
IL-31: State of the Art for an Inflammation-Oriented Interleukin

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
1.1. IL-31 Structure
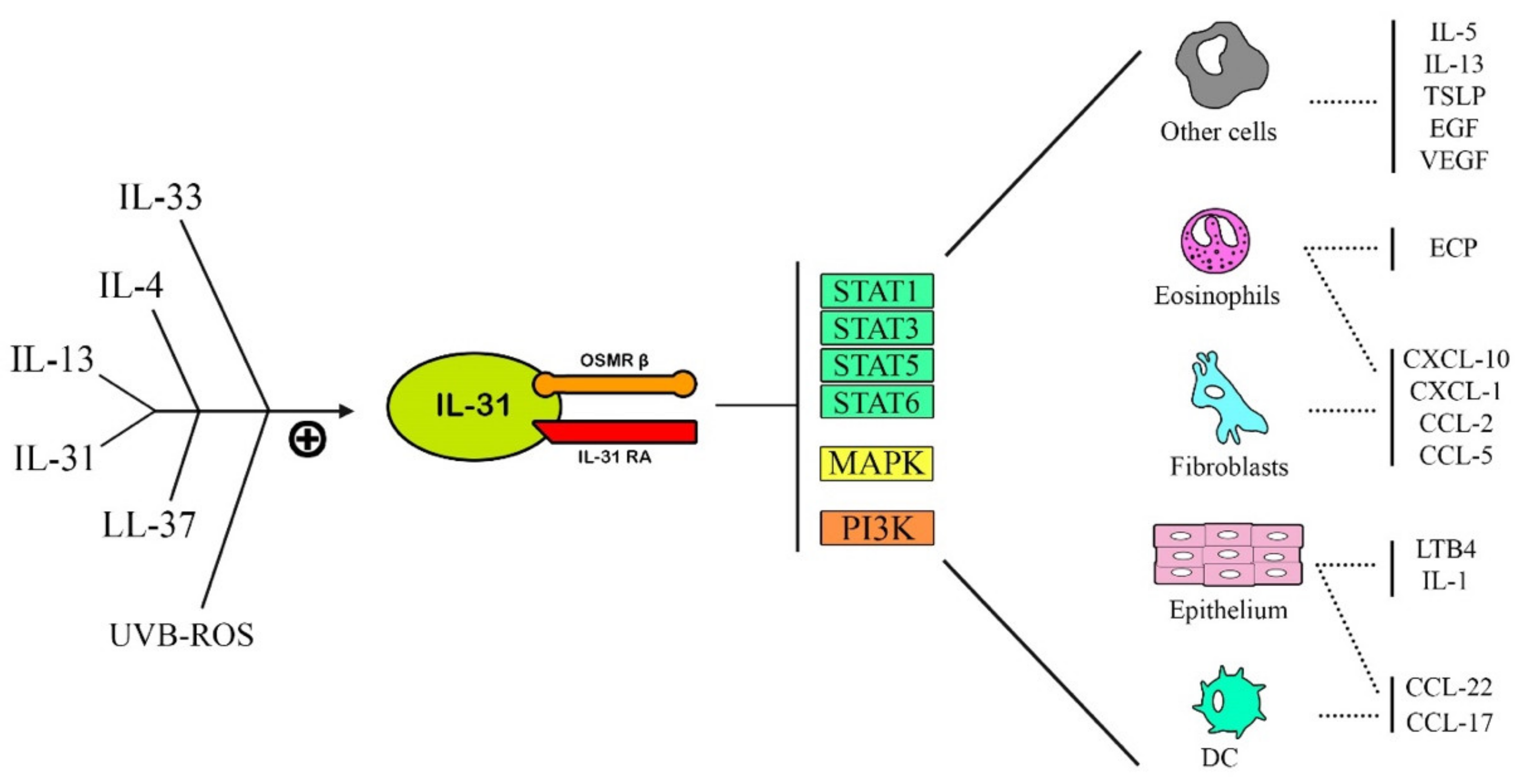
1.2. IL-31 Release Mechanism
1.3. IL-31 Receptors
1.4. IL-31 Function
- (1)
- JAK/STAT pathway (Janus-activated kinase/signal transducer and activator of transcription),
- (2)
- PI3K/AKT (phosphatidylinositol 3′-kinase/protein kinase) pathway,
- (3)
- MAPK-JNK/p38 (mitogen-activated protein kinase—Janus kinase/p38) pathway.

