
The Complex Mechanism of the Salmonella typhi Biofilm Formation That Facilitates Pathogenicity: A Review


 , ,
, ,  , and
, and
Abstract
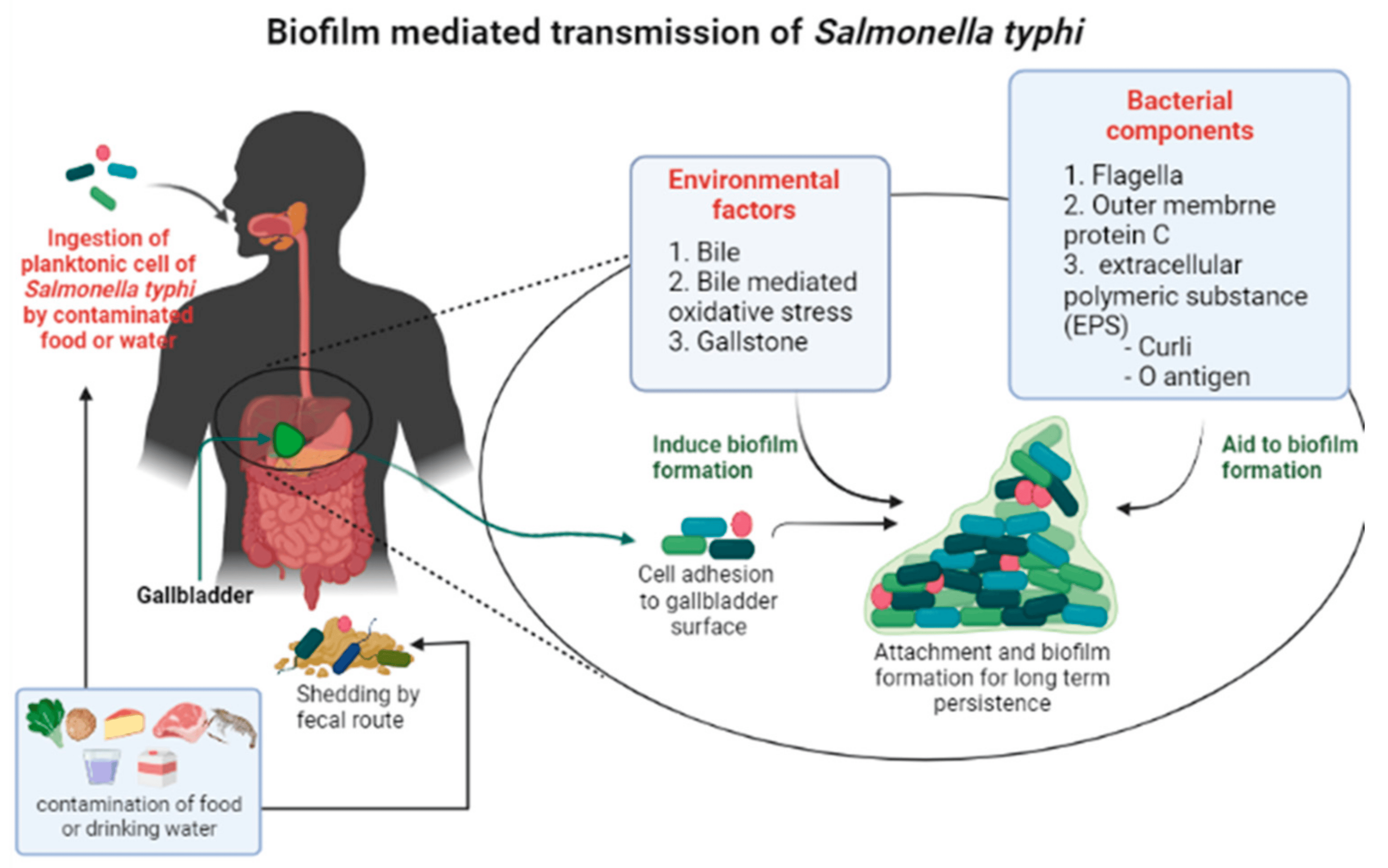
:1. Introduction
2. Environmental Factors Associated with Biofilm Development
2.1. Bile Mediated
2.2. Gallstone Mediated
3. Bacterial Components That Aid in Biofilm Formation
4. Genes and Regulatory Molecules Involved in Biofilm Formation
4.1. Salmonella Pathogenic Islands (SPIs)
4.2. Non-Coding RNAs
4.3. Plasmid Containing Genes Related to Biofilm
4.4. Genes Related to Biofilm
4.5. Role of Quorum Sensing (QS) in Biofilm Formation
4.6. QseB- and QseC-Mediated Biofilm Formation

{kind=link}
| Biofilm-Related Regulatory Molecules in S. typhi | Function | References |
|---|---|---|
| lncRNA AsfD | Increase S. typhi motility by up-regulating different flagellar genes | [51] |
| ncRNA Ribs | Up-regulates the expression of cyclopropane fatty acids synthase gene (cfa) that promotes biofilm | [53] |
| Mig-14 and Virk genes | Decrease the permeability of the outer membrane for PB and encourage the growth of biofilms. | [61] |
| GalE gene | Synthesize galactose which are added to the outer core and the O-antigen | [41] |
| Rck gene | Enhance cellular adhesion of bacteria and promote biofilm formation. | [54] |
| LuxS gene | Encodes autoinducer 2 (AI-2), an essential part of the quorum-sensing mechanism. | [27] |
| pR ST98 plasmid | Contains genes that may be related to biofilm formation. | [55] |
| QseB and QseC | Involved in motility and biofilm formation. | [75] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dougan, G.; Baker, S. Salmonella Enterica Serovar Typhi and the Pathogenesis of Typhoid Fever. Annu. Rev. Microbiol. 2014, 68, 317–336. [Google Scholar] [CrossRef] [PubMed]
- Davies, D. Understanding Biofilm Resistance to Antibacterial Agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Kariuki, S.; Revathi, G.; Kiiru, J.; Mengo, D.M.; Mwituria, J.; Muyodi, J.; Munyalo, A.; Teo, Y.Y.; Holt, K.E.; Kingsley, R.A.; et al. Typhoid in Kenya Is Associated with a Dominant Multidrug-Resistant Salmonella Enterica Serovar Typhi Haplotype That Is Also Widespread in Southeast Asia. J. Clin. Microbiol. 2010, 48, 2171–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flemming, H.; Wuertz, S. Bacteria and Archaea on Earth and Their Abundance in Biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Branda, S.S.; Chu, F.; Kearns, D.B.; Losick, R.; Kolter, R. A Major Protein Component of the Bacillus Subtilis Biofilm Matrix. Mol. Microbiol. 2006, 59, 1229–1238. [Google Scholar] [CrossRef]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial Biofilms: A Common Cause of Persistent Infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Dimakopoulou-Papazoglou, D.; Lianou, A.; Koutsoumanis, K.P. Modelling Biofilm Formation of Salmonella Enterica Ser. Newport as a Function of PH and Water Activity. Food Microbiol. 2016, 53, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.-C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An Emergent Form of Bacterial Life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Stoodley, P. Evolving Concepts in Biofilm Infections. Cell. Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef]
- Steenackers, H.; Hermans, K.; Vanderleyden, J.; De Keersmaecker, S.C.J. Salmonella Biofilms: An Overview on Occurrence, Structure, Regulation and Eradication. Food Res. Int. 2012, 45, 502–531. [Google Scholar] [CrossRef]
- Jensen, P.Ø.; Givskov, M.; Bjarnsholt, T.; Moser, C. The Immune System vs. Pseudomonas Aeruginosa Biofilms. FEMS Immunol. Med. Microbiol. 2010, 59, 292–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, B.M.; Jabra-Rizk, M.A.; O’May, G.A.; Costerton, J.W.; Shirtliff, M.E. Polymicrobial Interactions: Impact on Pathogenesis and Human Disease. Clin. Microbiol. Rev. 2012, 25, 193–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, A.; Mikkelsen, H.; Jungersen, G. Intracellular Pathogens: Host Immunity and Microbial Persistence Strategies. J. Immunol. Res. 2019, 2019, e1356540. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Kuchma, S.L.; O’Toole, G.A. Keeping Their Options Open: Acute versus Persistent Infections. J. Bacteriol. 2006, 188, 1211–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crump, J.A.; Sjölund-Karlsson, M.; Gordon, M.A.; Parry, C.M. Epidemiology, Clinical Presentation, Laboratory Diagnosis, Antimicrobial Resistance, and Antimicrobial Management of Invasive Salmonella Infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [Green Version]
- Gaines, S.; Landy, M.; Edsall, G.; Mandel, A.D.; Trapani, R.-J.; Benenson, A.S. Studies on infection and immunity in experimental typhoid fever. J. Exp. Med. 1961, 114, 327–342. [Google Scholar] [CrossRef] [Green Version]
- Nath, G.; Singh, Y.K.; Kumar, K.; Gulati, A.K.; Shukla, V.K.; Khanna, A.K.; Tripathi, S.K.; Jain, A.K.; Kumar, M.; Singh, T.B. Association of Carcinoma of the Gallbladder with Typhoid Carriage in a Typhoid Endemic Area Using Nested PCR. J. Infect. Dev. Ctries 2008, 2, 302–307. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhu, S.; Zhao, J.-H.; Bao, H.-X.; Liu, H.; Ding, T.-M.; Liu, G.-R.; Li, Y.-G.; Johnston, R.N.; Cao, F.-L.; et al. Genetic Boundaries Delineate the Potential Human Pathogen Salmonella Bongori into Discrete Lineages: Divergence and Speciation. BMC Genom. 2019, 20, 930. [Google Scholar] [CrossRef]
- Parkhill, J.; Dougan, G.; James, K.D.; Thomson, N.R.; Pickard, D.; Wain, J.; Churcher, C.; Mungall, K.L.; Bentley, S.D.; Holden, M.T.G.; et al. Complete Genome Sequence of a Multiple Drug Resistant Salmonella Enterica Serovar Typhi CT18. Nature 2001, 413, 848–852. [Google Scholar] [CrossRef]
- Tabak, M.; Scher, K.; Chikindas, M.L.; Yaron, S. The Synergistic Activity of Triclosan and Ciprofloxacin on Biofilms of Salmonella Typhimurium. FEMS Microbiol. Lett. 2009, 301, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Caygill, C.P.; Hill, M.J.; Braddick, M.; Sharp, J.C. Cancer Mortality in Chronic Typhoid and Paratyphoid Carriers. Lancet 1994, 343, 83–84. [Google Scholar] [CrossRef]
- Dutta, U.; Garg, P.K.; Kumar, R.; Tandon, R.K. Typhoid Carriers among Patients with Gallstones Are at Increased Risk for Carcinoma of the Gallbladder. Am. J. Gastroenterol. 2000, 95, 784–787. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S.; Marshall, J.M.; Baker, S.; Dongol, S.; Charles, R.C.; Ryan, E.T. Salmonella Chronic Carriage: Epidemiology, Diagnosis and Gallbladder Persistence. Trends Microbiol. 2014, 22, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Systematic Review with Meta-Analysis: The Relationship between Chronic Salmonella Typhi Carrier Status and Gall-Bladder Cancer-Nagaraja-2014-Alimentary Pharmacology &Amp; Therapeutics-Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/apt.12655 (accessed on 6 February 2022).
- Parry, C.M.; Hien, T.T.; Dougan, G.; White, N.J.; Farrar, J.J. Typhoid Fever. N. Engl. J. Med. 2002, 347, 1770–1782. [Google Scholar] [CrossRef] [Green Version]
- Reeve, K. Salmonella Binding to and Biofilm Formation on Cholesterol/Gallstone Surfaces in the Chronic Carrier State; Ohio State University: Columbus, OH, USA, 2010. [Google Scholar]
- Gunn, J.S. Mechanisms of Bacterial Resistance and Response to Bile. Microbes Infect. 2000, 2, 907–913. [Google Scholar] [CrossRef]
- Stress Responses as Determinants of Antimicrobial Resistance in Gram-Negative Bacteria—ScienceDirect. Available online: https://www.sciencedirect.com/science/article/abs/pii/S0966842X12000261 (accessed on 28 March 2022).
- Walawalkar, Y.D.; Vaidya, Y.; Nayak, V. Response of Salmonella Typhi to Bile-Generated Oxidative Stress: Implication of Quorum Sensing and Persister Cell Populations. Pathog. Dis. 2016, 74, ftw090. [Google Scholar] [CrossRef] [Green Version]
- Begley, M.; Gahan, C.G.M.; Hill, C. The Interaction between Bacteria and Bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [Green Version]
- Merritt, M.E.; Donaldson, J.R. Effect of Bile Salts on the DNA and Membrane Integrity of Enteric Bacteria. J. Med. Microbiol. 2009, 58, 1533–1541. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.; Ravenhall, M.; Pickard, D.; Dougan, G.; Byrne, A.; Frankel, G. Comparison of Salmonella Enterica Serovars Typhi and Typhimurium Reveals Typhoidal Serovar-Specific Responses to Bile. Infect. Immun. 2018, 86, e00490-17. [Google Scholar] [CrossRef] [Green Version]
- Hernández, S.B.; Cota, I.; Ducret, A.; Aussel, L.; Casadesús, J. Adaptation and Preadaptation of Salmonella Enterica to Bile. PLoS Genet. 2012, 8, e1002459. [Google Scholar] [CrossRef] [Green Version]
- Oxidative Stress Responses in Escherichia Coli and Salmonella Typhimurium. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC372838/ (accessed on 4 February 2022).
- Tsolis, R.M.; Bäumler, A.J.; Heffron, F. Role of Salmonella Typhimurium Mn-Superoxide Dismutase (SodA) in Protection against Early Killing by J774 Macrophages. Infect. Immun. 1995, 63, 1739–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, R.W.; Gibson, D.L.; Kay, W.W.; Gunn, J.S. Identification of a Bile-Induced Exopolysaccharide Required for Salmonella Biofilm Formation on Gallstone Surfaces. Infect. Immun. 2008, 76, 5341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, C.W.; Chan, R.C.Y.; Cheng, A.F.B.; Sung, J.Y.; Leung, J. Common Bile Duct Stones: A Cause of Chronic Salmonellosis. Am. J. Gastroenterol. 1992, 87, 1198–1199. [Google Scholar] [PubMed]
- Schirmer, B.D.; Winters, K.L.; Edlich, R.F. Cholelithiasis and Cholecystitis. J. Long Term Eff. Med. Implants. 2005, 15, 329–338. [Google Scholar] [CrossRef]
- Karaki, K.; Matsubara, Y. Surgical treatment of chronic biliary typhoid and paratyphoid carriers. Nihon Shokakibyo Gakkai Zasshi 1984, 81, 2978–2985. [Google Scholar]
- Schiøler, H.; Christiansen, E.D.; Høybye, G.; Rasmussen, S.N.; Greibe, J. Biliary Calculi in Chronic Salmonella Carriers and Healthy Controls: A Controlled Study. Scand. J. Infect. Dis. 1983, 15, 17–19. [Google Scholar] [CrossRef]
- Surette, M.G.; Bassler, B.L. Regulation of Autoinducer Production in Salmonella Typhimurium. Mol. Microbiol. 1999, 31, 585–595. [Google Scholar] [CrossRef] [Green Version]
- Maurer, K.J.; Carey, M.C.; Fox, J.G. Roles of Infection, Inflammation, and the Immune System in Cholesterol Gallstone Formation. Gastroenterology 2009, 136, 425–440. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.L.; Zhang, S.; Tsolis, R.M.; Kingsley, R.A.; Adams, L.G.; Bäumler, A.J. Animal Models of Salmonella Infections: Enteritis versus Typhoid Fever. Microbes Infect. 2001, 3, 1335–1344. [Google Scholar] [CrossRef]
- Gonzalez-Escobedo, G.; Marshall, J.M.; Gunn, J.S. Chronic and Acute Infection of the Gall Bladder by Salmonella Typhi: Understanding the Carrier State. Nat. Rev. Microbiol. 2011, 9, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Prouty, A.M.; Schwesinger, W.H.; Gunn, J.S. Biofilm Formation and Interaction with the Surfaces of Gallstones by Salmonella Spp. Infect. Immun. 2002, 70, 2640–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratt, L.A.; Kolter, R. Genetic Analysis of Escherichia Coli Biofilm Formation: Roles of Flagella, Motility, Chemotaxis and Type I Pili. Mol. Microbiol. 1998, 30, 285–293. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A.; Kolter, R. Flagellar and Twitching Motility Are Necessary for Pseudomonas Aeruginosa Biofilm Development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial Biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef]
- Winter, S.E.; Raffatellu, M.; Wilson, R.P.; Rüssmann, H.; Bäumler, A.J. The Salmonella Enterica Serotype Typhi Regulator TviA Reduces Interleukin-8 Production in Intestinal Epithelial Cells by Repressing Flagellin Secretion. Cell. Microbiol. 2008, 10, 247–261. [Google Scholar] [CrossRef]
- Crawford, R.W.; Reeve, K.E.; Gunn, J.S. Flagellated but Not Hyperfimbriated Salmonella Enterica Serovar Typhimurium Attaches to and Forms Biofilms on Cholesterol-Coated Surfaces. J. Bacteriol. 2010, 192, 2981–2990. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.L.; White, A.P.; Snyder, S.D.; Martin, S.; Heiss, C.; Azadi, P.; Surette, M.; Kay, W.W. Salmonella Produces an O-Antigen Capsule Regulated by AgfD and Important for Environmental Persistence. J. Bacteriol. 2006, 188, 7722–7730. [Google Scholar] [CrossRef] [Green Version]
- Jonas, K.; Tomenius, H.; Kader, A.; Normark, S.; Römling, U.; Belova, L.M.; Melefors, O. Roles of Curli, Cellulose and BapA in Salmonella Biofilm Morphology Studied by Atomic Force Microscopy. BMC Microbiol. 2007, 7, 70. [Google Scholar] [CrossRef] [Green Version]
- Adcox, H.E.; Vasicek, E.M.; Dwivedi, V.; Hoang, K.V.; Turner, J.; Gunn, J.S. Salmonella Extracellular Matrix Components Influence Biofilm Formation and Gallbladder Colonization. Infect. Immun. 2016, 84, 3243–3251. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Gu, L.; Geng, X.; Xu, G.; Huang, X.; Zhu, X. A Novel Cis Antisense RNA AsfD Promotes Salmonella Enterica Serovar Typhi Motility and Biofilm Formation. Microb. Pathog. 2020, 142, 104044. [Google Scholar] [CrossRef]
- Grandvalet, C.; Assad-García, J.S.; Chu-Ky, S.; Tollot, M.; Guzzo, J.; Gresti, J.; Tourdot-Maréchal, R. Changes in Membrane Lipid Composition in Ethanol- and Acid-Adapted Oenococcus Oeni Cells: Characterization of the Cfa Gene by Heterologous Complementation. Microbiol. Read. 2008, 154, 2611–2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Liu, R.; Tang, H.; Osei-Adjei, G.; Xu, S.; Zhang, Y.; Huang, X. A 3’ UTR-Derived Non-Coding RNA RibS Increases Expression of Cfa and Promotes Biofilm Formation of Salmonella Enterica Serovar Typhi. Res. Microbiol. 2018, 169, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Que, F.; Liao, L.; Zhou, M.; You, L.; Zhao, Q.; Li, Y.; Niu, H.; Wu, S.; Huang, R. Study on the Promotion of Bacterial Biofilm Formation by a Salmonella Conjugative Plasmid and the Underlying Mechanism. PLoS ONE 2014, 9, e109808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zautner, A.E.; Hage, A.; Schneider, K.; Schlösser, K.; Zimmermann, O.; Hornecker, E.; Mausberg, R.F.; Frickmann, H.; Groß, U.; Ziebolz, D. Effects of Easy-to-Perform Procedures to Reduce Bacterial Colonization with Streptococcus Mutans and Staphylococcus Aureus on Toothbrushes. Eur. J. Microbiol. Immunol. 2013, 3, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; Yang, Y.; Li, Y.; Ye, Y.; Yan, J.; Wang, T.; Shuyan, W.; Huang, R. A Salmonella Enterica Conjugative Plasmid Impairs Autophagic Flux in Infected Macrophages. Microbes Infect. 2014, 16, 553–561. [Google Scholar] [CrossRef]
- Huang, R.; Wu, S.; Wen, Y. Studies on virulence mediated by drug resistant Salmonella typhi R plasmid. Chin. J. Microbiol. Immunol. 2001, 12, 302–305. [Google Scholar]
- Wu, S.; Chu, Y.; Yang, Y.; Li, Y.; He, P.; Zheng, Y.; Zhang, C.; Liu, Q.; Han, L.; Huang, R. Inhibition of Macrophage Autophagy Induced by Salmonella Enterica Serovar Typhi Plasmid. Front. Biosci.-Landmark 2014, 19, 490–503. [Google Scholar] [CrossRef] [Green Version]
- Brodsky, I.; Ghori, N.; Falkow, S.; Monack, D. Mig-14 Is an Inner Membrane-Associated Protein That Promotes Salmonella Typhimurium Resistance to CRAMP, Survival within Activated Macrophages and Persistent Infection. Mol. Microbiol. 2005, 55, 954–972. [Google Scholar] [CrossRef]
- Ochman, H.; Groisman, E.A. Distribution of Pathogenicity Islands in Salmonella Spp. Infect. Immun. 1996, 64, 5410–5412. [Google Scholar] [CrossRef] [Green Version]
- Sheng, X.; Wang, W.; Chen, L.; Zhang, H.; Zhang, Y.; Xu, S.; Xu, H.; Huang, X. Mig-14 May Contribute to Salmonella Enterica Serovar Typhi Resistance to Polymyxin B by Decreasing the Permeability of the Outer-Membrane and Promoting the Formation of Biofilm. Int. J. Med. Microbiol. 2019, 309, 143–150. [Google Scholar] [CrossRef]
- Brodsky, I.E.; Ernst, R.K.; Miller, S.I.; Falkow, S. Mig-14 Is a Salmonella Gene That Plays a Role in Bacterial Resistance to Antimicrobial Peptides. J. Bacteriol. 2002, 184, 3203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danese, P.N.; Pratt, L.A.; Kolter, R. Exopolysaccharide Production Is Required for Development of Escherichia Coli K-12 Biofilm Architecture. J. Bacteriol. 2000, 182, 3593–3596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevenson, G.; Andrianopoulos, K.; Hobbs, M.; Reeves, P.R. Organization of the Escherichia Coli K-12 Gene Cluster Responsible for Production of the Extracellular Polysaccharide Colanic Acid. J. Bacteriol. 1996, 178, 4885–4893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rana, K.; Nayak, S.R.; Bihary, A.; Sahoo, A.K.; Mohanty, K.C.; Palo, S.K.; Sahoo, D.; Pati, S.; Dash, P. Association of Quorum Sensing and Biofilm Formation with Salmonella Virulence: Story beyond Gathering and Cross-Talk. Arch. Microbiol. 2021, 203, 5887–5897. [Google Scholar] [CrossRef]
- Sholpan, A.; Lamas, A.; Cepeda, A.; Franco, C.M. Salmonella Spp. Quorum Sensing: An Overview from Environmental Persistence to Host Cell Invasion. AIMS Microbiol. 2021, 7, 238–256. [Google Scholar] [CrossRef]
- Perrett, C.A.; Karavolos, M.H.; Humphrey, S.; Mastroeni, P.; Martinez-Argudo, I.; Spencer, H.; Jepson, M.A. LuxS-Based Quorum Sensing Does Not Affect the Ability of Salmonella enterica Serovar Typhimurium to Express the SPI-1 Type 3 Secretion System, Induce Membrane Ruffles, or Invade Epithelial Cells. J. Bacteriol. 2009, 191, 7253–7259. [Google Scholar] [CrossRef] [Green Version]
- Moreira, C.G.; Weinshenker, D.; Sperandio, V. QseC Mediates Salmonella enterica Serovar Typhimurium Virulence In Vitro and In Vivo. Infect. Immun. 2009, 78, 914–926. [Google Scholar] [CrossRef] [Green Version]
- Widmer, K.W.; Jesudhasan, P.; Pillai, S.D. Fatty Acid Modulation of Autoinducer (AI-2) Influenced Growth and Macrophage Invasion by Salmonella Typhimurium. Foodborne Pathog. Dis. 2012, 9, 211–217. [Google Scholar] [CrossRef]
- Jesudhasan, P.R.; Cepeda, M.L.; Widmer, K.; Dowd, S.E.; Soni, K.A.; Hume, M.E.; Zhu, J.; Pillai, S.D. Transcriptome Analysis of Genes Controlled by LuxS/Autoinducer-2 in Salmonella Enterica Serovar Typhimurium. Foodborne Pathog. Dis. 2010, 7, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Kint, G.; De Coster, D.; Marchal, K.; Vanderleyden, J.; De Keersmaecker, S.C.J. The Small Regulatory RNA Molecule MicA Is Involved in Salmonella Enterica Serovar Typhimurium Biofilm Formation. BMC Microbiol. 2010, 10, 276. [Google Scholar] [CrossRef] [Green Version]
- Michael, B.; Smith, J.; Swift, S.; Heffron, F.; Ahmer, B. SdiA of Salmonella Enterica Is a LuxR Homolog That Detects Mixed Microbial Communities. J. Bacteriol. 2001, 183, 5733–5742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutsukake, K.; Ohya, Y.; Iino, T. Transcriptional Analysis of the Flagellar Regulon of Salmonella Typhimurium. J. Bacteriol. 1990, 172, 741–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutsukake, K.; Iino, T. Role of the FliA-FlgM regulatory system on the transcriptional control of the flagellar regulon and flagellar formation in Salmonella typhimurium. J. Bacteriol. 1994, 176, 3598–3605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.; Li, W.; Zhang, Y.; Chen, L.; Zhang, Y.; Zheng, X.; Ni, B. QseB mediates biofilm formation and invasion in Salmonella enterica serovar Typhi. Microb. Pathog. 2017, 104, 6–11. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jahan, F.; Chinni, S.V.; Samuggam, S.; Reddy, L.V.; Solayappan, M.; Su Yin, L. The Complex Mechanism of the Salmonella typhi Biofilm Formation That Facilitates Pathogenicity: A Review. Int. J. Mol. Sci. 2022, 23, 6462. https://doi.org/10.3390/ijms23126462
Jahan F, Chinni SV, Samuggam S, Reddy LV, Solayappan M, Su Yin L. The Complex Mechanism of the Salmonella typhi Biofilm Formation That Facilitates Pathogenicity: A Review. International Journal of Molecular Sciences. 2022; 23(12):6462. https://doi.org/10.3390/ijms23126462
Chicago/Turabian StyleJahan, Fahmida, Suresh V. Chinni, Sumitha Samuggam, Lebaka Veeranjaneya Reddy, Maheswaran Solayappan, and Lee Su Yin. 2022. "The Complex Mechanism of the Salmonella typhi Biofilm Formation That Facilitates Pathogenicity: A Review" International Journal of Molecular Sciences 23, no. 12: 6462. https://doi.org/10.3390/ijms23126462
APA StyleJahan, F., Chinni, S. V., Samuggam, S., Reddy, L. V., Solayappan, M., & Su Yin, L. (2022). The Complex Mechanism of the Salmonella typhi Biofilm Formation That Facilitates Pathogenicity: A Review. International Journal of Molecular Sciences, 23(12), 6462. https://doi.org/10.3390/ijms23126462