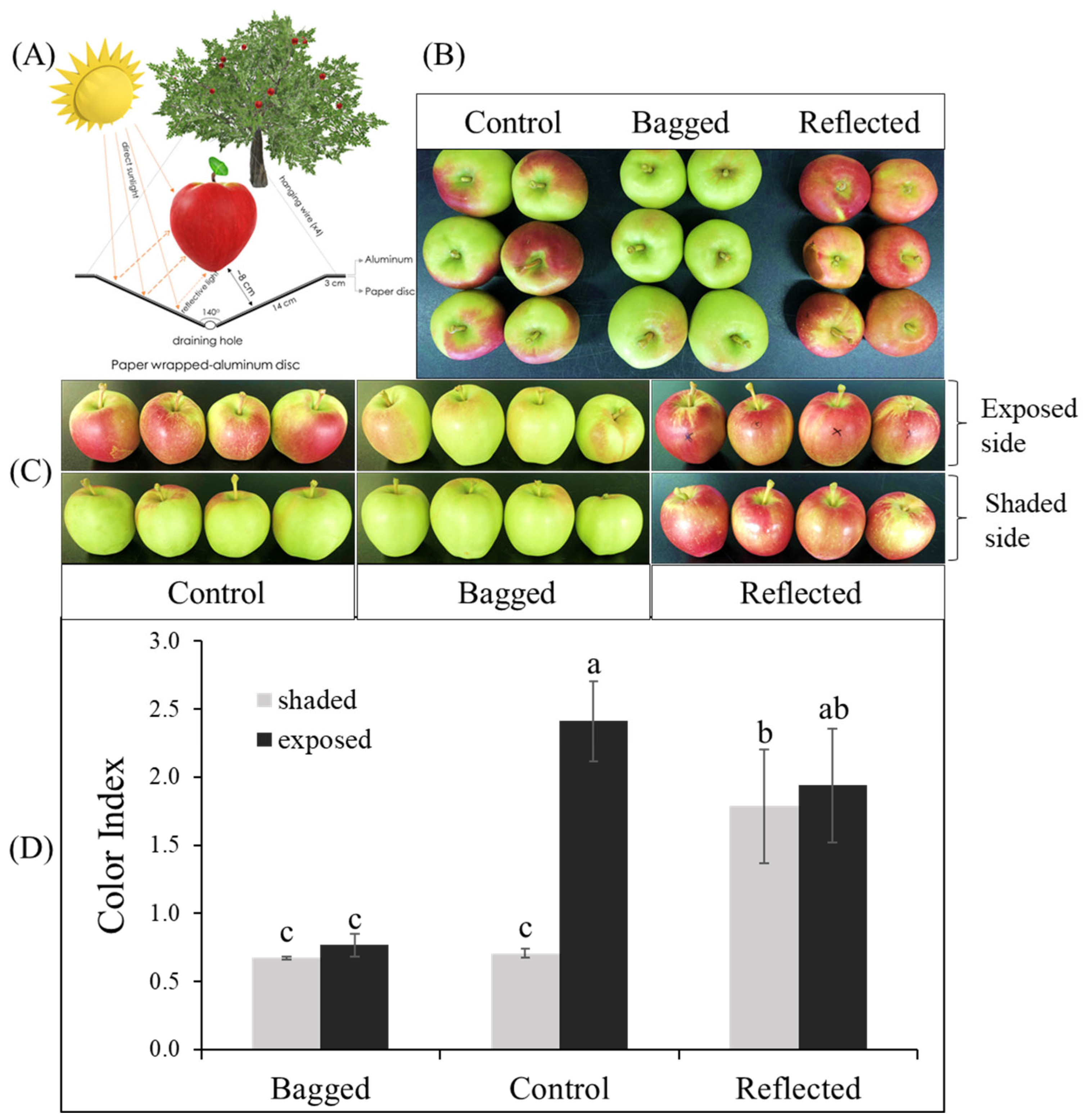
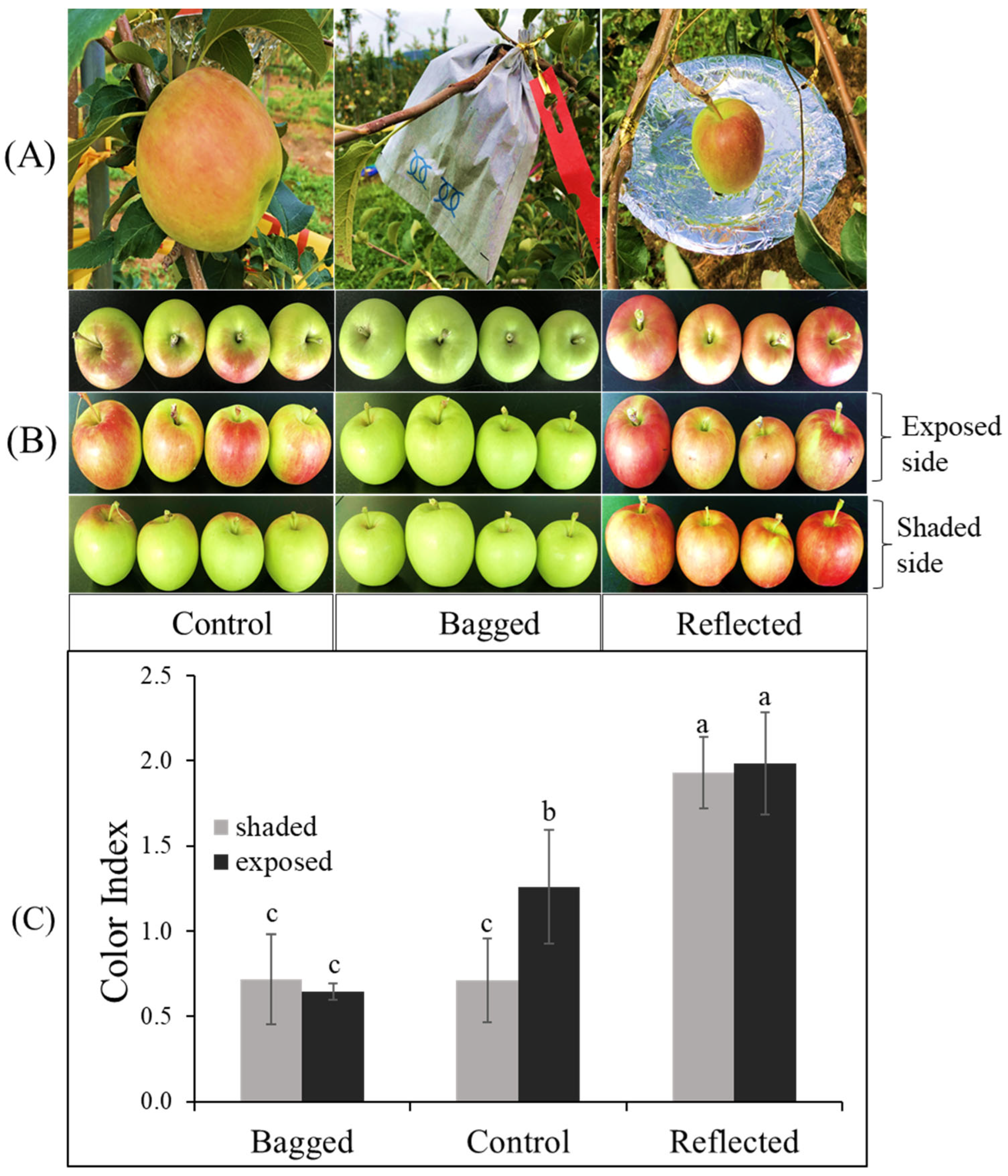
2.1. Different Coloration Patterns of Apple Fruit under Light Treatment
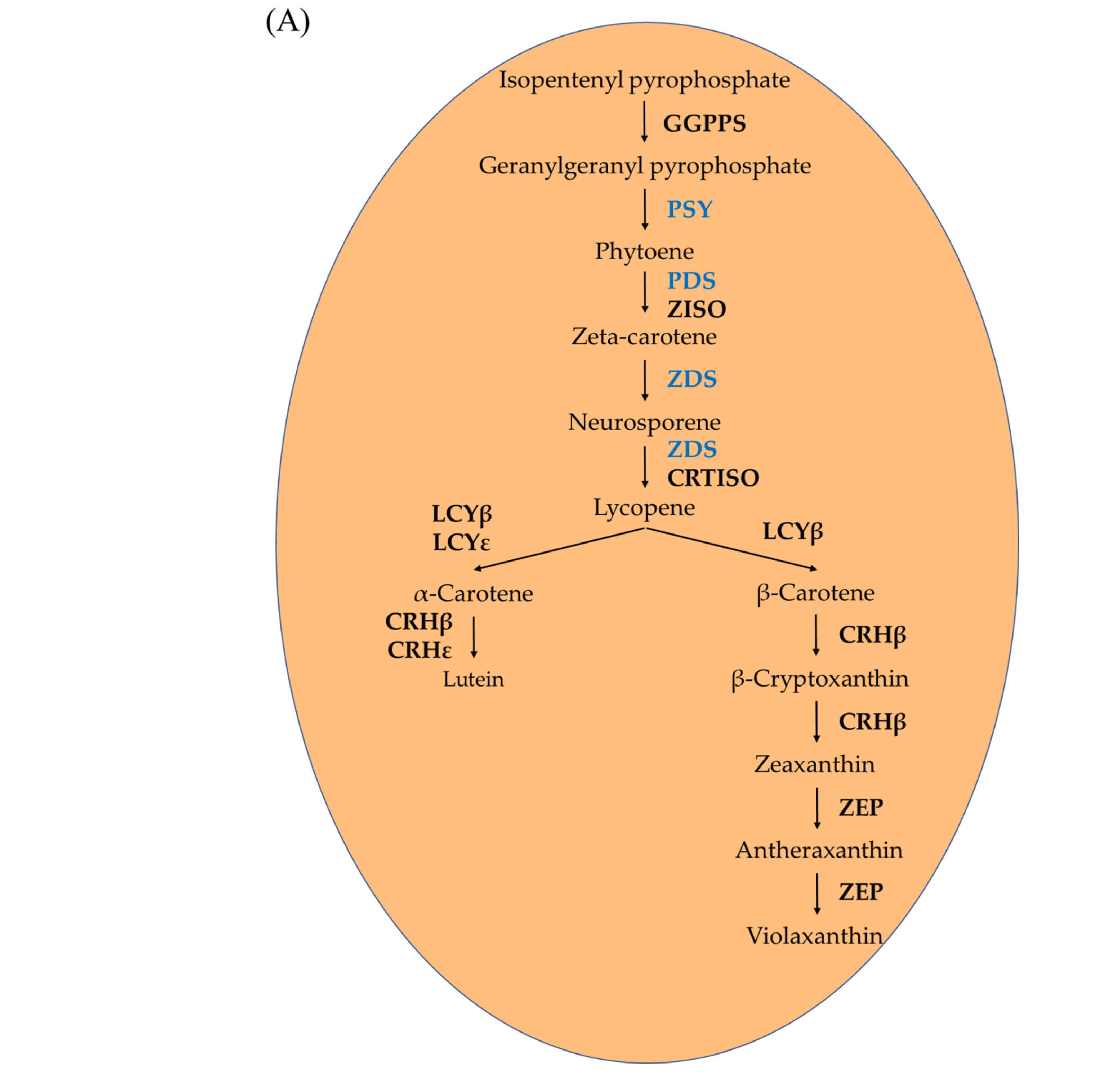
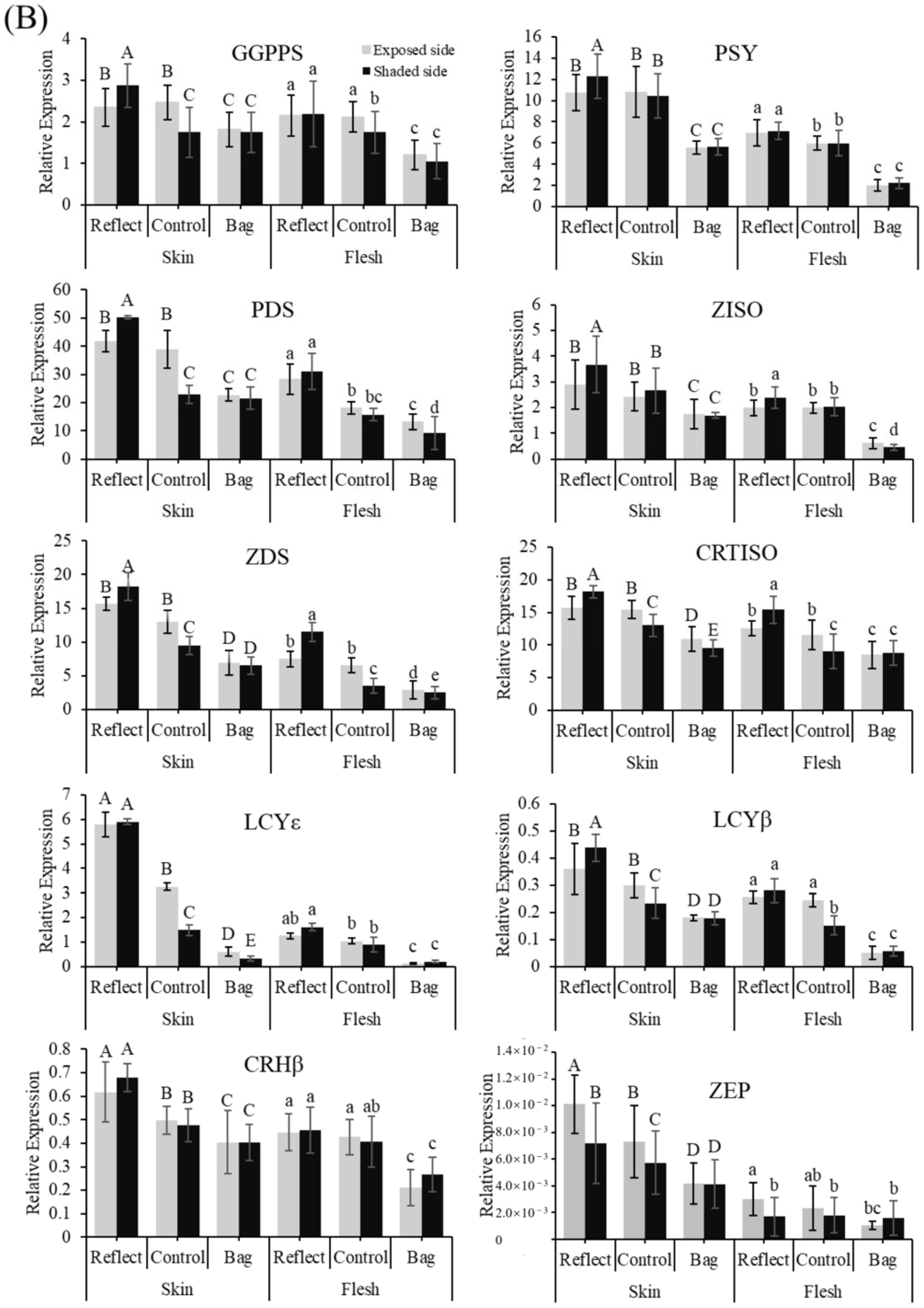
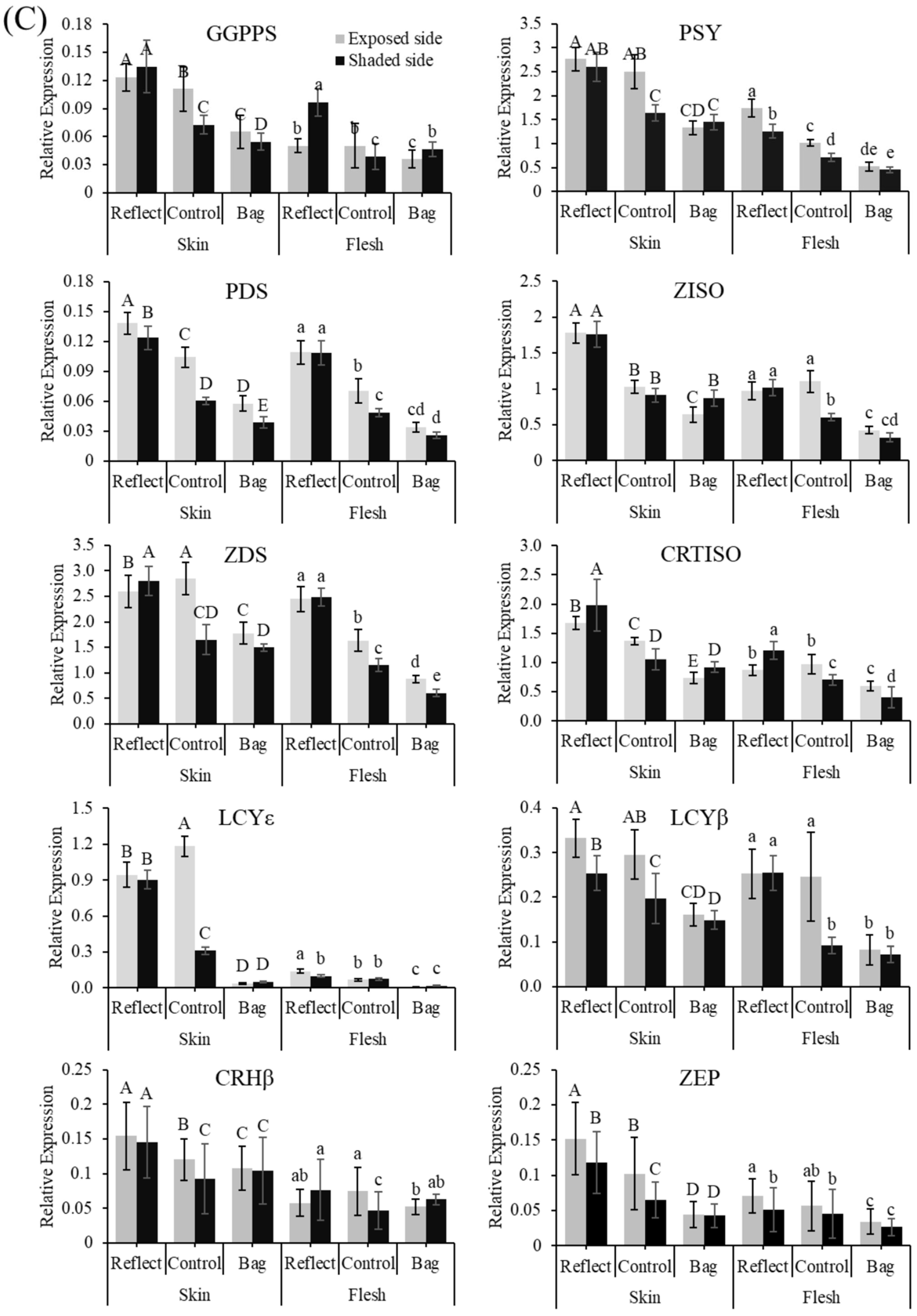
2.2. Expression Profiles of the Carotenoid Biosynthesis-Related Genes under Light Treatment
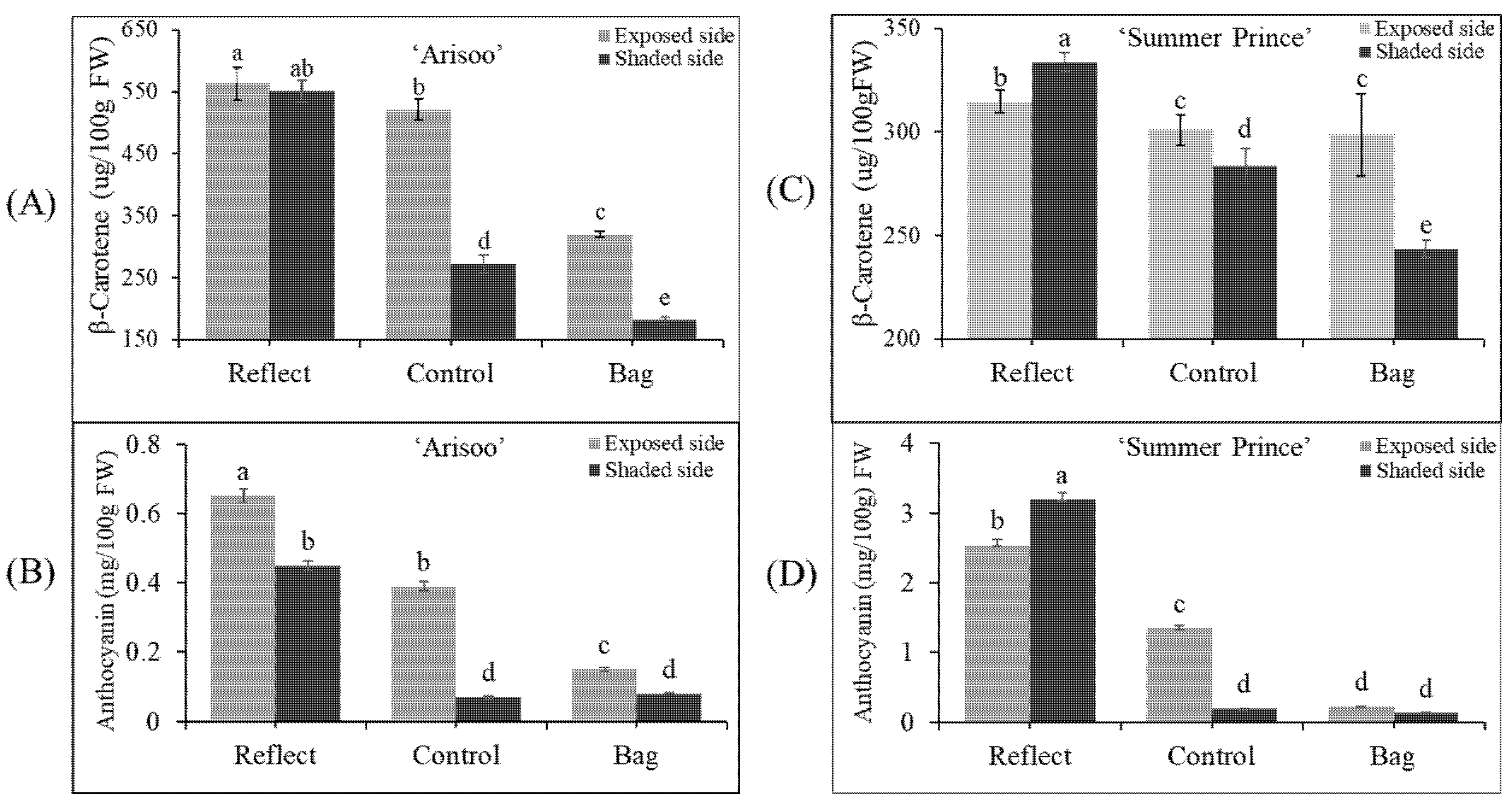
2.3. Accumulation of Pigment Content under Light Treatment
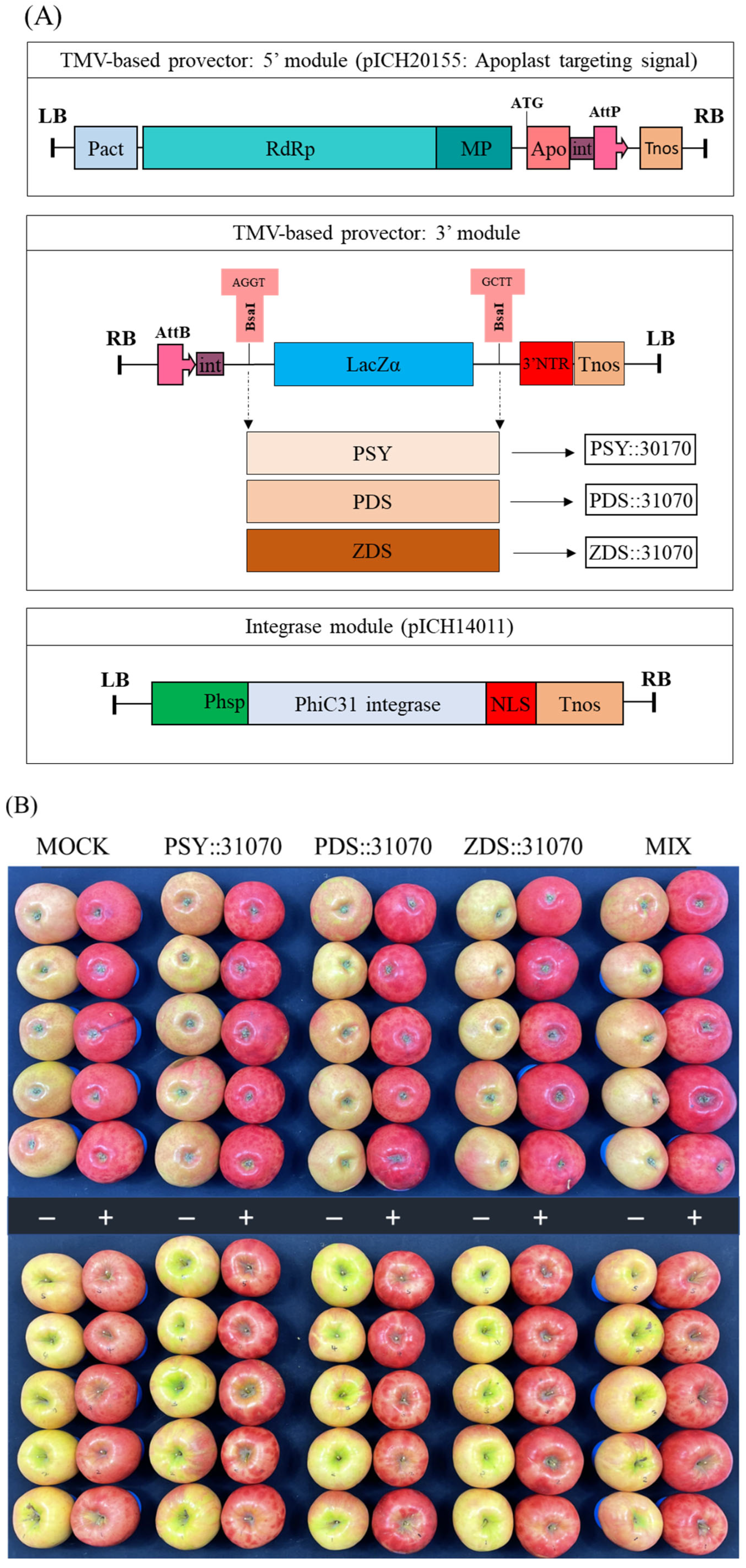
2.4. Construction of Expression Vectors
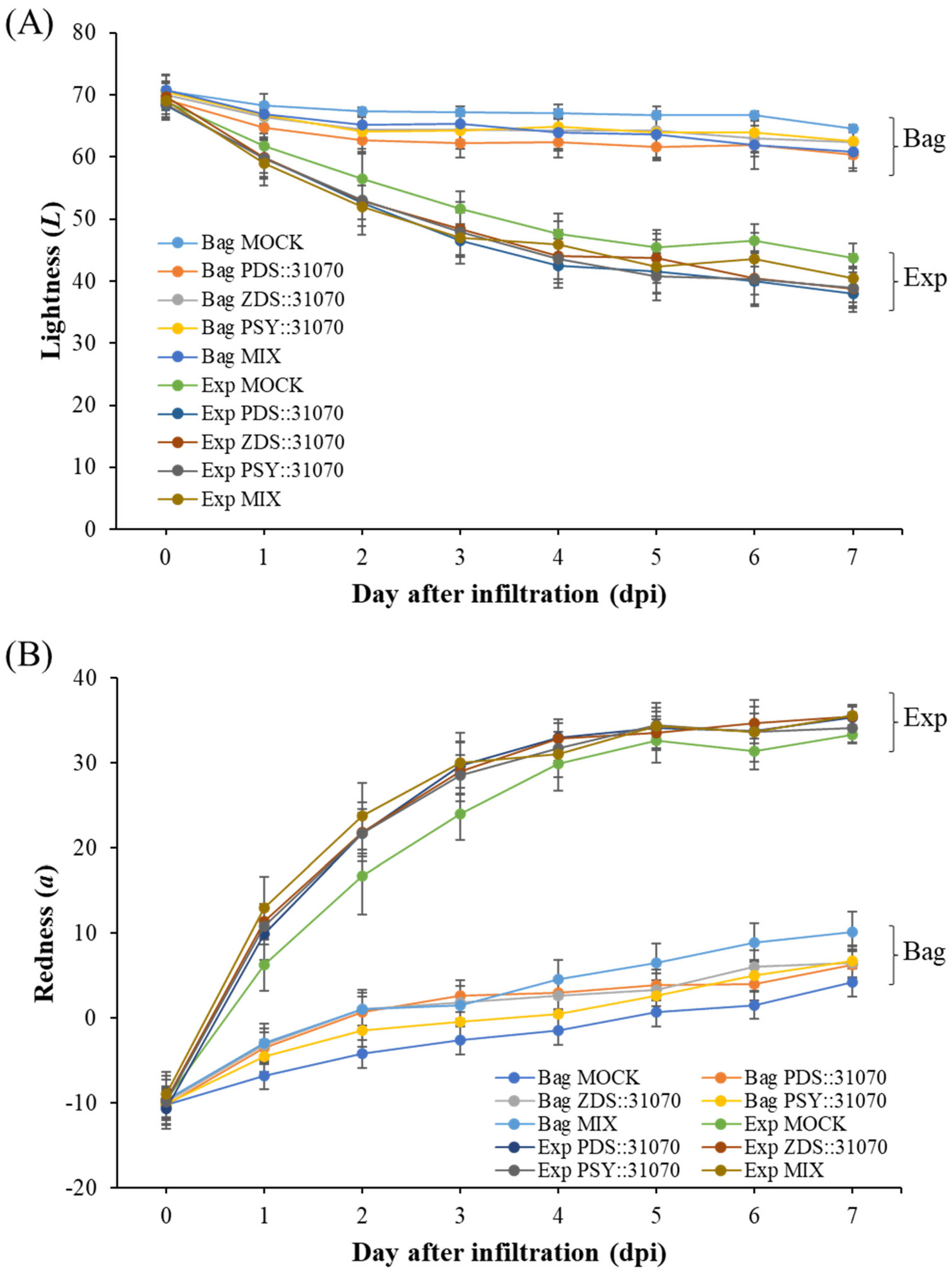
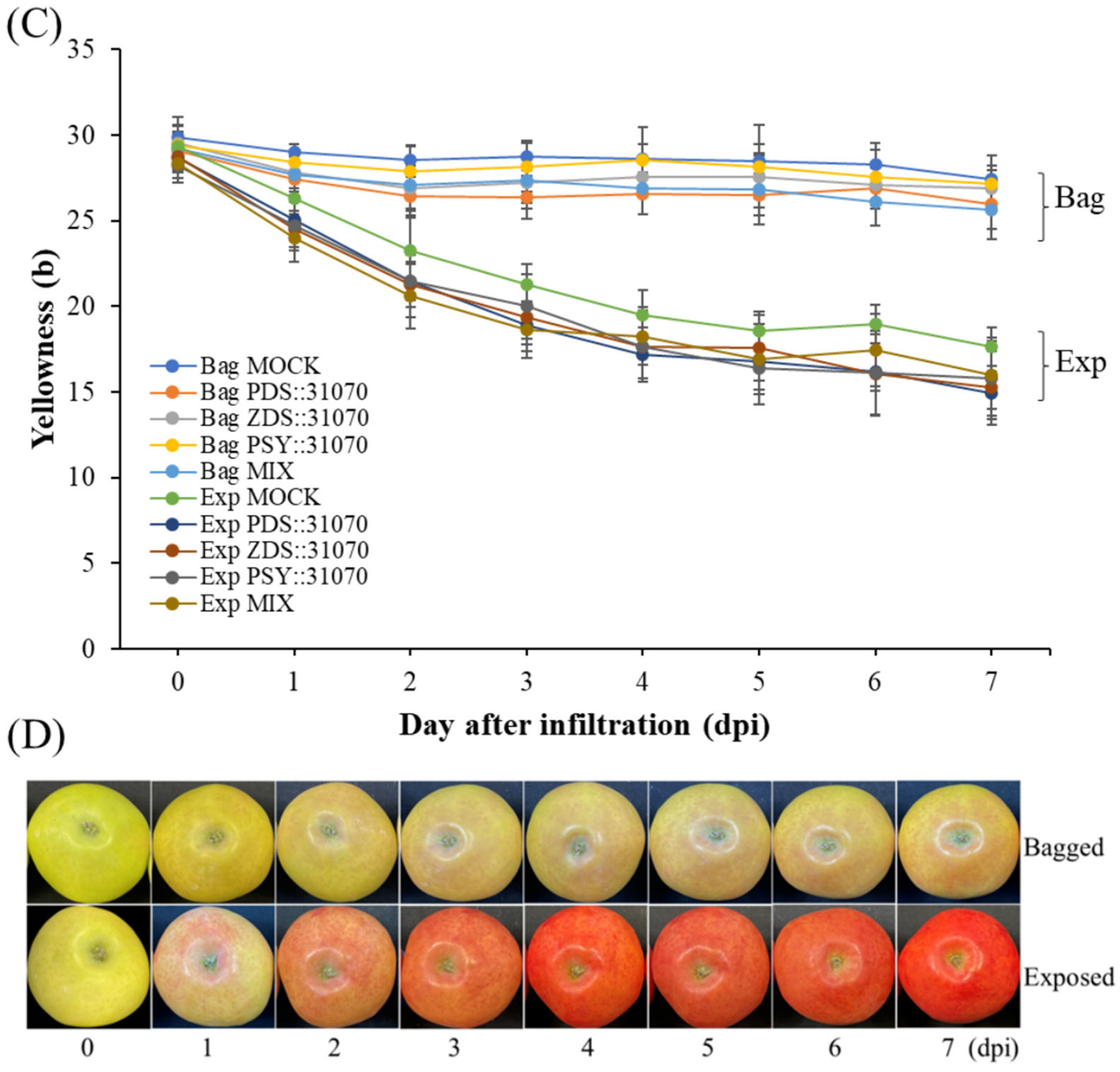
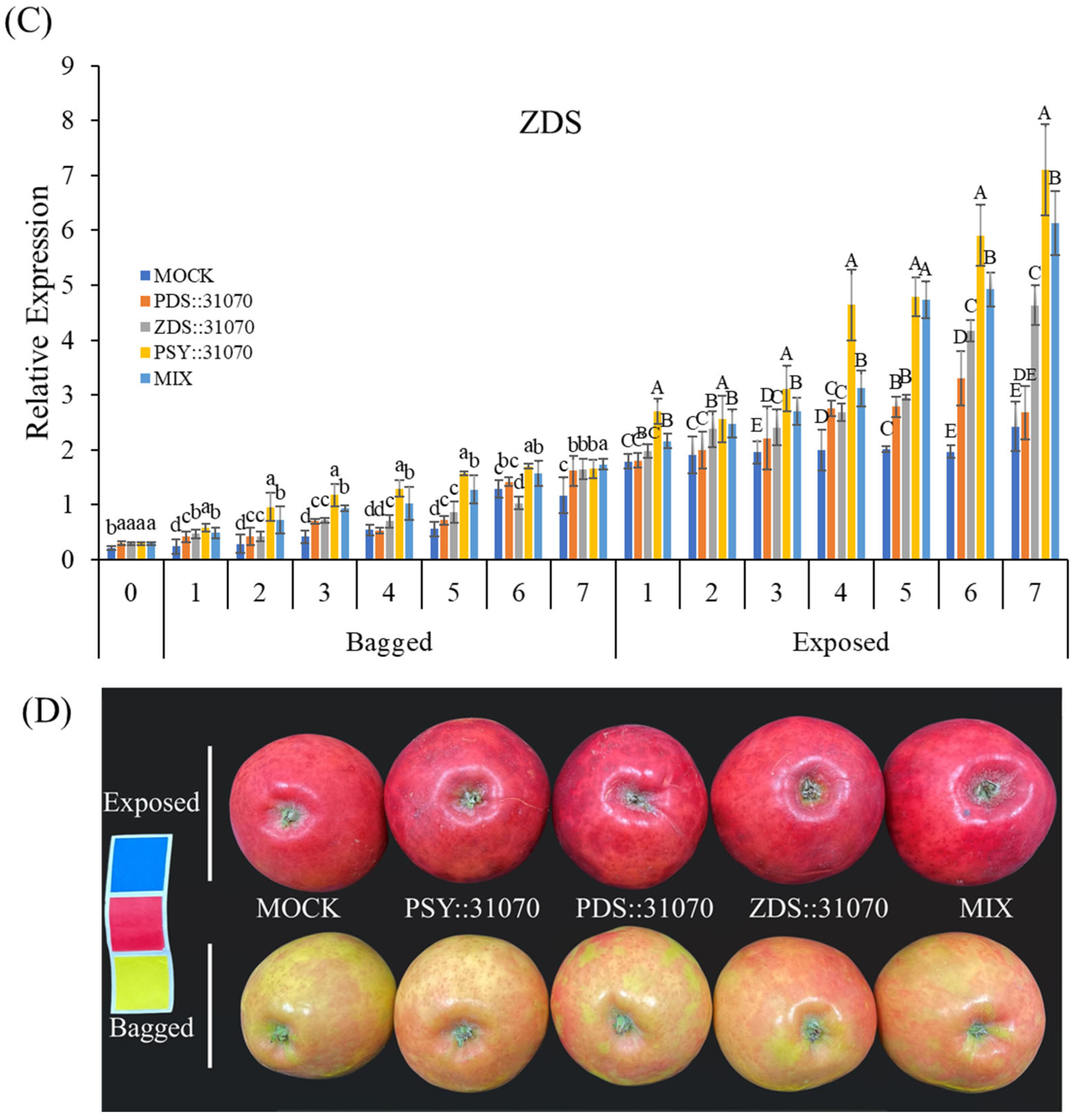
2.5. Effect of Light Treatment and Transient Expression of Carotenogenic Genes on Apple Fruit Coloration
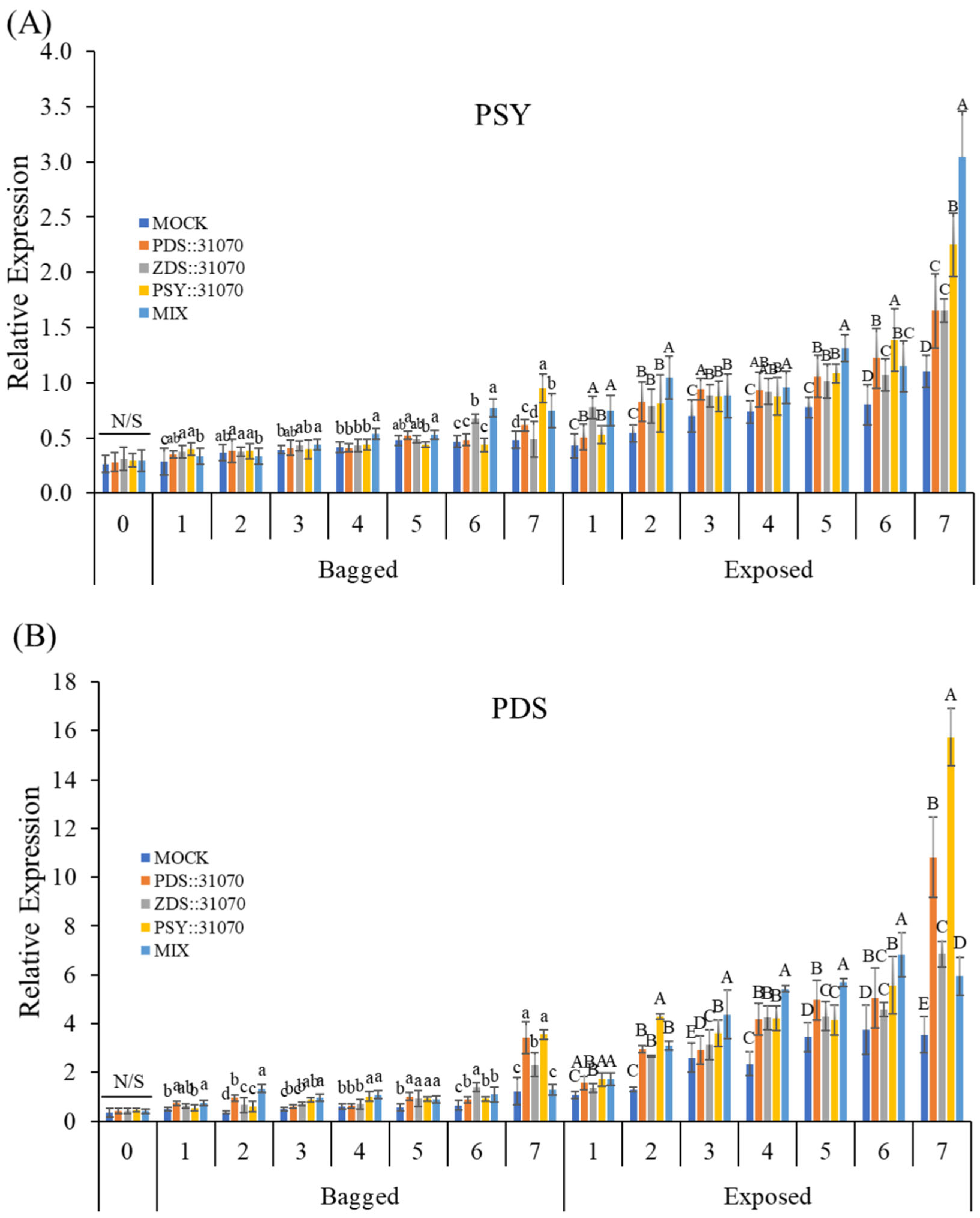
2.6. Expression Profiles of the Carotenogenic Genes under Light Treatment and Transient Expression
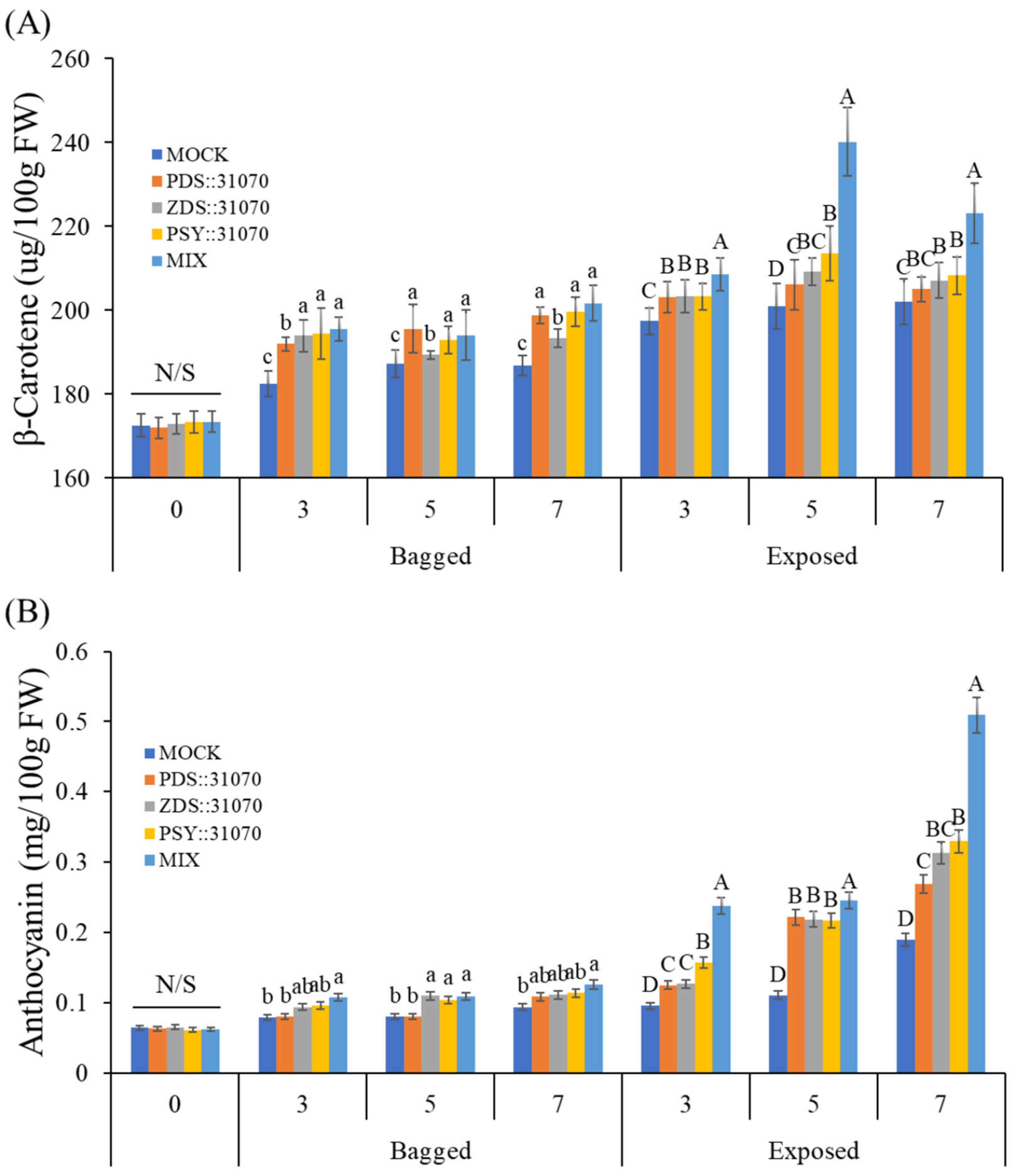
2.7. Accumulation of Pigment Content under Light Treatment and Transient Expression
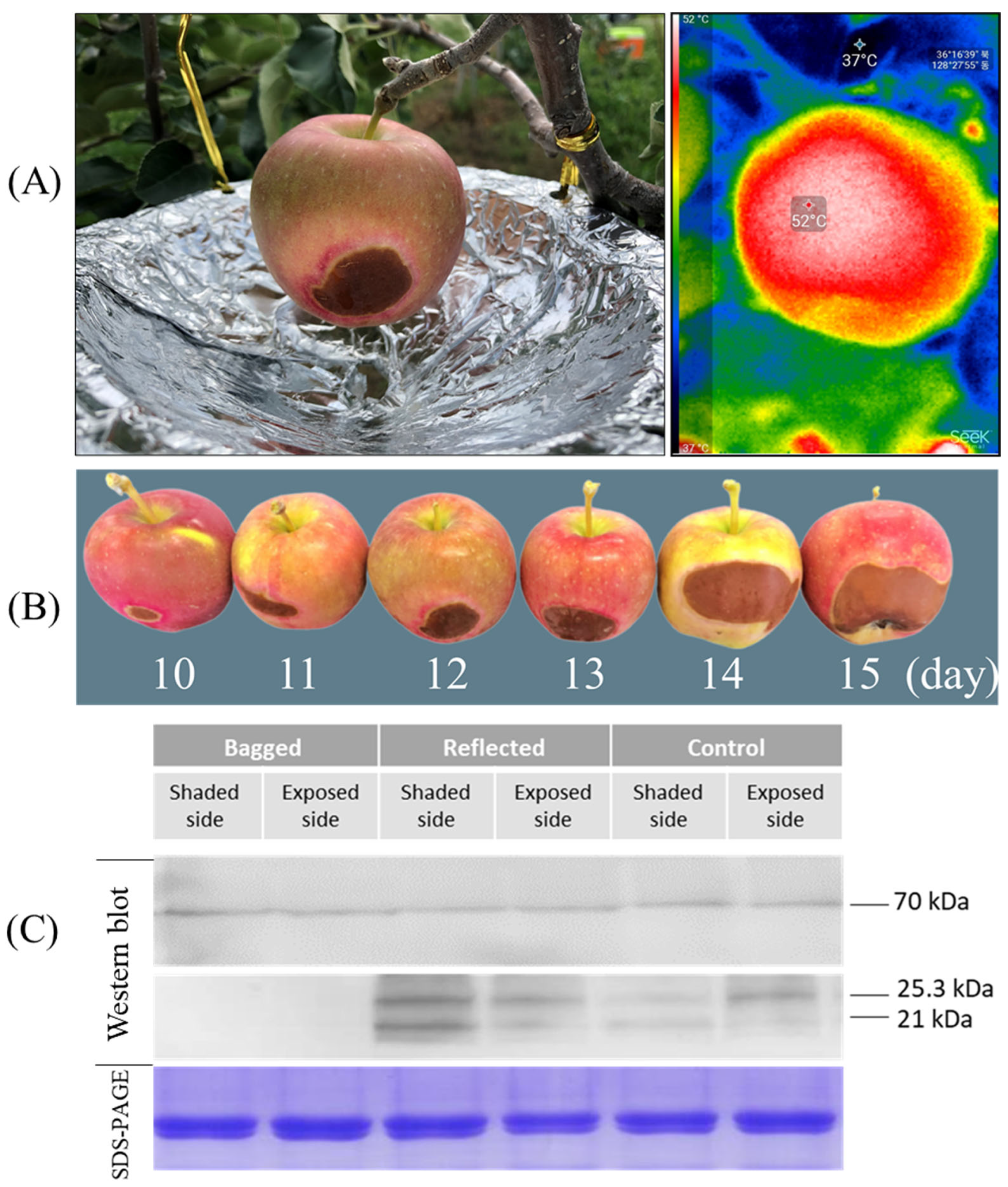
2.8. Accumulation of Small Heat Shock Proteins (smHSPs) in Response to Heat Stress
3. Materials and Methods
3.1. Fruit Materials and Sunlight Treatments
3.2. Measurements of Fruit Color and Fruit Characteristics
3.3. Construction of Plant Expression Vectors and Agrobacterium Transformation
3.4. Agrobacterium Vacuum-Infiltration
3.5. RNA Extraction and Quantification of Gene Expression
3.6. Pigment Content Analysis
3.6.1. Carotenoid Analysis
3.6.2. Anthocyanin Analysis
3.7. Heat Treatment and Protein Extraction
3.8. SDS-PAGE and Western Blot Analysis
3.9. Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nishino, H.; Tokuda, H.; Satomi, Y.; Masuda, M.; Bu, P.; Onozuka, M.; Yamaguchi, S.; Okuda, Y.; Takayasu, J.; Tsuruta, J. Cancer prevention by carotenoids. Pure Appl. Chem. 1999, 71, 2273–2278. [Google Scholar] [CrossRef]
- Paolini, M.; Abdel-Rahman, S.Z.; Sapone, A.; Pedulli, G.F.; Perocco, P.; Cantelli-Forti, G.; Legator, M.S. β-Carotene: A cancer chemopreventive agent or a co-carcinogen? Mutat. Res. Rev. Mutat. Res. 2003, 543, 195–200. [Google Scholar] [CrossRef]
- Voutilainen, S.; Nurmi, T.; Mursu, J.; Rissanen, T.H. Carotenoids and cardiovascular health. Am. J. Clin. Nutr. 2006, 83, 1265–1271. [Google Scholar] [CrossRef] [PubMed]
- Sesso, H.D. Carotenoids and cardiovascular disease: What research gaps remain? Curr. Opin. Lipidol. 2006, 17, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Meléndez-Martínez, A.J.; Böhm, V.; Borge, G.I.A.; Cano, M.P.; Fikselová, M.; Gruskiene, R.; Lavelli, V.; Loizzo, M.R.; Mandić, A.I.; Brahm, P.M. Carotenoids: Considerations for their use in functional foods, nutraceuticals, nutricosmetics, supplements, botanicals, and novel foods in the context of sustainability, circular economy, and climate change. Annu. Rev. Food Sci. Technol. 2021, 12, 433–460. [Google Scholar] [CrossRef]
- Khachik, F. Distribution and metabolism of dietary carotenoids in humans as a criterion for development of nutritional supplements. Pure Appl. Chem. 2006, 78, 1551–1557. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J. An overview of carotenoids, apocarotenoids, and vitamin A in agro-food, nutrition, health, and disease. Mol. Nutr. Food Res. 2019, 63, 1801045. [Google Scholar] [CrossRef] [Green Version]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef]
- Cunningham Jr, F.X.; Gantt, E. Genes and enzymes of carotenoid biosynthesis in plants. Annu. Rev. Plant Biol. 1998, 49, 557–583. [Google Scholar] [CrossRef]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid metabolism and regulation in horticultural crops. Hortic. Res. 2015, 2, 15036. [Google Scholar] [CrossRef] [Green Version]
- Ampomah-Dwamena, C.; Dejnoprat, S.; Lewis, D.; Sutherland, P.; Volz, R.K.; Allan, A.C. Metabolic and gene expression analysis of apple (Malus× domestica) carotenogenesis. J. Exp. Bot. 2012, 63, 4497–4511. [Google Scholar] [CrossRef] [PubMed]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Al-Babili, S.; Kloti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the provitamin A (β-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 2000, 287, 303–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, P.D.; Truesdale, M.R.; Bird, C.R.; Schuch, W.; Bramley, P.M. Carotenoid biosynthesis during tomato fruit development (evidence for tissue-specific gene expression). Plant Physiol. 1994, 105, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Lois, L.M.; Rodríguez-Concepción, M.; Gallego, F.; Campos, N.; Boronat, A. Carotenoid biosynthesis during tomato fruit development: Regulatory role of 1-deoxy-D-xylulose 5-phosphate synthase. Plant J. 2000, 22, 503–513. [Google Scholar] [CrossRef]
- Hugueney, P.; Bouvier, F.; Badillo, A.; Quennemet, J.; d’Harlingue, A.; Camara, B. Developmental and stress regulation of gene expression for plastid and cytosolic isoprenoid pathways in pepper fruits. Plant Physiol. 1996, 111, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Fraser, P.D.; Romer, S.; Shipton, C.A.; Mills, P.B.; Kiano, J.W.; Misawa, N.; Drake, R.G.; Schuch, W.; Bramley, P.M. Evaluation of transgenic tomato plants expressing an additional phytoene synthase in a fruit-specific manner. Proc. Natl. Acad. Sci. USA 2002, 99, 1092–1097. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Vallabhaneni, R.; Yu, J.; Rocheford, T.; Wurtzel, E.T. The maize phytoene synthase gene family: Overlapping roles for carotenogenesis in endosperm, photomorphogenesis, and thermal stress tolerance. Plant Physiol. 2008, 147, 1334–1346. [Google Scholar] [CrossRef] [Green Version]
- Welsch, R.; Wust, F.; Bar, C.; Al-Babili, S.; Beyer, P. A third phytoene synthase is devoted to abiotic stress-induced abscisic acid formation in rice and defines functional diversification of phytoene synthase genes. Plant Physiol. 2008, 147, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.-J.; Jeong, H.-B.; Jung, A.; Kang, M.-Y.; Kim, S.; Ha, S.-H.; Kwon, J.-K.; Kang, B.-C. Phytoene synthase 2 can compensate for the absence of PSY1 in the control of color in Capsicum fruit. J. Exp. Bot. 2020, 71, 3417–3427. [Google Scholar] [CrossRef]
- Ji, J.; Wang, G.; Wang, J.; Wang, P. Functional analysis of multiple carotenogenic genes from Lycium barbarum and Gentiana lutea L. for their effects on β-carotene production in transgenic tobacco. Biotechnol. Lett. 2009, 31, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Ampomah-Dwamena, C.; Driedonks, N.; Lewis, D.; Shumskaya, M.; Chen, X.; Wurtzel, E.T.; Espley, R.V.; Allan, A.C. The Phytoene synthase gene family of apple (Malus x domestica) and its role in controlling fruit carotenoid content. BMC Plant Biol. 2015, 15, 185. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-J.; Gao, Y.; Wen, P.-F.; Hao, Y.-Y.; Chen, X.-X. Cloning and expression analysis of the phytoene synthase gene in ‘Granny Smith’apple (Malus× domestica Borkh.). Biotechnol. Biotechnol. Equip. 2018, 32, 1105–1112. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Ji, J.; Wang, G.; Li, Z.; Wang, Y.; Fan, Y. Over-expression of LcPDS, LcZDS, and LcCRTISO, genes from wolfberry for carotenoid biosynthesis, enhanced carotenoid accumulation, and salt tolerance in tobacco. Front. Plant Sci. 2020, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Welsch, R.; Yang, Y.; Álvarez, D.; Riediger, M.; Yuan, H.; Fish, T.; Liu, J.; Thannhauser, T.W.; Li, L. Arabidopsis OR proteins are the major posttranscriptional regulators of phytoene synthase in controlling carotenoid biosynthesis. Proc. Natl. Acad. Sci. USA 2015, 112, 3558–3563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Kim, H.S.; Jung, Y.J.; Kim, S.H.; Ji, C.Y.; Wang, Z.; Jeong, J.C.; Lee, H.-S.; Lee, S.Y.; Kwak, S.-S. Orange protein has a role in phytoene synthase stabilization in sweetpotato. Sci. Rep. 2016, 6, 33563. [Google Scholar] [CrossRef]
- Lu, S.; Zhang, Y.; Zhu, K.; Yang, W.; Ye, J.; Chai, L.; Xu, Q.; Deng, X. The citrus transcription factor CsMADS6 modulates carotenoid metabolism by directly regulating carotenogenic genes. Plant Physiol. 2018, 176, 2657–2676. [Google Scholar] [CrossRef] [Green Version]
- Llorente, B.; D’Andrea, L.; Rodríguez-Concepción, M. Evolutionary recycling of light signaling components in fleshy fruits: New insights on the role of pigments to monitor ripening. Front. Plant Sci. 2016, 7, 263. [Google Scholar] [CrossRef] [Green Version]
- Meléndez-Martínez, A.J.; Mandić, A.I.; Bantis, F.; Böhm, V.; Borge, G.I.A.; Brnčić, M.; Bysted, A.; Cano, M.P.; Dias, M.G.; Elgersma, A. A comprehensive review on carotenoids in foods and feeds: Status quo, applications, patents, and research needs. Crit. Rev. Food Sci. Nutr. 2022, 62, 1999–2049. [Google Scholar] [CrossRef]
- Llorente, B.; Martinez-Garcia, J.F.; Stange, C.; Rodriguez-Concepcion, M. Illuminating colors: Regulation of carotenoid biosynthesis and accumulation by light. Curr. Opin. Plant Biol. 2017, 37, 49–55. [Google Scholar] [CrossRef]
- Domonkos, I.; Kis, M.; Gombos, Z.; Ughy, B. Carotenoids, versatile components of oxygenic photosynthesis. Prog. Lipid Res. 2013, 52, 539–561. [Google Scholar] [CrossRef] [PubMed]
- Llorente, B.; D’Andrea, L.; Ruiz-Sola, M.A.; Botterweg, E.; Pulido, P.; Andilla, J.; Loza-Alvarez, P.; Rodriguez-Concepcion, M. Tomato fruit carotenoid biosynthesis is adjusted to actual ripening progression by a light-dependent mechanism. Plant J. 2016, 85, 107–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledo-Ortiz, G.; Huq, E.; Rodríguez-Concepción, M. Direct regulation of phytoene synthase gene expression and carotenoid biosynthesis by phytochrome-interacting factors. Proc. Natl. Acad. Sci. USA 2010, 107, 11626–11631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Fang, W.; Chen, C.; Wang, L.; Cao, K. Effects of Shading by Bagging on Carotenoid Accumulation in Peach Fruit Flesh. J. Plant Growth Regul. 2021, 40, 1912–1921. [Google Scholar] [CrossRef]
- Van Giap, D.; Kim, S.; Lee, Y.; Kweon, H.-J. Effect of Reflected Sunlight on Differential Expression of Anthocyanin Synthesis-Related Genes in Young Apple Fruit. Int. J. Fruit Sci. 2021, 21, 440–455. [Google Scholar] [CrossRef]
- Barua, D.; Downs, C.A.; Heckathorn, S.A. Variation in chloroplast small heat-shock protein function is a major determinant of variation in thermotolerance of photosynthetic electron transport among ecotypes of Chenopodium album. Funct. Plant Biol. 2003, 30, 1071–1079. [Google Scholar] [CrossRef]
- Hu, S.; Ding, Y.; Zhu, C. Sensitivity and responses of chloroplasts to heat stress in plants. Front. Plant Sci. 2020, 11, 375. [Google Scholar] [CrossRef] [Green Version]
- Zhong, L.; Zhou, W.; Wang, H.; Ding, S.; Lu, Q.; Wen, X.; Peng, L.; Zhang, L.; Lu, C. Chloroplast small heat shock protein HSP21 interacts with plastid nucleoid protein pTAC5 and is essential for chloroplast development in Arabidopsis under heat stress. Plant Cell 2013, 25, 2925–2943. [Google Scholar] [CrossRef] [Green Version]
- Ritenour, M.A.; Kochhar, S.; Schrader, L.E.; Hsu, T.-P.; Ku, M.S. Characterization of heat shock protein expression in apple peel under field and laboratory conditions. J. Am. Soc. Hortic. Sci. 2001, 126, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Che, F.; Wang, L.; Meng, R.; Zhang, X.; Zhao, Z. Fruit coloration and anthocyanin biosynthesis after bag removal in non-red and red apples (Malus× domestica Borkh.). Molecules 2013, 18, 1549–1563. [Google Scholar] [CrossRef]
- Sun, S.; Xin, L.; Gao, H.; Wang, J.; Li, P. Response of phenolic compounds in ‘Golden Delicious’ and ‘Red Delicious’ apples peel to fruit bagging and subsequent sunlight re-exposure. Sci. Hortic. 2014, 168, 161–167. [Google Scholar] [CrossRef]
- Chen, C.-S.; Zhang, D.; Wang, Y.-Q.; Li, P.-M.; Ma, F.-W. Effects of fruit bagging on the contents of phenolic compounds in the peel and flesh of ‘Golden Delicious’,‘Red Delicious’, and ‘Royal Gala’apples. Sci. Hortic. 2012, 142, 68–73. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, M.; Zhang, G.; Li, P.; Ma, F. Differential regulation of anthocyanin synthesis in apple peel under different sunlight intensities. Int. J. Mol. Sci. 2019, 20, 6060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Y.; Ren, H.; Guo, P. Effects of bagging on the accumulation and transformation of photosynthates in apple fruits. Acta Hortic. Sin. 2011, 38, 233–239. [Google Scholar]
- Shaomin, W.; Huajun, G.; Xiaobing, Z. Effects of bagging on pigment, sugar and acid development in ‘Red Fuji’apple fruits. Acta Hortic. Sin. 2002, 29, 263. [Google Scholar]
- Gleba, Y.Y.; Giritch, A. Plant viral vectors for protein expression. Recent Adv. Plant Virol. 2011, 387–412. [Google Scholar] [CrossRef]
- Cao, H.; Wang, J.; Dong, X.; Han, Y.; Ma, Q.; Ding, Y.; Zhao, F.; Zhang, J.; Chen, H.; Xu, Q. Carotenoid accumulation affects redox status, starch metabolism, and flavonoid/anthocyanin accumulation in citrus. BMC Plant Biol. 2015, 15, 27. [Google Scholar] [CrossRef] [Green Version]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. Am. J. Enol. Vitic. 2002, 53, 171–182. [Google Scholar]
- Tsai, M.-S.; Lee, T.-C.; Chang, P.-T. Comparison of paper bags, calcium carbonate, and shade nets for sunscald protection in ‘Murcott’tangor fruit. HortTechnology 2013, 23, 659–667. [Google Scholar] [CrossRef]
- DeRocher, A.E.; Helm, K.W.; Lauzon, L.M.; Vierling, E. Expression of a conserved family of cytoplasmic low molecular weight heat shock proteins during heat stress and recovery. Plant Physiol. 1991, 96, 1038–1047. [Google Scholar] [CrossRef] [Green Version]
- Holsters, M.; De Waele, D.; Depicker, A.; Messens, E.; Van Montagu, M.; Schell, J. Transfection and transformation of Agrobacterium tumefaciens. Mol. Gen. Genet. MGG 1978, 163, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, K.; Li, J.; Huang, H.; Que, Q.; Li, P.; Chen, X. A simple method for RNA isolation from various tissues of the tree Neolamarckia cadamba. Biotechnol. Biotechnol. Equip. 2014, 28, 1008–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Cong, P.; Tian, Y.; Zhu, Y. Using RNA-seq data to select reference genes for normalizing gene expression in apple roots. PLoS ONE 2017, 12, e0185288. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zhang, M.; Li, X. Comparisons of extraction solvents and quantitative-methods for analysis of anthocyanins in strawberry and blueberry fruits. Acta Hortic. Sin. 2008, 35, 655–660. [Google Scholar]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Do, V.G.; Lee, Y.; Kweon, H.; Kim, S. Light Induces Carotenoid Biosynthesis-Related Gene Expression, Accumulation of Pigment Content, and Expression of the Small Heat Shock Protein in Apple Fruit. Int. J. Mol. Sci. 2022, 23, 6153. https://doi.org/10.3390/ijms23116153
Do VG, Lee Y, Kweon H, Kim S. Light Induces Carotenoid Biosynthesis-Related Gene Expression, Accumulation of Pigment Content, and Expression of the Small Heat Shock Protein in Apple Fruit. International Journal of Molecular Sciences. 2022; 23(11):6153. https://doi.org/10.3390/ijms23116153
Chicago/Turabian StyleDo, Van Giap, Youngsuk Lee, Hunjoong Kweon, and Seonae Kim. 2022. "Light Induces Carotenoid Biosynthesis-Related Gene Expression, Accumulation of Pigment Content, and Expression of the Small Heat Shock Protein in Apple Fruit" International Journal of Molecular Sciences 23, no. 11: 6153. https://doi.org/10.3390/ijms23116153
APA StyleDo, V. G., Lee, Y., Kweon, H., & Kim, S. (2022). Light Induces Carotenoid Biosynthesis-Related Gene Expression, Accumulation of Pigment Content, and Expression of the Small Heat Shock Protein in Apple Fruit. International Journal of Molecular Sciences, 23(11), 6153. https://doi.org/10.3390/ijms23116153