2. Discussion
2.1. IL-31: Role in Skin Diseases
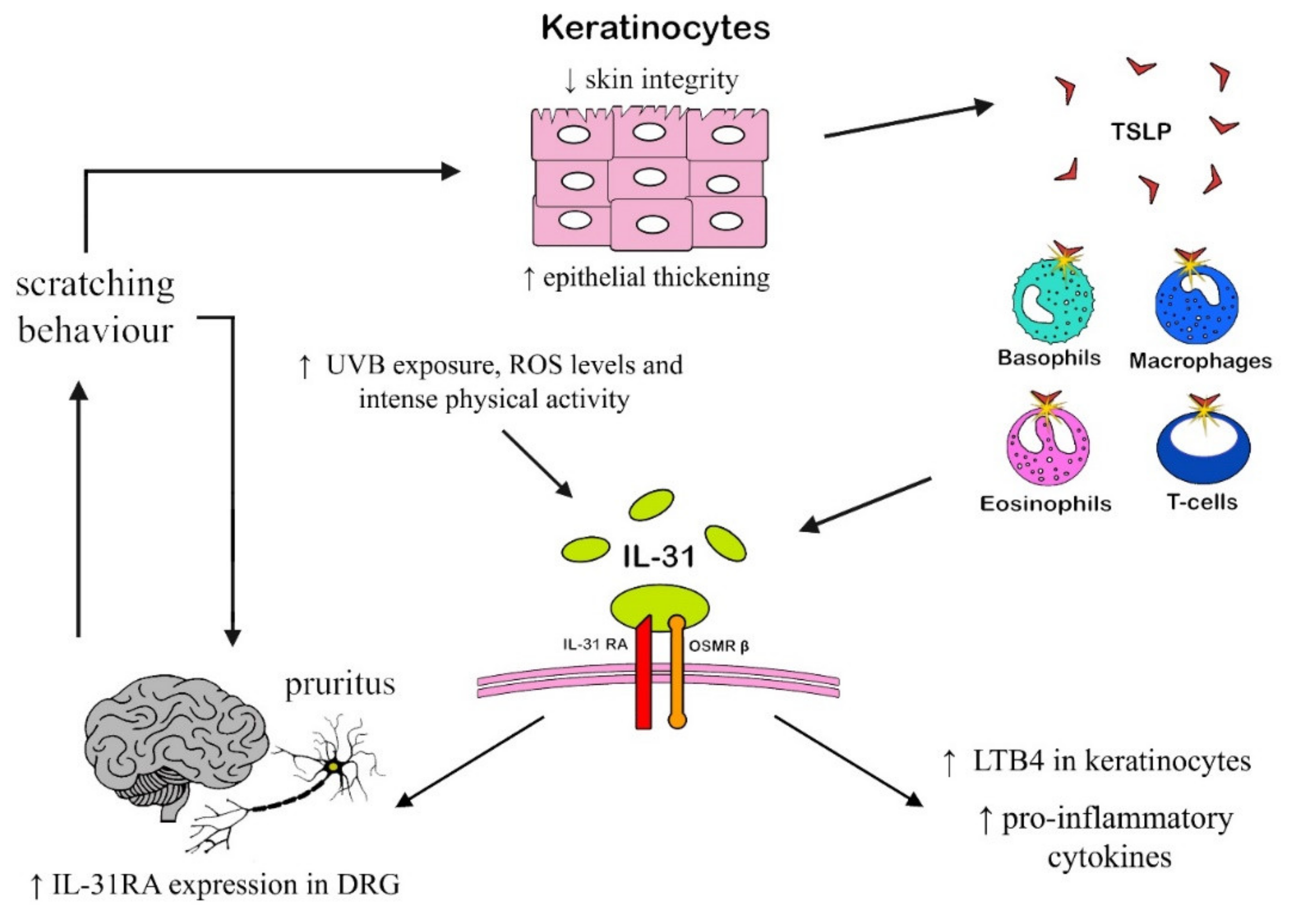
2.1.1. Atopic Dermatitis
2.1.2. Psoriasis
2.1.3. Chronic itch
2.2. IL-31: Role in Allergic Diseases
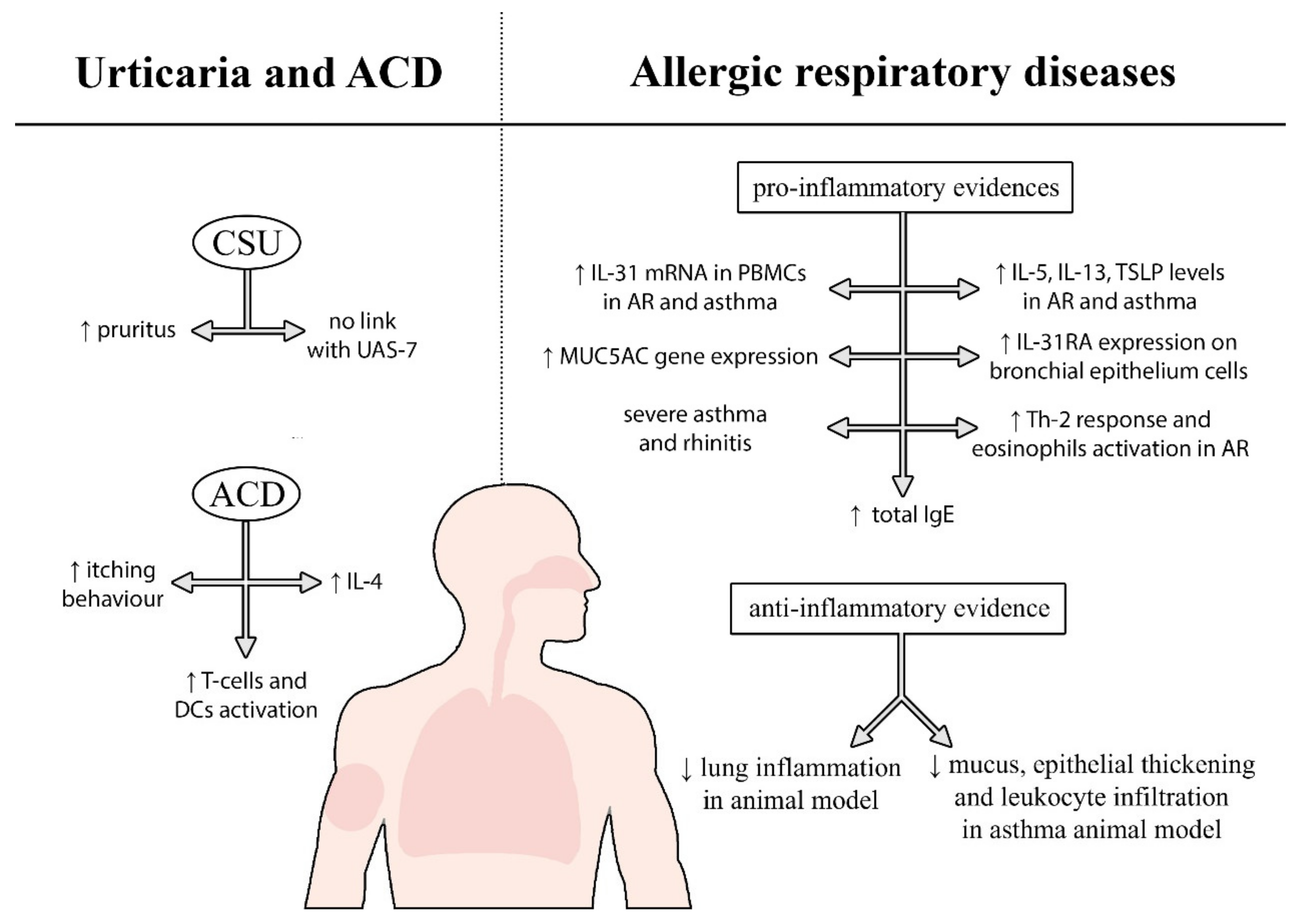
2.2.1. Allergic Respiratory Diseases
2.2.2. Urticaria and Allergic-Contact Dermatitis
2.3. IL-31 in Hematological Diseases
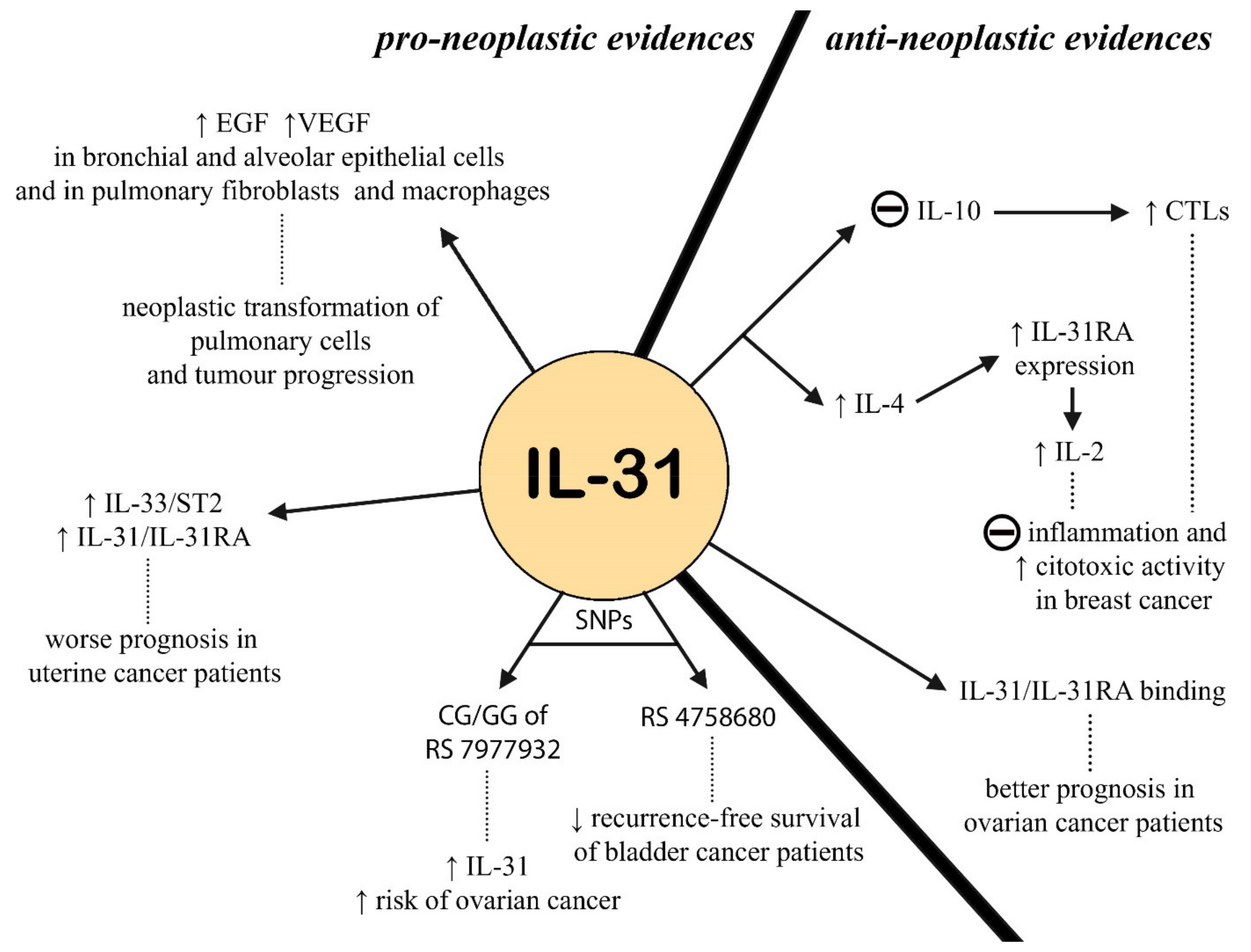
2.4. IL-31 in Oncology
2.5. IL-31 in Infectious Diseases
2.6. Miscellaneous
3. IL-31: Target for Current and Future Therapies
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dillon, S.R.; Sprecher, C.; Hammond, A.; Bilsborough, J.; Rosenfeld-Franklin, M.; Presnell, S.R.; Haugen, H.S.; Maurer, M.; Harder, B.; Johnston, J.; et al. Interleukin 31, a Cytokine Produced by Activated T Cells, Induces Dermatitis in Mice. Nat. Immunol. 2004, 5, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Duca, E.; Sur, G.; Armat, I.; Samasca, G.; Sur, L. Correlation between Interleukin 31 and Clinical Manifestations in Children with Atopic Dermatitis: An Observational Study. Allergol. Et Immunopathol. 2022, 50, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Tada, H.; Nishioka, T.; Takase, A.; Numazaki, K.; Bando, K.; Matsushita, K. Porphyromonas gingivalis Induces the Production of Interleukin-31 by Human Mast Cells, Resulting in Dysfunction of the Gingival Epithelial Barrier. Cell. Microbiol. 2019, 21, e12972. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, H.-J.; Yang, H.S.; Kim, E.; Huh, I.-S.; Yang, J.-M. IL-31 Serum Protein and Tissue MRNA Levels in Patients with Atopic Dermatitis. Ann. Dermatol. 2011, 23, 468. [Google Scholar] [CrossRef]
- Schulz, F.; Marenholz, I.; Fölster-Holst, R.; Chen, C.; Sternjak, A.; Baumgrass, R.; Esparza-Gordillo, J.; Grüber, C.; Nickel, R.; Schreiber, S.; et al. A Common Haplotype of the IL-31 Gene Influencing Gene Expression Is Associated with Nonatopic Eczema. J. Allergy Clin. Immunol. 2007, 120, 1097–1102. [Google Scholar] [CrossRef]
- Dreuw, A.; Radtke, S.; Pflanz, S.; Lippok, B.E.; Heinrich, P.C.; Hermanns, H.M. Characterization of the Signaling Capacities of the Novel Gp130-like Cytokine Receptor. J. Biol. Chem. 2004, 279, 36112–36120. [Google Scholar] [CrossRef]
- Venereau, E.; Diveu, C.; Grimaud, L.; Ravon, E.; Froger, J.; Preisser, L.; Danger, Y.; Maillasson, M.; Garrigue-Antar, L.; Jacques, Y.; et al. Definition and Characterization of an Inhibitor for Interleukin-31. J. Biol. Chem. 2010, 285, 14955–14963. [Google Scholar] [CrossRef]
- Jurca, T.; Baldea, I.; Filip, G.A.; Olteanu, D.; Clichici, S.; Pallag, A.; Vicaş, L.; Marian, E.; Micle, O.; Crivii, C.B.; et al. A Phytocomplex Consisting of Tropaeolum majus L. and Salvia officinalis L. Extracts Alleviates the Inflammatory Response of Dermal Fibroblasts to Bacterial Lipopolysaccharides. Oxidative Med. Cell. Longev. 2020, 2020, 8516153. [Google Scholar] [CrossRef]
- Murdaca, G.; Greco, M.; Tonacci, A.; Negrini, S.; Borro, M.; Puppo, F.; Gangemi, S. IL-33/IL-31 Axis in Immune-Mediated and Allergic Diseases. Int. J. Mol. Sci. 2019, 20, 5856. [Google Scholar] [CrossRef]
- Gangemi, S.; Quartuccio, S.; Casciaro, M.; Trapani, G.; Minciullo, P.L.; Imbalzano, E. Interleukin 31 and Skin Diseases: A Systematic Review. Allergy Asthma Proc. 2017, 38, 401–408. [Google Scholar] [CrossRef]
- Stott, B.; Lavender, P.; Lehmann, S.; Pennino, D.; Durham, S.; Schmidt-Weber, C.B. Human IL-31 Is Induced by IL-4 and Promotes TH2-Driven Inflammation. J. Allergy Clin. Immunol. 2013, 132, 446–454.e5. [Google Scholar] [CrossRef]
- Sonkoly, E.; Muller, A.; Lauerma, A.I.; Pivarcsi, A.; Soto, H.; Kemeny, L.; Alenius, H.; Dieu-Nosjean, M.-C.; Meller, S.; Rieker, J.; et al. IL-31: A New Link between T Cells and Pruritus in Atopic Skin Inflammation. J. Allergy Clin. Immunol. 2006, 117, 411–417. [Google Scholar] [CrossRef]
- Guarneri, F.; Minciullo, P.L.; Mannucci, C.; Calapai, F.; Saitta, S.; Cannavò, S.P.; Gangemi, S. IL-31 and IL-33 Circulating Levels in Allergic Contact Dermatitis. Eur. Ann. Allergy Clin. Immunol. 2015, 47, 156–158. [Google Scholar]
- Olomski, F.; Fettelschoss, V.; Jonsdottir, S.; Birkmann, K.; Thoms, F.; Marti, E.; Bachmann, M.F.; Kündig, T.M.; Fettelschoss-Gabriel, A. Interleukin 31 in Insect Bite Hypersensitivity—Alleviating Clinical Symptoms by Active Vaccination against Itch. Allergy 2020, 75, 862–871. [Google Scholar] [CrossRef]
- Huang, J.; Yue, H.; Jiang, T.; Gao, J.; Shi, Y.; Shi, B.; Wu, X.; Gou, X. IL-31 Plays Dual Roles in Lung Inflammation in an OVA-Induced Murine Asthma Model. Biol. Open 2019, 8, bio036244. [Google Scholar] [CrossRef]
- Wong, C.-K.; Leung, K.M.-L.; Qiu, H.-N.; Chow, J.Y.-S.; Choi, A.O.K.; Lam, C.W.-K. Activation of Eosinophils Interacting with Dermal Fibroblasts by Pruritogenic Cytokine IL-31 and Alarmin IL-33: Implications in Atopic Dermatitis. PLoS ONE 2012, 7, e29815. [Google Scholar] [CrossRef]
- Horejs-Hoeck, J.; Schwarz, H.; Lamprecht, S.; Maier, E.; Hainzl, S.; Schmittner, M.; Posselt, G.; Stoecklinger, A.; Hawranek, T.; Duschl, A. Dendritic Cells Activated by IFN-γ/STAT1 Express IL-31 Receptor and Release Proinflammatory Mediators upon IL-31 Treatment. J. Immunol. 2012, 188, 5319–5326. [Google Scholar] [CrossRef]
- Lan, C.-C.E.; Tu, H.-P.; Wu, C.-S.; Ko, Y.-C.; Yu, H.-S.; Lu, Y.-W.; Li, W.-C.; Chen, Y.-C.; Chen, G.-S. Distinct SPINK5 and IL-31 Polymorphisms Are Associated with Atopic Eczema and Non-Atopic Hand Dermatitis in Taiwanese Nursing Population. Exp. Dermatol. 2011, 20, 975–979. [Google Scholar] [CrossRef]
- Miake, S.; Tsuji, G.; Takemura, M.; Hashimoto-Hachiya, A.; Vu, Y.H.; Furue, M.; Nakahara, T. IL-4 Augments IL-31/IL-31 Receptor Alpha Interaction Leading to Enhanced Ccl 17 and Ccl 22 Production in Dendritic Cells: Implications for Atopic Dermatitis. Int. J. Mol. Sci. 2019, 20, 4053. [Google Scholar] [CrossRef]
- Hofbauer, G.; Nobbe, S.; Dziunycz, P.; Mühleisen, B.; Bilsborough, J.; Dillon, S.; French, L. IL-31 Expression by Inflammatory Cells Is Preferentially Elevated in Atopic Dermatitis. Acta Derm. Venereol. 2012, 92, 24–28. [Google Scholar] [CrossRef]
- Raap, U.; Weißmantel, S.; Gehring, M.; Eisenberg, A.M.; Kapp, A.; Fölster-Holst, R. IL-31 Significantly Correlates with Disease Activity and Th2 Cytokine Levels in Children with Atopic Dermatitis. Pediatric Allergy Immunol. 2012, 23, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Byeon, J.H.; Yoon, W.; Ahn, S.H.; Lee, H.S.; Kim, S.; Yoo, Y. Correlation of Serum Interleukin-31 with Pruritus and Blood Eosinophil Markers in Children with Atopic Dermatitis. Allergy Asthma Proc. 2020, 41, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Ozceker, D.; Bulut, M.; Ozbay, A.C.; Dilek, F.; Koser, M.; Tamay, Z.; Guler, N. Assessment of IL-31 Levels and Disease Severity in Children with Atopic Dermatitis. Allergol. Et Immunopathol. 2018, 46, 322–325. [Google Scholar] [CrossRef] [PubMed]
- Siniewicz-Luzeńczyk, K.; Stańczyk-Przyłuska, A.; Zeman, K. Correlation between Serum Interleukin 31 Level and the Severity of Disease in Children with Atopic Dermatitis. Adv. Dermatol. Allergol. 2013, 5, 282–285. [Google Scholar] [CrossRef]
- Ezzat, M.; Hasan, Z.; Shaheen, K. Serum Measurement of Interleukin-31 (IL-31) in Paediatric Atopic Dermatitis: Elevated Levels Correlate with Severity Scoring. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 334–339. [Google Scholar] [CrossRef]
- Kasraie, S.; Niebuhr, M.; Werfel, T. Interleukin (IL)-31 Activates Signal Transducer and Activator of Transcription (STAT)-1, STAT-5 and Extracellular Signal-Regulated Kinase 1/2 and down-Regulates IL-12p40 Production in Activated Human Macrophages. Allergy 2013, 68, 739–747. [Google Scholar] [CrossRef]
- van Drongelen, V.; Haisma, E.M.; Out-Luiting, J.J.; Nibbering, P.H.; el Ghalbzouri, A. Reduced Filaggrin Expression Is Accompanied by Increased Staphylococcus aureus Colonization of Epidermal Skin Models. Clin. Exp. Allergy 2014, 44, 1515–1524. [Google Scholar] [CrossRef]
- Kanda, N.; Hau, C.S.; Tada, Y.; Sato, S.; Watanabe, S. Decreased Serum LL-37 and Vitamin D3 Levels in Atopic Dermatitis: Relationship between IL-31 and Oncostatin M. Allergy 2012, 67, 804–812. [Google Scholar] [CrossRef]
- Cheon, B.R.; Shin, J.E.; Kim, Y.J.; Shim, J.W.; Kim, D.S.; Jung, H.L.; Park, M.S.; Shim, J.Y. Relationship between Serum 25-Hydroxyvitamin D and Interleukin-31 Levels, and the Severity of Atopic Dermatitis in Children. Korean J. Pediatrics 2015, 58, 96. [Google Scholar] [CrossRef]
- Yamamura, K.; Uruno, T.; Shiraishi, A.; Tanaka, Y.; Ushijima, M.; Nakahara, T.; Watanabe, M.; Kido-Nakahara, M.; Tsuge, I.; Furue, M.; et al. The Transcription Factor EPAS1 Links DOCK8 Deficiency to Atopic Skin Inflammation via IL-31 Induction. Nat. Commun. 2017, 8, 13946. [Google Scholar] [CrossRef]
- Tang, L.; Gao, J.; Cao, X.; Chen, L.; Wang, H.; Ding, H. TRPV1 Mediates Itch-associated Scratching and Skin Barrier Dysfunction in DNFB-induced Atopic Dermatitis Mice. Exp. Dermatol. 2022, 31, 398–405. [Google Scholar] [CrossRef]
- Kim, S.-H.; Kim, E.-K.; Choi, E.-J. High-Intensity Swimming Exercise Increases Dust Mite Extract and 1-Chloro-2,4-Dinitrobenzene-Derived Atopic Dermatitis in BALB/c Mice. Inflammation 2014, 37, 1179–1185. [Google Scholar] [CrossRef]
- Grimstad, Ø.; Sawanobori, Y.; Vestergaard, C.; Bilsborough, J.; Olsen, U.B.; Grønhøj-Larsen, C.; Matsushima, K. Anti-Interleukin-31-Antibodies Ameliorate Scratching Behaviour in NC/Nga Mice: A Model of Atopic Dermatitis. Exp. Dermatol. 2009, 18, 35–43. [Google Scholar] [CrossRef]
- Ningombam, A.; Handa, S.; Srivastava, N.; Mahajan, R.; De, D. Addition of Oral Fexofenadine to Topical Therapy Leads to a Significantly Greater Reduction in the Serum Interleukin-31 Levels in Mild to Moderate Paediatric Atopic Dermatitis. Clin. Exp. Dermatol. 2022, 47, 724–729. [Google Scholar] [CrossRef]
- Cornelissen, C.; Brans, R.; Czaja, K.; Skazik, C.; Marquardt, Y.; Zwadlo-Klarwasser, G.; Kim, A.; Bickers, D.R.; Lüscher-Firzlaff, J.; Lüscher, B.; et al. Ultraviolet B Radiation and Reactive Oxygen Species Modulate Interleukin-31 Expression in T Lymphocytes, Monocytes and Dendritic Cells. Br. J. Dermatol. 2011, 165, 966–975. [Google Scholar] [CrossRef]
- Gambichler, T.; Kreuter, A.; Tomi, N.S.; Othlinghaus, N.; Altmeyer, P.; Skrygan, M. Gene Expression of Cytokines in Atopic Eczema before and after Ultraviolet A1 Phototherapy. Br. J. Dermatol. 2008, 158, 1117–1120. [Google Scholar] [CrossRef]
- Bogaczewicz, J.; Malinowska, K.; Sysa-Jedrzejowska, A.; Wozniacka, A. Medium-Dose Ultraviolet A1 Phototherapy Improves SCORAD Index and Increases MRNA Expression of Interleukin-4 without Direct Effect on Human β Defensin-1, Interleukin-10, and Interleukin-31. Int. J. Dermatol. 2016, 55, e380–e385. [Google Scholar] [CrossRef]
- Kang, M.; Cho, K.; Lee, J.; Subedi, L.; Yumnam, S.; Kim, S. Effect of Resveratrol-Enriched Rice on Skin Inflammation and Pruritus in the NC/Nga Mouse Model of Atopic Dermatitis. Int. J. Mol. Sci. 2019, 20, 1428. [Google Scholar] [CrossRef]
- Lee, J.W.; Wu, Q.; Jang, Y.P.; Choung, S.Y. Pinus Densiflora Bark Extract Ameliorates 2,4-Dinitrochlorobenzene-Induced Atopic Dermatitis in NC/Nga Mice by Regulating Th1/Th2 Balance and Skin Barrier Function. Phytother. Res. 2018, 32, 1135–1143. [Google Scholar] [CrossRef]
- Yu, Z.; Deng, T.; Wang, P.; Sun, T.; Xu, Y. Ameliorative Effects of Total Coumarins from the Fructus of Cnidium monnieri (L.) Cuss. on 2,4-dinitrochlorobenzene-induced Atopic Dermatitis in Rats. Phytother. Res. 2021, 35, 3310–3324. [Google Scholar] [CrossRef]
- Kim, W.-K.; Jang, Y.J.; Han, D.H.; Seo, B.; Park, S.; Lee, C.H.; Ko, G. Administration of Lactobacillus Fermentum KBL375 Causes Taxonomic and Functional Changes in Gut Microbiota Leading to Improvement of Atopic Dermatitis. Front. Mol. Biosci. 2019, 6, 92. [Google Scholar] [CrossRef]
- Nattkemper, L.A.; Tey, H.L.; Valdes-Rodriguez, R.; Lee, H.; Mollanazar, N.K.; Albornoz, C.; Sanders, K.M.; Yosipovitch, G. The Genetics of Chronic Itch: Gene Expression in the Skin of Patients with Atopic Dermatitis and Psoriasis with Severe Itch. J. Investig. Dermatol. 2018, 138, 1311–1317. [Google Scholar] [CrossRef] [PubMed]
- Narbutt, J.; Olejniczak, I.; Sobolewska-Sztychny, D.; Sysa-Jedrzejowska, A.; Słowik-Kwiatkowska, I.; Hawro, T.; Lesiak, A. Narrow Band Ultraviolet B Irradiations Cause Alteration in Interleukin-31 Serum Level in Psoriatic Patients. Arch. Dermatol. Res. 2013, 305, 191–195. [Google Scholar] [CrossRef]
- Park, S.-M.; Kim, G.-W.; Kim, H.-S.; Ko, H.-C.; Kim, M.-B.; Kim, B.-S. Characteristics of Pruritus According to Morphological Phenotype of Psoriasis and Association with Neuropeptides and Interleukin-31. Ann. Dermatol. 2020, 32, 1. [Google Scholar] [CrossRef] [PubMed]
- Chaowattanapanit, S.; Choonhakarn, C.; Salao, K.; Winaikosol, K.; Julanon, N.; Wongjirattikarn, R.; Foocharoen, C.; Sompornrattanaphan, M. Increased Serum IL-31 Levels in Chronic Spontaneous Urticaria and Psoriasis with Pruritic Symptoms. Heliyon 2020, 6, e05621. [Google Scholar] [CrossRef] [PubMed]
- Salao, K.; Sawanyawisuth, K.; Winaikosol, K.; Choonhakarn, C.; Chaowattanapanit, S. Interleukin-31 and Chronic Pruritus of Unknown Origin. Biomark. Insights 2020, 15, 117727192094071. [Google Scholar] [CrossRef]
- Arai, I.; Tsuji, M.; Miyagawa, K.; Takeda, H.; Akiyama, N.; Saito, S. Repeated Administration of IL-31 Upregulates IL-31 Receptor A (IL-31RA) in Dorsal Root Ganglia and Causes Severe Itch-Associated Scratching Behaviour in Mice. Exp. Dermatol. 2015, 24, 75–78. [Google Scholar] [CrossRef]
- Feld, M.; Garcia, R.; Buddenkotte, J.; Katayama, S.; Lewis, K.; Muirhead, G.; Hevezi, P.; Plesser, K.; Schrumpf, H.; Krjutskov, K.; et al. The Pruritus- and TH2-Associated Cytokine IL-31 Promotes Growth of Sensory Nerves. J. Allergy Clin. Immunol. 2016, 138, 500–508.e24. [Google Scholar] [CrossRef]
- Xing, Y.; Steele, H.R.; Hilley, H.B.; Zhu, Y.; Lawson, K.; Niehoff, T.; Han, L. Visualizing the Itch-Sensing Skin Arbors. J. Investig. Dermatol. 2021, 141, 1308–1316. [Google Scholar] [CrossRef]
- Andoh, T.; Harada, A.; Kuraishi, Y. Involvement of Leukotriene B4 Released from Keratinocytes in Itch-Associated Response to Intradermal Interleukin-31 in Mice. Acta Derm. Venereol. 2017, 97, 922–927. [Google Scholar] [CrossRef]
- Edukulla, R.; Singh, B.; Jegga, A.G.; Sontake, V.; Dillon, S.R.; Madala, S.K. Th2 Cytokines Augment IL-31/IL-31RA Interactions via STAT6-Dependent IL-31RA Expression. J. Biol. Chem. 2015, 290, 13510–13520. [Google Scholar] [CrossRef]
- Parker, J.C.; Thavagnanam, S.; Skibinski, G.; McBrien, M.; Heaney, L.G.; Shields, M.D. IL-31 Does Not Induce Normal Human Ciliated Epithelial Cells to Differentiate into a Phenotype Consistent with the Pathophysiology of Asthma. Results Immunol. 2012, 2, 104–111. [Google Scholar] [CrossRef]
- Lai, T.; Wu, D.; Li, W.; Chen, M.; Yi, Z.; Huang, D.; Jing, Z.; Lü, Y.; Lv, Q.; Li, D.; et al. Interleukin-31 Expression and Relation to Disease Severity in Human Asthma. Sci. Rep. 2016, 6, 22835. [Google Scholar] [CrossRef]
- Lei, Z.; Liu, G.; Huang, Q.; Lv, M.; Zu, R.; Zhang, G.-M.; Feng, Z.-H.; Huang, B. Original Article: SCF and IL-31 Rather than IL-17 and BAFF Are Potential Indicators in Patients with Allergic Asthma. Allergy 2007, 63, 327–332. [Google Scholar] [CrossRef]
- Moaaz, M.; Abo El-Nazar, S.; Abd El-Rahman, M.; Soliman, E. Stem Cell Factor and Interleukin-31 Expression: Association with IgE among Egyptian Patients with Atopic and Nonatopic Bronchial Asthma. Immunol. Investig. 2016, 45, 87–106. [Google Scholar] [CrossRef]
- Yu, J.-I.; Han, W.-C.; Yun, K.-J.; Moon, H.-B.; Oh, G.-J.; Chae, S.-C. Identifying Polymorphisms in IL-31 and Their Association with Susceptibility to Asthma. Korean J. Pathol. 2012, 46, 162. [Google Scholar] [CrossRef]
- Bonanno, A.; Gangemi, S.; la Grutta, S.; Malizia, V.; Riccobono, L.; Colombo, P.; Cibella, F.; Profita, M. 25-Hydroxyvitamin D, IL-31, and IL-33 in Children with Allergic Disease of the Airways. Mediat. Inflamm. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Ip, W.K.; Wong, C.K.; Li, M.L.Y.; Li, P.W.; Cheung, P.F.Y.; Lam, C.W.K. Interleukin-31 Induces Cytokine and Chemokine Production from Human Bronchial Epithelial Cells through Activation of Mitogen-Activated Protein Kinase Signalling Pathways: Implications for the Allergic Response. Immunology 2007, 122, 532–541. [Google Scholar] [CrossRef]
- Chai, R.; Liu, B.; Qi, F. The Significance of the Levels of IL-4, IL-31 and TLSP in Patients with Asthma and/or Rhinitis. Immunotherapy 2017, 9, 331–337. [Google Scholar] [CrossRef]
- Ulambayar, B.; Lee, S.-H.; Yang, E.-M.; Ye, Y.-M.; Park, H.-S. Association Between Epithelial Cytokines and Clinical Phenotypes of Elderly Asthma. Allergy Asthma Immunol. Res. 2019, 11, 79. [Google Scholar] [CrossRef]
- Neuper, T.; Neureiter, D.; Sarajlic, M.; Strandt, H.; Bauer, R.; Schwarz, H.; Suchanek, P.; Korotchenko, E.; Dillon, S.R.; Hammerl, P.; et al. IL-31 Transgenic Mice Show Reduced Allergen-induced Lung Inflammation. Eur. J. Immunol. 2021, 51, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Perrigoue, J.G.; Li, J.; Zaph, C.; Goldschmidt, M.; Scott, P.; de Sauvage, F.J.; Pearce, E.J.; Ghilardi, N.; Artis, D. IL-31–IL-31R Interactions Negatively Regulate Type 2 Inflammation in the Lung. J. Exp. Med. 2007, 204, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Okano, M.; Fujiwara, T.; Higaki, T.; Makihara, S.; Haruna, T.; Noda, Y.; Kanai, K.; Kariya, S.; Yasueda, H.; Nishizaki, K. Characterization of Pollen Antigen–Induced IL-31 Production by PBMCs in Patients with Allergic Rhinitis. J. Allergy Clin. Immunol. 2011, 127, 277–279.e11. [Google Scholar] [CrossRef] [PubMed]
- Baumann, R.; Rabaszowski, M.; Stenin, I.; Gaertner-Akerboom, M.; Scheckenbach, K.; Wiltfang, J.; Schipper, J.; Wagenmann, M. The Release of IL-31 and IL-13 after Nasal Allergen Challenge and Their Relation to Nasal Symptoms. Clin. Transl. Allergy 2012, 2, 13. [Google Scholar] [CrossRef]
- Shah, S.A.; Ishinaga, H.; Hou, B.; Okano, M.; Takeuchi, K. Effects of Interleukin-31 on MUC5AC Gene Expression in Nasal Allergic Inflammation. Pharmacology 2013, 91, 158–164. [Google Scholar] [CrossRef]
- Liu, W.; Luo, R.; Chen, Y.; Sun, C.; Wang, J.; Zhou, L.; Li, Y.; Deng, L. Interleukin-31 Promotes Helper T Cell Type-2 Inflammation in Children with Allergic Rhinitis. Pediatric Res. 2015, 77, 20–28. [Google Scholar] [CrossRef]
- Saito, S.; Aoki, A.; Arai, I.; Takaishi, S.; Ito, H.; Akiyama, N.; Kiyonari, H. Regulation of Th2 Responses by Different Cell Types Expressing the Interleukin-31 Receptor. Allergy Asthma Clin. Immunol. 2017, 13, 23. [Google Scholar] [CrossRef]
- Qiao, Y.; Chen, J. Serum Levels of IL-31, IL-33 and ST2 in Allergic Rhinitis of Children in China. Cell. Mol. Biol. (Noisy-le-grand) 2018, 64, 52–55. [Google Scholar] [CrossRef]
- Auyeung, P.; Mittag, D.; Hodgkin, P.D.; Harrison, L.C. Autoreactive T Cells in Chronic Spontaneous Urticaria Target the IgE Fc Receptor Iα Subunit. J. Allergy Clin. Immunol. 2016, 138, 761–768.e4. [Google Scholar] [CrossRef]
- Raap, U.; Gehring, M.; Kleiner, S.; Rüdrich, U.; Eiz-Vesper, B.; Haas, H.; Kapp, A.; Gibbs, B.F. Human Basophils Are a Source of- and Are Differentially Activated by- IL-31. Clin. Exp. Allergy 2017, 47, 499–508. [Google Scholar] [CrossRef]
- Raap, U.; Wieczorek, D.; Gehring, M.; Pauls, I.; Ständer, S.; Kapp, A.; Wedi, B. Increased Levels of Serum IL-31 in Chronic Spontaneous Urticaria. Exp. Dermatol. 2010, 19, 464–466. [Google Scholar] [CrossRef]
- Lin, W.; Zhou, Q.; Liu, C.; Ying, M.; Xu, S. Increased Plasma IL-17, IL-31, and IL-33 Levels in Chronic Spontaneous Urticaria. Sci. Rep. 2017, 7, 17797. [Google Scholar] [CrossRef]
- Altrichter, S.; Hawro, T.; Hänel, K.; Czaja, K.; Lüscher, B.; Maurer, M.; Church, M.K.; Baron, J.M. Successful Omalizumab Treatment in Chronic Spontaneous Urticaria Is Associated with Lowering of Serum IL-31 Levels. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 454–455. [Google Scholar] [CrossRef]
- Rauber, M.M.; Pickert, J.; Holiangu, L.; Möbs, C.; Pfützner, W. Omalizumab Response Correlates with Reduced IFN -γ-, IL -10- and IL -31-secreting Cells in Chronic Spontaneous Urticaria. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 2078–2085. [Google Scholar] [CrossRef]
- Neis, M.; Peters, B.; Dreuw, A.; Wenzel, J.; Bieber, T.; Mauch, C.; Krieg, T.; Stanzel, S.; Heinrich, P.; Merk, H. Enhanced Expression Levels of IL-31 Correlate with IL-4 and IL-13 in Atopic and Allergic Contact Dermatitis. J. Allergy Clin. Immunol. 2006, 118, 930–937. [Google Scholar] [CrossRef]
- Takamori, A.; Nambu, A.; Sato, K.; Yamaguchi, S.; