
Antifungal Activity of the Frog Skin Peptide Temporin G and Its Effect on Candida albicans Virulence Factors

 ,
,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
2.1. TG Exerts Activity on Different Fungal Strains
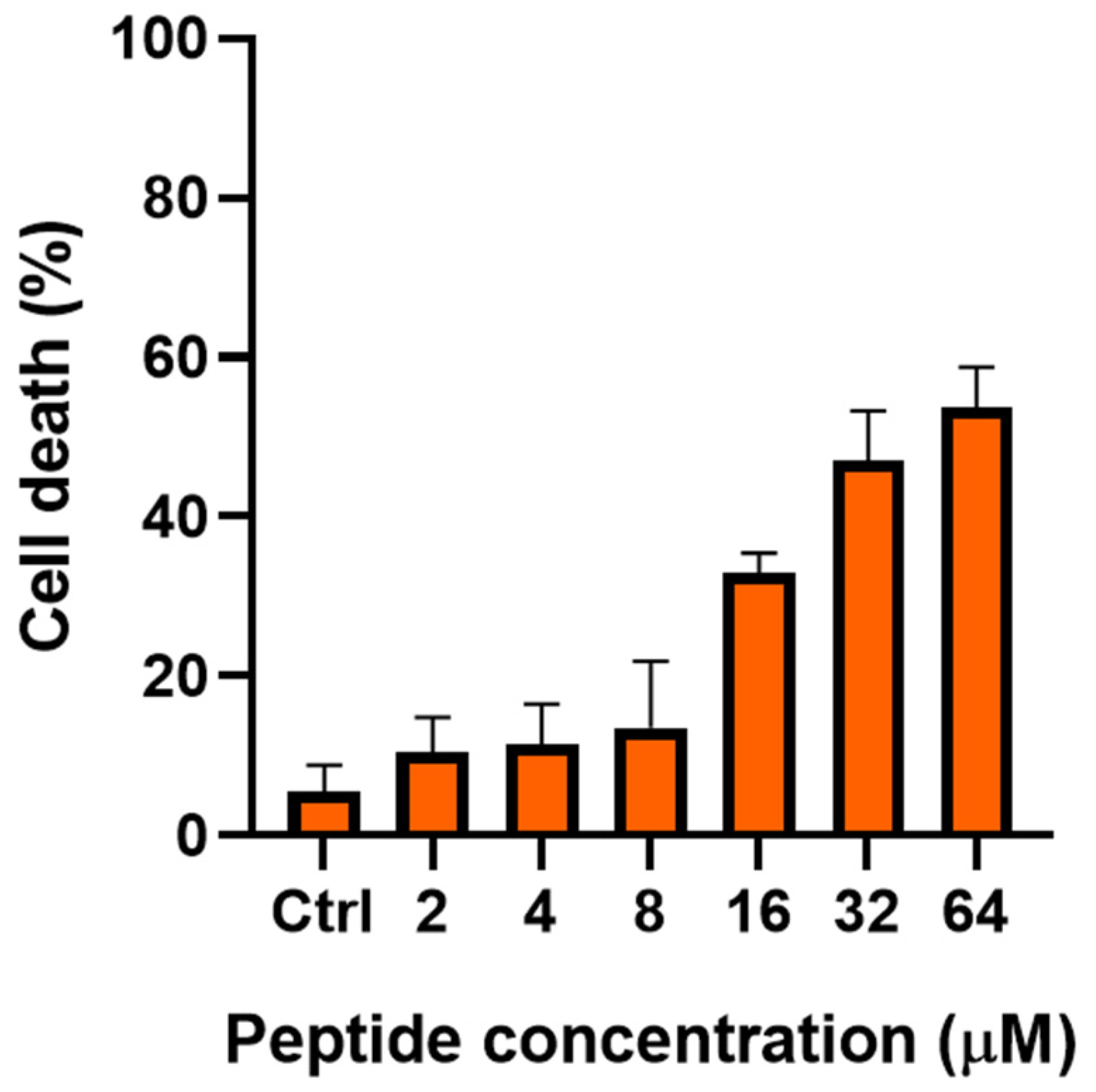
2.2. Fungal Cell Viability Decreases at Increasing Concentrations of TG
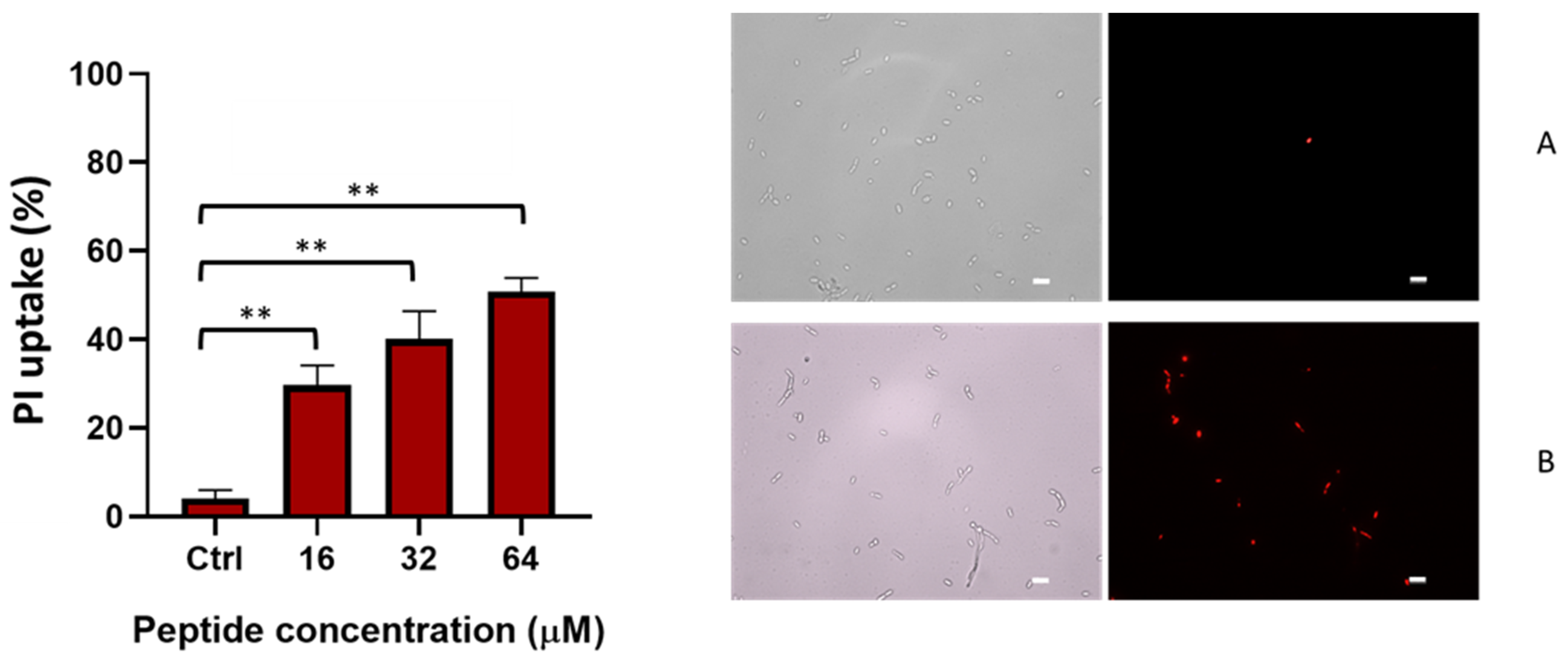
2.3. TG Induces Membrane Perturbation
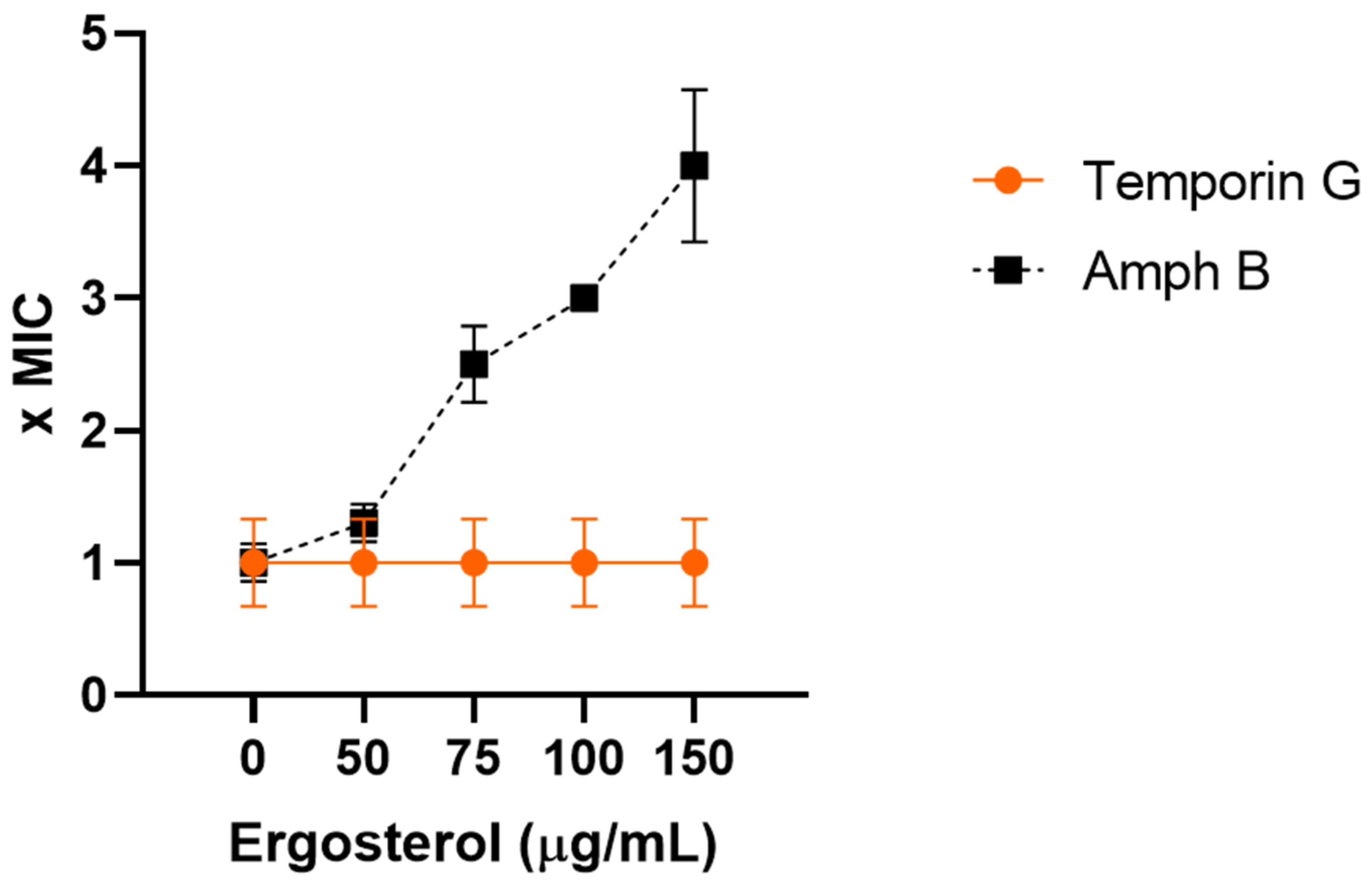
2.4. TG Does Not Directly Bind Ergosterol
2.5. TG Does Not Affect Fungal Cell Wall Synthesis
2.6. TG Is Active against Some Candida Virulence Factors
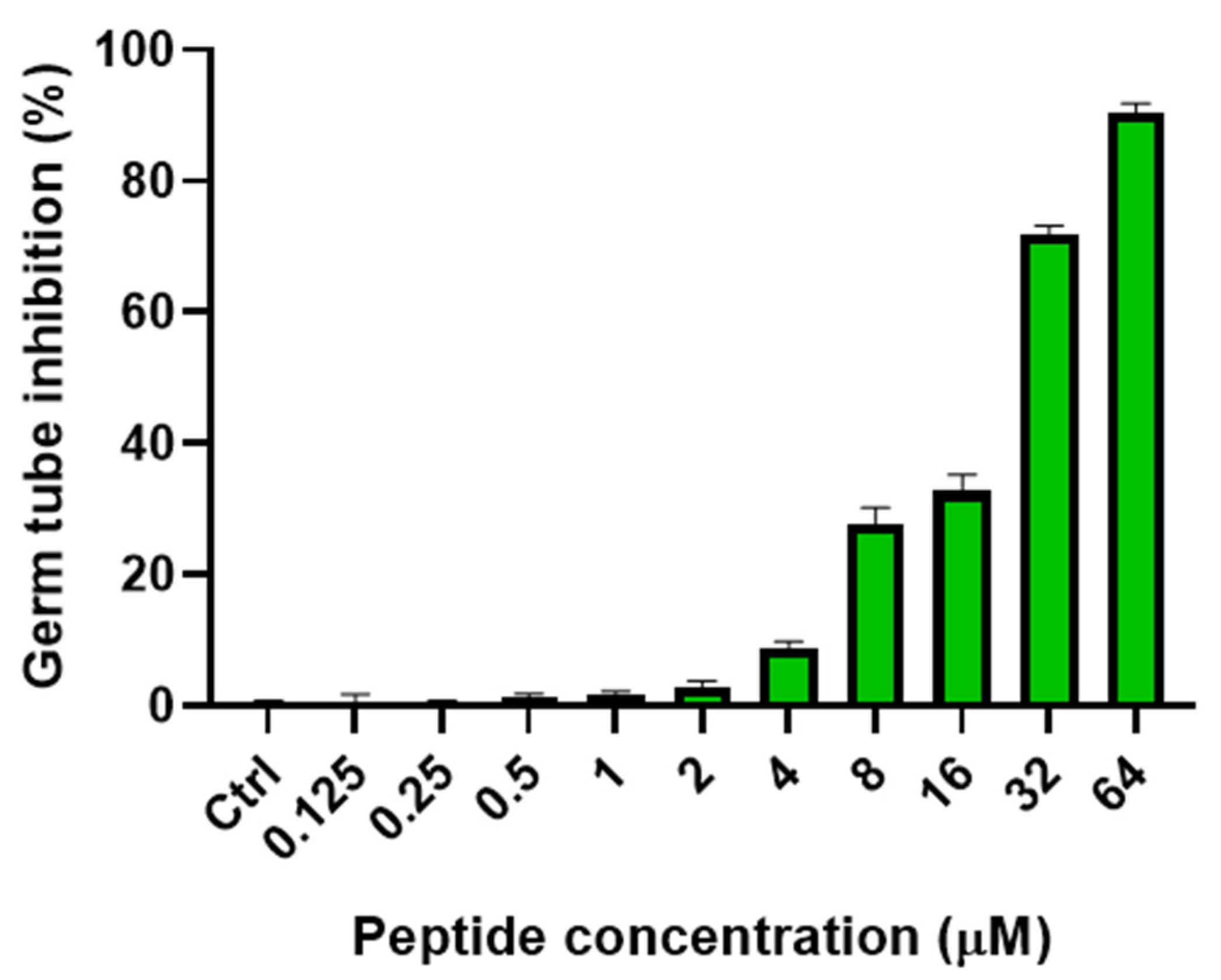
2.6.1. Inhibition of Yeast-Mycelial Conversion
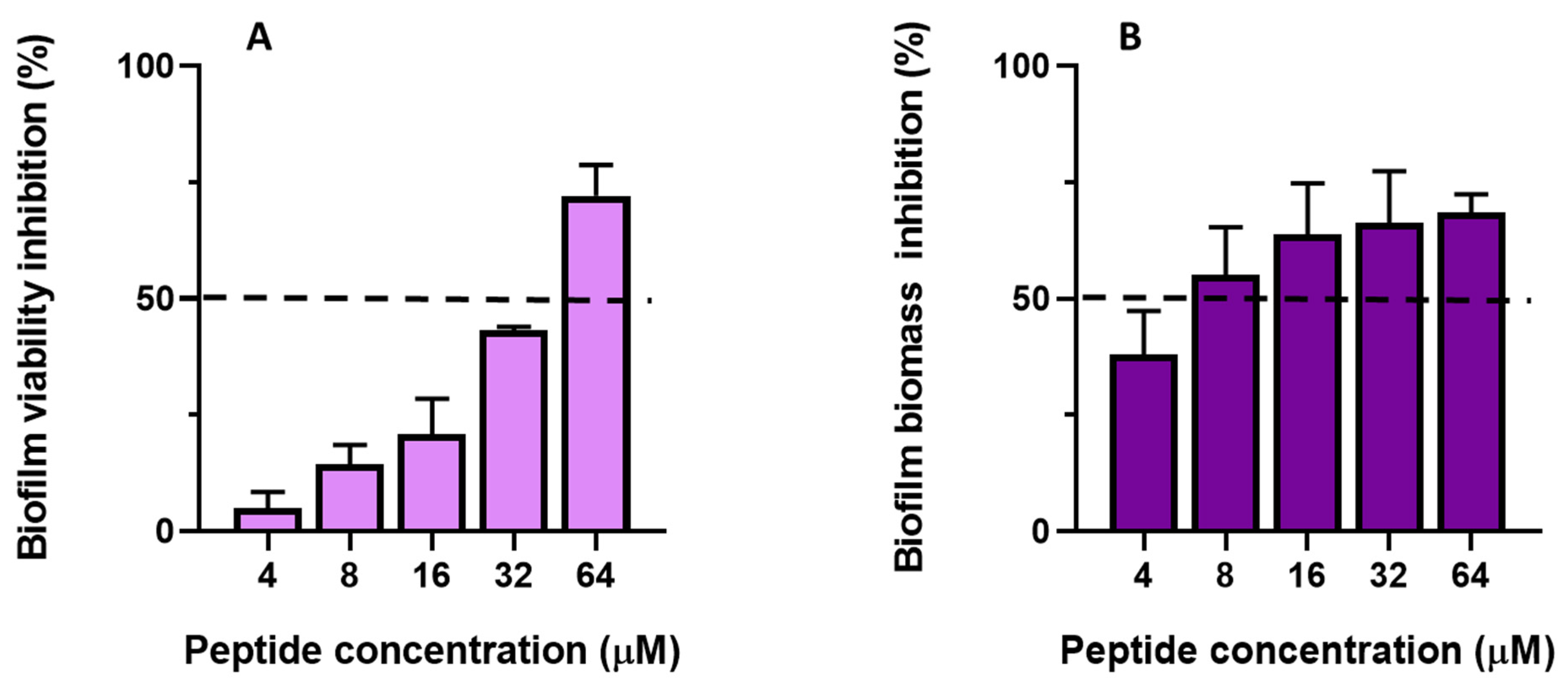
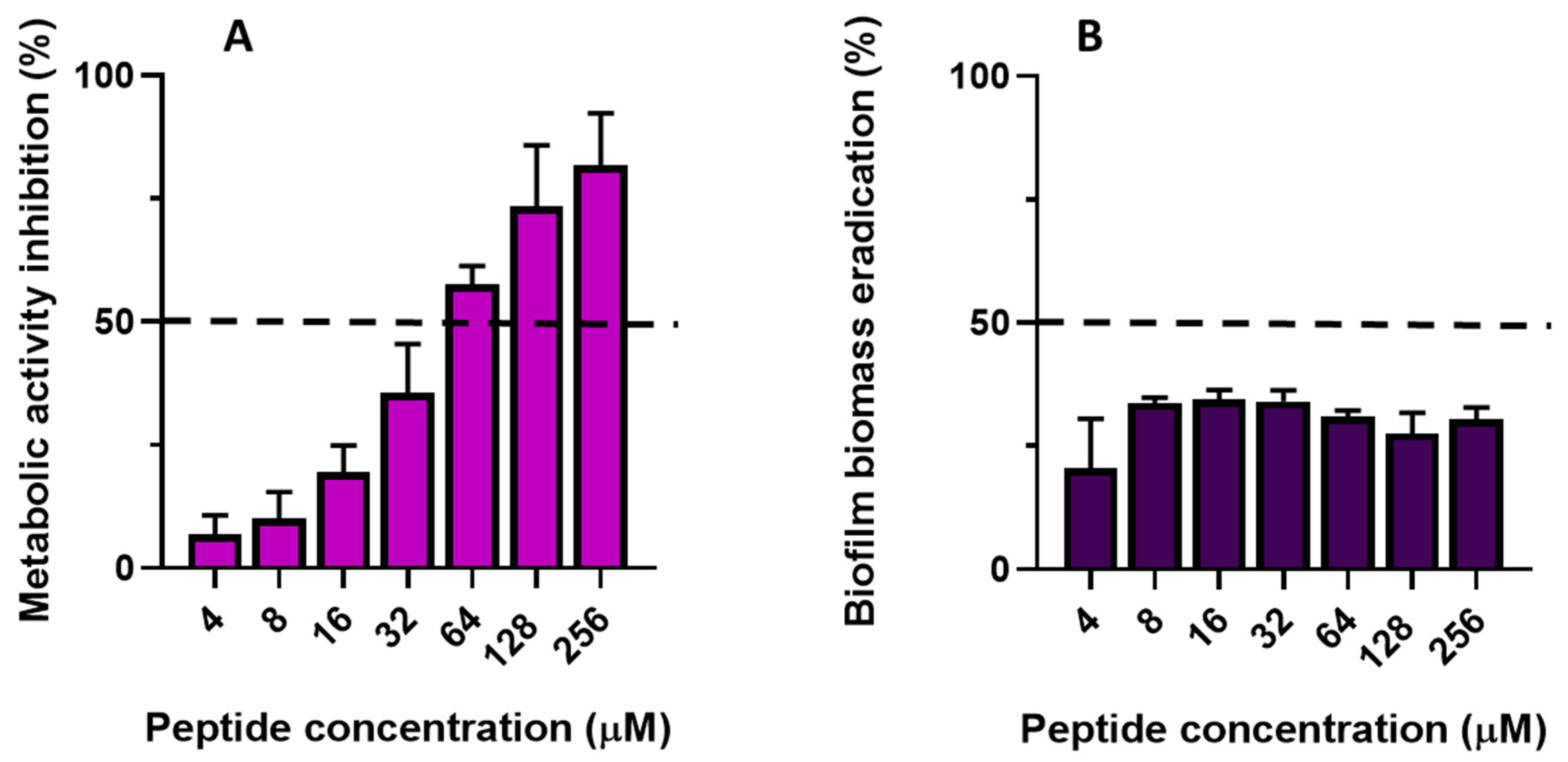
2.6.2. Antibiofilm Activity of TG
3. Discussion
4. Materials and Methods
4.1. Fungal Strains
4.2. TG and Antifungal Drugs
4.3. Antifungal Susceptibility Testing
4.4. Cells Viability Assay
4.5. Membrane Permeabilization
4.5.1. Sytox Green Assay
4.5.2. PI Uptake Assay
4.6. Ergosterol Binding Assay
4.7. Sorbitol Effect Assay
4.8. Effect of TG on C. albicans Virulence Factors
4.8.1. Yeast–Mycelial Conversion Assay
4.8.2. Anti-Biofilm Assay
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and Multi-National Prevalence of Fungal Diseases-Estimate Precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y. Human fungal pathogens: Why should we learn? J. Microbiol. 2016, 54, 145–148. [Google Scholar] [CrossRef] [Green Version]
- Denning, D.W.; Kneale, M.; Sobel, J.D.; Rautemaa-Richardson, R. Global burden of recurrent vulvovaginal candidiasis: A systematic review. Lancet. Infect. Dis. 2018, 18, e339–e347. [Google Scholar] [CrossRef]
- Firacative, C. Invasive fungal disease in humans: Are we aware of the real impact? Mem. Inst. Oswaldo Cruz 2020, 115, e200430. [Google Scholar] [CrossRef]
- Schmiedel, Y.; Zimmerli, S. Common invasive fungal diseases: An overview of invasive candidiasis, aspergillosis, cryptococcosis, and Pneumocystis pneumonia. Swiss Med. Wkly. 2016, 146, w14281. [Google Scholar] [CrossRef] [Green Version]
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 2018, 360, 739–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, S.; Rodrigues, C.F.; Araujo, D.; Rodrigues, M.E.; Henriques, M. Candida Species Biofilms’ Antifungal Resistance. J. Fungi 2017, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, D.M.; Ghannoum, M.A. Candida biofilms: Antifungal resistance and emerging therapeutic options. Curr. Opin. Investig. Drugs 2004, 5, 186–197. [Google Scholar]
- Boman, H.G. Innate immunity and the normal microflora. Immunol. Rev. 2000, 173, 5–16. [Google Scholar] [CrossRef]
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmaceuticals 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [Green Version]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Stocklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef]
- Vanzolini, T.; Bruschi, M.; Rinaldi, A.C.; Magnani, M.; Fraternale, A. Multitalented Synthetic Antimicrobial Peptides and Their Antibacterial, Antifungal and Antiviral Mechanisms. Int. J. Mol. Sci. 2022, 23, 545. [Google Scholar] [CrossRef]
- Chianese, A.; Zannella, C.; Monti, A.; De Filippis, A.; Doti, N.; Franci, G.; Galdiero, M. The Broad-Spectrum Antiviral Potential of the Amphibian Peptide AR-23. Int. J. Mol. Sci. 2022, 23, 883. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Feix, J.B. Peptide-membrane interactions and mechanisms of membrane destruction by amphipathic alpha-helical antimicrobial peptides. Biochim. Biophys. Acta 2006, 1758, 1245–1256. [Google Scholar] [CrossRef] [Green Version]
- Romero, S.M.; Cardillo, A.B.; Martinez Ceron, M.C.; Camperi, S.A.; Giudicessi, S.L. Temporins: An Approach of Potential Pharmaceutic Candidates. Surg. Infect. 2020, 21, 309–322. [Google Scholar] [CrossRef]
- Rinaldi, A.C.; Mangoni, M.L.; Rufo, A.; Luzi, C.; Barra, D.; Zhao, H.; Kinnunen, P.K.; Bozzi, A.; Di Giulio, A.; Simmaco, M. Temporin L: Antimicrobial, haemolytic and cytotoxic activities, and effects on membrane permeabilization in lipid vesicles. Biochem. J. 2002, 368, 91–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raja, Z.; Andre, S.; Abbassi, F.; Humblot, V.; Lequin, O.; Bouceba, T.; Correia, I.; Casale, S.; Foulon, T.; Sereno, D.; et al. Insight into the mechanism of action of temporin-SHa, a new broad-spectrum antiparasitic and antibacterial agent. PLoS ONE 2017, 12, e0174024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wade, D.; Silberring, J.; Soliymani, R.; Heikkinen, S.; Kilpelainen, I.; Lankinen, H.; Kuusela, P. Antibacterial activities of temporin A analogs. FEBS Lett. 2000, 479, 6–9. [Google Scholar] [CrossRef]
- Marcocci, M.E.; Amatore, D.; Villa, S.; Casciaro, B.; Aimola, P.; Franci, G.; Grieco, P.; Galdiero, M.; Palamara, A.T.; Mangoni, M.L.; et al. The Amphibian Antimicrobial Peptide Temporin B Inhibits In Vitro Herpes Simplex Virus 1 Infection. Antimicrob. Agents Chemother. 2018, 62, 367. [Google Scholar] [CrossRef] [Green Version]
- Simmaco, M.; Mignogna, G.; Canofeni, S.; Miele, R.; Mangoni, M.L.; Barra, D. Temporins, antimicrobial peptides from the European red frog Rana temporaria. Eur. J. Biochem. 1996, 242, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, A.C.; Conlon, J.M. Chapter 56–Temporins. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 400–406. [Google Scholar] [CrossRef]
- Capparelli, R.; Romanelli, A.; Iannaccone, M.; Nocerino, N.; Ripa, R.; Pensato, S.; Pedone, C.; Iannelli, D. Synergistic antibacterial and anti-inflammatory activity of temporin A and modified temporin B in vivo. PLoS ONE 2009, 4, e7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roscetto, E.; Bellavita, R.; Paolillo, R.; Merlino, F.; Molfetta, N.; Grieco, P.; Buommino, E.; Catania, M.R. Antimicrobial Activity of a Lipidated Temporin L Analogue against Carbapenemase-Producing Klebsiella pneumoniae Clinical Isolates. Antibiotics 2021, 10, 1312. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Grazia, A.D.; Cappiello, F.; Casciaro, B.; Luca, V. Naturally Occurring Peptides from Rana temporaria: Antimicrobial Properties and More. Curr. Top. Med. Chem. 2016, 16, 54–64. [Google Scholar] [CrossRef]
- De Angelis, M.; Casciaro, B.; Genovese, A.; Brancaccio, D.; Marcocci, M.E.; Novellino, E.; Carotenuto, A.; Palamara, A.T.; Mangoni, M.L.; Nencioni, L. Temporin G, an amphibian antimicrobial peptide against influenza and parainfluenza respiratory viruses: Insights into biological activity and mechanism of action. FASEB J. 2021, 35, e21358. [Google Scholar] [CrossRef]
- Casciaro, B.; Loffredo, M.R.; Cappiello, F.; Fabiano, G.; Torrini, L.; Mangoni, M.L. The Antimicrobial Peptide Temporin G: Anti-Biofilm, Anti-Persister Activities, and Potentiator Effect of Tobramycin Efficacy Against Staphylococcus aureus. Int. J. Mol. Sci. 2020, 21, 9410. [Google Scholar] [CrossRef]
- Tyc, K.M.; Kuhn, C.; Wilson, D.; Klipp, E. Assessing the advantage of morphological changes in Candida albicans: A game theoretical study. Front. Microbiol. 2014, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida albicans biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, R.; Dos Santos Fontenelle, R.O.; de Brito, E.H.S.; de Morais, S.M. Biofilm of Candida albicans: Formation, regulation and resistance. J. Appl. Microbiol. 2021, 131, 11–22. [Google Scholar] [CrossRef]
- Rajendran, R.; Sherry, L.; Nile, C.J.; Sherriff, A.; Johnson, E.M.; Hanson, M.F.; Williams, C.; Munro, C.A.; Jones, B.J.; Ramage, G. Biofilm formation is a risk factor for mortality in patients with Candida albicans bloodstream infection-Scotland, 2012–2013. Clin. Microbiol. Infect. 2016, 22, 87–93. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 4th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Luca, V.; Olivi, M.; Di Grazia, A.; Palleschi, C.; Uccelletti, D.; Mangoni, M.L. Anti-Candida activity of 1–18 fragment of the frog skin peptide esculentin-1b: In vitro and in vivo studies in a Caenorhabditis elegans infection model. Cell. Mol. Life Sci. CMLS 2014, 71, 2535–2546. [Google Scholar] [CrossRef] [PubMed]
- Casciaro, B.; Loffredo, M.R.; Cappiello, F.; Verrusio, W.; Corleto, V.D.; Mangoni, M.L. Frog Skin-Derived Peptides Against Corynebacterium jeikeium: Correlation between Antibacterial and Cytotoxic Activities. Antibiotics 2020, 9, 448. [Google Scholar] [CrossRef]
- Odds, F.C.; Brown, A.J.; Gow, N.A. Antifungal agents: Mechanisms of action. Trends Microbiol. 2003, 11, 272–279. [Google Scholar] [CrossRef]
- Escalante, A.; Gattuso, M.; Perez, P.; Zacchino, S. Evidence for the mechanism of action of the antifungal phytolaccoside B isolated from Phytolacca tetramera Hauman. J. Nat. Prod. 2008, 71, 1720–1725. [Google Scholar] [CrossRef] [PubMed]
- Biernasiuk, A.; Berecka-Rycerz, A.; Gumieniczek, A.; Malm, M.; Laczkowski, K.Z.; Szymanska, J.; Malm, A. The newly synthesized thiazole derivatives as potential antifungal compounds against Candida albicans. Appl. Microbiol. Biotechnol. 2021, 105, 6355–6367. [Google Scholar] [CrossRef]
- Sentandreu, R.; Herrero, E.; Elorza, M.V.; Rico, H.; Pastor, J. Synthesis and assembly of wall polymers on regenerating yeast protoplasts. Exp. Suppl. 1983, 46, 187–195. [Google Scholar] [CrossRef]
- de Oliveira Filho, G.B.; Cardoso, M.V.O.; Espindola, J.W.P.; Oliveira, E.S.D.A.; Ferreira, R.S.; Coelho, P.L.; Anjos, P.S.D.; Santos, E.S.; Meira, C.S.; Moreira, D.R.M.; et al. Structural design, synthesis and pharmacological evaluation of thiazoles against Trypanosoma cruzi. Eur. J. Med. Chem. 2017, 141, 346–361. [Google Scholar] [CrossRef]
- Chen, H.; Zhou, X.; Ren, B.; Cheng, L. The regulation of hyphae growth in Candida albicans. Virulence 2020, 11, 337–348. [Google Scholar] [CrossRef] [Green Version]
- Mukaremera, L.; Lee, K.K.; Mora-Montes, H.M.; Gow, N.A.R. Candida albicans Yeast, Pseudohyphal, and Hyphal Morphogenesis Differentially Affects Immune Recognition. Front. Immunol. 2017, 8, 629. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, I.D.; Wilson, D.; Wachtler, B.; Brunke, S.; Naglik, J.R.; Hube, B. Candida albicans dimorphism as a therapeutic target. Expert Rev. Anti-Infect. Ther. 2012, 10, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Lo, H.J.; Kohler, J.R.; DiDomenico, B.; Loebenberg, D.; Cacciapuoti, A.; Fink, G.R. Nonfilamentous C. albicans mutants are avirulent. Cell 1997, 90, 939–949. [Google Scholar] [CrossRef] [Green Version]
- Richardson, M.D.W. Fungal Infection: Diagnosis and Management, 4th ed.; Wiley-Blackwell: Oxford, UK, 2012. [Google Scholar]
- Hallen-Adams, H.E.; Suhr, M.J. Fungi in the healthy human gastrointestinal tract. Virulence 2017, 8, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Colombo, A.L.; Guimaraes, T.; Sukienik, T.; Pasqualotto, A.C.; Andreotti, R.; Queiroz-Telles, F.; Nouer, S.A.; Nucci, M. Prognostic factors and historical trends in the epidemiology of candidemia in critically ill patients: An analysis of five multicenter studies sequentially conducted over a 9-year period. Intensive Care Med. 2014, 40, 1489–1498. [Google Scholar] [CrossRef] [Green Version]
- Moran, G.; Sullivan, D. An introduction to the medically important Candida species. In Candida and Candidiasis; Calderone, R.A., Ed.; ASM Press: Washington, DC, USA, 2019; pp. 11–27. [Google Scholar]
- Arastehfar, A.; Carvalho, A.; Nguyen, M.H.; Hedayati, M.T.; Netea, M.G.; Perlin, D.S.; Hoenigl, M. COVID-19-Associated Candidiasis (CAC): An Underestimated Complication in the Absence of Immunological Predispositions? J. Fungi 2020, 6, 211. [Google Scholar] [CrossRef]
- Rolling, T.; Hohl, T.M.; Zhai, B. Minority report: The intestinal mycobiota in systemic infections. Curr. Opin. Microbiol. 2020, 56, 1–6. [Google Scholar] [CrossRef]
- Wadhwa, R.; Pandey, P.; Gupta, G.; Aggarwal, T.; Kumar, N.; Mehta, M.; Satija, S.; Gulati, M.; Madan, J.; Dureja, H.; et al. Emerging Complexity and the Need for Advanced Drug Delivery in Targeting Candida Species. Curr. Top. Med. Chem. 2019, 19, 2593–2609. [Google Scholar] [CrossRef] [PubMed]
- Wall, G.; Montelongo-Jauregui, D.; Vidal Bonifacio, B.; Lopez-Ribot, J.L.; Uppuluri, P. Candida albicans biofilm growth and dispersal: Contributions to pathogenesis. Curr. Opin. Microbiol. 2019, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Kernien, J.F.; Snarr, B.D.; Sheppard, D.C.; Nett, J.E. The Interface between Fungal Biofilms and Innate Immunity. Front. Immunol. 2017, 8, 1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalheiro, M.; Teixeira, M.C. Candida Biofilms: Threats, Challenges, and Promising Strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Candida biofilms associated with CVC and medical devices. Mycoses 2012, 55, 46–57. [Google Scholar] [CrossRef]
- Tumbarello, M.; Fiori, B.; Trecarichi, E.M.; Posteraro, P.; Losito, A.R.; De Luca, A.; Sanguinetti, M.; Fadda, G.; Cauda, R.; Posteraro, B. Risk factors and outcomes of candidemia caused by biofilm-forming isolates in a tertiary care hospital. PLoS ONE 2012, 7, e33705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitalis, E.; Nagy, F.; Toth, Z.; Forgacs, L.; Bozo, A.; Kardos, G.; Majoros, L.; Kovacs, R. Candida biofilm production is associated with higher mortality in patients with candidaemia. Mycoses 2020, 63, 352–360. [Google Scholar] [CrossRef]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Kowalski, C.H.; Morelli, K.A.; Schultz, D.; Nadell, C.D.; Cramer, R.A. Fungal biofilm architecture produces hypoxic microenvironments that drive antifungal resistance. Proc. Natl. Acad. Sci. USA 2020, 117, 22473–22483. [Google Scholar] [CrossRef]
- Costa-de-Oliveira, S.; Rodrigues, A.G. Candida albicans Antifungal Resistance and Tolerance in Bloodstream Infections: The Triad Yeast-Host-Antifungal. Microorganisms 2020, 8, 154. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rubio, R.; Cuenca-Estrella, M.; Mellado, E. Triazole Resistance in Aspergillus Species: An Emerging Problem. Drugs 2017, 77, 599–613. [Google Scholar] [CrossRef]
- Alexander, B.D.; Johnson, M.D.; Pfeiffer, C.D.; Jimenez-Ortigosa, C.; Catania, J.; Booker, R.; Castanheira, M.; Messer, S.A.; Perlin, D.S.; Pfaller, M.A. Increasing Echinocandin Resistance in Candida glabrata: Clinical Failure Correlates with Presence of FKS Mutations and Elevated Minimum Inhibitory Concentrations. Clin. Infect. Dis. 2013, 56, 1724–1732. [Google Scholar] [CrossRef] [Green Version]
- Peyclit, L.; Yousfi, H.; Rolain, J.M.; Bittar, F. Drug Repurposing in Medical Mycology: Identification of Compounds as Potential Antifungals to Overcome the Emergence of Multidrug-Resistant Fungi. Pharm. Base 2021, 14, 488. [Google Scholar] [CrossRef] [PubMed]
- Taylor, G. The importance of stewardship. Can. Commun. Dis. Rep. 2015, 41, 3–4. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Antimicrobial Resistance Global Report on Surveillance: 2014 Summary; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- Geddes-McAlister, J.; Shapiro, R.S. New pathogens, new tricks: Emerging, drug-resistant fungal pathogens and future prospects for antifungal therapeutics. Annu. N. Y. Acad. Sci. 2019, 1435, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L.; Miele, R.; Renda, T.G.; Barra, D.; Simmaco, M. The synthesis of antimicrobial peptides in the skin of Rana esculenta is stimulated by microorganisms. FASEB J. 2001, 15, 1431–1432. [Google Scholar] [CrossRef]
- Grieco, P.; Carotenuto, A.; Auriemma, L.; Saviello, M.R.; Campiglia, P.; Gomez-Monterrey, I.M.; Marcellini, L.; Luca, V.; Barra, D.; Novellino, E.; et al. The effect of D-amino acid substitution on the selectivity of temporin L towards target cells: Identification of a potent anti-Candida peptide. Biochim. Biophys. Acta (BBA)-Biomembr. 2013, 1828, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Bellavita, R.; Maione, A.; Merlino, F.; Siciliano, A.; Dardano, P.; De Stefano, L.; Galdiero, S.; Galdiero, E.; Grieco, P.; Falanga, A. Antifungal and Antibiofilm Activity of Cyclic Temporin L Peptide Analogues against Albicans and Non-Albicans Candida Species. Pharmaceutics 2022, 14, 454. [Google Scholar] [CrossRef] [PubMed]
- Brunet, K.; Verdon, J.; Ladram, A.; Arnault, S.; Rodier, M.H.; Cateau, E. Antifungal activity of [K3] temporin-SHa against medically relevant yeasts and moulds. Can. J. Microbiol. 2022, 68, 427–434. [Google Scholar] [CrossRef]
- Iyer, K.R.; Revie, N.M.; Fu, C.; Robbins, N.; Cowen, L.E. Treatment strategies for cryptococcal infection: Challenges, advances and future outlook. Nat. Rev. Microbiol. 2021, 19, 454–466. [Google Scholar] [CrossRef]
- Sahoo, A.K.; Mahajan, R. Management of tinea corporis, tinea cruris, and tinea pedis: A comprehensive review. Indian Dermatol. Online J. 2016, 7, 77–86. [Google Scholar] [CrossRef]
- Khurana, A.; Sardana, K.; Chowdhary, A. Antifungal resistance in dermatophytes: Recent trends and therapeutic implications. Fungal Genet. Biol. 2019, 132, 103255. [Google Scholar] [CrossRef]
- Reichert-Lima, F.; Lyra, L.; Pontes, L.; Moretti, M.L.; Pham, C.D.; Lockhart, S.R.; Schreiber, A.Z. Surveillance for azoles resistance in Aspergillus spp. highlights a high number of amphotericin B-resistant isolates. Mycoses 2018, 61, 360–365. [Google Scholar] [CrossRef]
- Lima, P.G.; Souza, P.F.N.; Freitas, C.D.T.; Bezerra, L.P.; Neto, N.A.S.; Silva, A.F.B.; Oliveira, J.T.A.; Sousa, D.O.B. Synthetic peptides against Trichophyton mentagrophytes and T. rubrum: Mechanisms of action and efficiency compared to griseofulvin and itraconazole. Life Sci. 2021, 265, 118803. [Google Scholar] [CrossRef]
- Cavalcante, C.S.; Falcao, C.B.; Fontenelle, R.O.; Andreu, D.; Radis-Baptista, G. Anti-fungal activity of Ctn [15–34], the C-terminal peptide fragment of crotalicidin, a rattlesnake venom gland cathelicidin. J. Antibiot. 2017, 70, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.J.; Xie, T.; Wu, L.; Liu, X.Y.; Zhu, L.; Chen, Y.; Mao, E.Q.; Han, L.Z.; Chen, E.Z.; Yang, Z.T. Epidemiology, species distribution, and outcome of nosocomial Candida spp. bloodstream infection in Shanghai: An 11-year retrospective analysis in a tertiary care hospital. Annu. Clin. Microbiol. Antimicrob. 2021, 20, 34. [Google Scholar] [CrossRef]
- Mahalka, A.K.; Kinnunen, P.K. Binding of amphipathic alpha-helical antimicrobial peptides to lipid membranes: Lessons from temporins B and L. Biochim. Biophys. Acta 2009, 1788, 1600–1609. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.; Van Dijck, P.; Datta, A. Environmental sensing and signal transduction pathways regulating morphopathogenic determinants of Candida albicans. Microbiol. Mol. Biol. Rev. 2007, 71, 348–376. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.P. Dimorphism and virulence in Candida albicans. Curr. Opin. Microbiol. 1998, 1, 687–692. [Google Scholar] [CrossRef]
- Saville, S.P.; Lazzell, A.L.; Monteagudo, C.; Lopez-Ribot, J.L. Engineered control of cell morphology in vivo reveals distinct roles for yeast and filamentous forms of Candida albicans during infection. Eukaryot. Cell. 2003, 2, 1053–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlisle, P.L.; Banerjee, M.; Lazzell, A.; Monteagudo, C.; Lopez-Ribot, J.L.; Kadosh, D. Expression levels of a filament-specific transcriptional regulator are sufficient to determine Candida albicans morphology and virulence. Proc. Natl. Acad. Sci. USA 2009, 106, 599–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachtler, B.; Wilson, D.; Haedicke, K.; Dalle, F.; Hube, B. From Attachment to Damage: Defined Genes of Candida albicans Mediate Adhesion, Invasion and Damage during Interaction with Oral Epithelial Cells. PLoS ONE 2011, 6, 7046. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, M.C.; Bender, J.A.; Fink, G.R. Transcriptional response of Candida albicans upon internalization by macrophages. Eukaryot. Cell 2004, 3, 1076–1087. [Google Scholar] [CrossRef] [Green Version]
- Richard, M.L.; Nobile, C.J.; Bruno, V.M.; Mitchell, A.P. Candida albicans biofilm-defective mutants. Eukaryot. Cell 2005, 4, 1493–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramage, G.; VandeWalle, K.; Lopez-Ribot, J.L.; Wickes, B.L. The filamentation pathway controlled by the Efg1 regulator protein is required for normal biofilm formation and development in Candida albicans. Fems. Microbiol. Lett. 2002, 214, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm formation by the fungal pathogen Candida albicans: Development, architecture, and drug resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef] [Green Version]
- Deveau, A.; Hogan, D.A. Linking Quorum Sensing Regulation and Biofilm Formation by Candida albicans. Quor. Sens. Methods Protoc. 2011, 692, 219–233. [Google Scholar] [CrossRef]
- Saville, S.P.; Lazzell, A.L.; Bryant, A.P.; Fretzen, A.; Monreal, A.; Solberg, E.O.; Monteagudo, C.; Lopez-Ribot, J.L.; Milne, G.T. Inhibition of filamentation can be used to treat disseminated candidiasis. Antimicrob. Agents Chemother. 2006, 50, 3312–3316. [Google Scholar] [CrossRef] [Green Version]
- Vila, T.; Romo, J.A.; Pierce, C.G.; McHardy, S.F.; Saville, S.P.; Lopez-Ribot, J.L. Targeting Candida albicans filamentation for antifungal drug development. Virulence 2017, 8, 150–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, P.F.N.; Marques, L.S.M.; Oliveira, J.T.A.; Lima, P.G.; Dias, L.P.; Neto, N.A.S.; Lopes, F.E.S.; Sousa, J.S.; Silva, A.F.B.; Caneiro, R.F.; et al. Synthetic antimicrobial peptides: From choice of the best sequences to action mechanisms. Biochimie 2020, 175, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Painting, K.; Kirsop, B. A Quick Method for Estimating the Percentage of Viable Cells in a Yeast Population, Using Methylene-Blue Staining. World J. Microb. Biot. 1990, 6, 346–347. [Google Scholar] [CrossRef]
- Van Zandyke, S.M.; Simal, O.; Gualdoni, S.; Smart, K.A. Yeast quality and fluorophore technologies. In Brewing Yeast Fermentation Performance, 2nd ed.; Smart, K.A., Ed.; Blackwell Science: Oxford, UK, 2003; Volume 14, pp. 149–160. [Google Scholar]
- Merlino, F.; Carotenuto, A.; Casciaro, B.; Martora, F.; Loffredo, M.R.; Di Grazia, A.; Yousif, A.M.; Brancaccio, D.; Palomba, L.; Novellino, E.; et al. Glycine-replaced derivatives of [Pro(3),DLeu(9)]TL, a temporin L analogue: Evaluation of antimicrobial, cytotoxic and hemolytic activities. Eur. J. Med. Chem. 2017, 139, 750–761. [Google Scholar] [CrossRef]
- D’Auria, F.D.; Tecca, M.; Scazzocchio, F.; Renzini, V.; Strippoli, V. Effect of propolis on virulence factors of Candida albicans. J. Chemother. 2003, 15, 454–460. [Google Scholar] [CrossRef]
- Li, R.F.; Yan, X.H.; Lu, Y.B.; Lu, Y.L.; Zhang, H.R.; Chen, S.H.; Liu, S.; Lu, Z.F. Anti-candidal activity of a novel peptide derived from human chromogranin A and its mechanism of action against Candida krusei. Exp. Ther. Med. 2015, 10, 1768–1776. [Google Scholar] [CrossRef] [Green Version]
- Frost, D.J.; Brandt, K.D.; Cugier, D.; Goldman, R. A whole-cell Candida albicans assay for the detection of inhibitors towards fungal cell wall synthesis and assembly. J. Antibiot. 1995, 48, 306–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassone, A.; Sullivan, P.A.; Shepherd, M.G. N-acetyl-D-glucosamine-induced morphogenesis in Candida albicans. Microbiologica 1985, 8, 85–99. [Google Scholar]
- Simonetti, N.; Strippoli, V.; Cassone, A. Yeast-mycelial conversion induced by N-acetyl-D-glucosamine in Candida albicans. Nature 1974, 250, 344–346. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, F.D.; Laino, L.; Strippoli, V.; Tecca, M.; Salvatore, G.; Battinelli, L.; Mazzanti, G. In vitro activity of tea tree oil against Candida albicans mycelial conversion and other pathogenic fungi. J. Chemother. 2001, 13, 377–383. [Google Scholar] [CrossRef]
- Pierce, C.G.; Uppuluri, P.; Tristan, A.R.; Wormley, F.L.; Mowat, E.; Ramage, G.; Lopez-Ribot, J.L. A simple and reproducible 96-well plate-based method for the formation of fungal biofilms and its application to antifungal susceptibility testing. Nat. Protoc. 2008, 3, 1494–1500. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H.; Shang, Z.C.; Li, T.X.; Yang, M.H.; Kong, L.Y. In Vitro Antibiofilm Activity of Eucarobustol E against Candida albicans. Antimicrob. Agents Chemother. 2017, 61, 707. [Google Scholar] [CrossRef] [Green Version]
- Melo, A.S.; Bizerra, F.C.; Freymuller, E.; Arthington-Skaggs, B.A.; Colombo, A.L. Biofilm production and evaluation of antifungal susceptibility amongst clinical Candida spp. isolates, including strains of the Candida parapsilosis complex. Med. Mycol. 2011, 49, 253–262. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temporin G | Amphotericin B | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| MIC50 | MIC100 | MIC100 | ||||||||
| 24 h | 48 h | 24 h | 48 h | 48 h | ||||||
| Mode | Range | Mode | Range | Mode | Range | Mode | Range | Mode | Range | |
| Candida albicans ATCC10231 | 4 | 4–8 | 8 | 4–8 | 16 | 8–32 | 16 | 8–32 | 0.25 | 0.125–0.25 |
| Candida albicans ATCC24433 | 16 | 16–32 | 32 | 32–64 | 32 | 32–64 | 32 | 32–64 | 0.25 | 0.125–0.25 |
| Candida tropicalis DSM11953 | 4 | 4–8 | 8 | 8–16 | 16 | 8–16 | 16 | 16–32 | 1 | 0.5–1 |
| Candida glabrata PMC0805 | 16 | 16–32 | 32 | 32–64 | 64 | 64–128 | 128 | 128 | 4 | 4–8 |
| Candida parapsilosis PMC0711 | 64 | 32–64 | 128 | 128 | 128 | 64–128 | 128 | 128 | 1 | 0.5–2 |
| Candida parapsilosis DSM11224 | 64 | 32–64 | 128 | 128 | 128 | 64–128 | 128 | 128 | 1 | 0.5–1 |
| Candida krusei PMC0610 | 64 | 64 | 128 | 128 | 128 | 64–128 | 128 | 128 | 1 | 0.5–1 |
| Candida krusei PMC0624 | 16 | 8–16 | 32 | 32–64 | 32 | 32–64 | 32 | 32–64 | 1 | 1–2 |
| Cryptococcus neoformans DSM11759 | 16 | 16–32 | 16 | 16–32 | 32 | 16–32 | 32 | 16–64 | 0.5 | 0.25–0.5 |
| Candida spp. Mean ± SD | 30 ± 24.5 | 65.9 ± 48.9 | 65.5 ± 45.9 | 80.2 ± 49.6 | 1.3 ± 1.7 | |||||
| Cryptococcus spp. Mean ± SD | 20 ± 7.4 | 22 ± 8.3 | 28 ± 7.4 | 32 ± 14.8 | 0.4 ± 0.1 | |||||
| Temporin G | Amphotericin B | ||
|---|---|---|---|
| MIC80 | MIC100 | MIC100 | |
| Microsporum gypseum DSM3824 | 32 | 64 | 0.25 |
| Microsporum gypseum PMC7330 | 32 | 128 | 0.25 |
| Microsporum canis DSM10708 | 16 | 64 | 0.125 |
| Trichophyton mentagrophytes DSM4870 | 4 | 16 | 0.125 |
| Trichophyton mentagrophytes PMC6530 | 32 | 64 | 0.25 |
| Trichophyton mentagrophytes PMC6529 | 8 | 32 | 0.25 |
| Trichophyton mentagrophytes PMC6528 | 16 | 32 | 0.5 |
| Aspergillus niger PMC7101 | 128 | 128 | 16 |
| Aspergillus brasiliensis DSM1988 | 128 | 128 | 2 |
| Aspergillus terreus PMC7201 | 128 | 128 | 16 |
| Microsporum spp. Mean ± SD | 27.7 ± 13.9 | 101.3 ± 32.2 | 0.6 ± 0.6 |
| Trichophyton spp. Mean ± SD | 19 ± 20.3 | 42 ± 28.7 | 0.3 ± 0.1 |
| Aspergillus spp. Mean ± SD | 112 ± 28.3 | 124.9 ± 49.6 | 12.7 ± 9.1 |
| −SORBITOL | +SORBITOL | |
|---|---|---|
| Temporin G | 21.3 ± 9.2 | 21.3 ± 9.2 |
| Caspofungin | 0.21 ± 0.07 | 26.7 ± 9.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Auria, F.D.; Casciaro, B.; De Angelis, M.; Marcocci, M.E.; Palamara, A.T.; Nencioni, L.; Mangoni, M.L. Antifungal Activity of the Frog Skin Peptide Temporin G and Its Effect on Candida albicans Virulence Factors. Int. J. Mol. Sci. 2022, 23, 6345. https://doi.org/10.3390/ijms23116345
D’Auria FD, Casciaro B, De Angelis M, Marcocci ME, Palamara AT, Nencioni L, Mangoni ML. Antifungal Activity of the Frog Skin Peptide Temporin G and Its Effect on Candida albicans Virulence Factors. International Journal of Molecular Sciences. 2022; 23(11):6345. https://doi.org/10.3390/ijms23116345
Chicago/Turabian StyleD’Auria, Felicia Diodata, Bruno Casciaro, Marta De Angelis, Maria Elena Marcocci, Anna Teresa Palamara, Lucia Nencioni, and Maria Luisa Mangoni. 2022. "Antifungal Activity of the Frog Skin Peptide Temporin G and Its Effect on Candida albicans Virulence Factors" International Journal of Molecular Sciences 23, no. 11: 6345. https://doi.org/10.3390/ijms23116345
APA StyleD’Auria, F. D., Casciaro, B., De Angelis, M., Marcocci, M. E., Palamara, A. T., Nencioni, L., & Mangoni, M. L. (2022). Antifungal Activity of the Frog Skin Peptide Temporin G and Its Effect on Candida albicans Virulence Factors. International Journal of Molecular Sciences, 23(11), 6345. https://doi.org/10.3390/ijms23116345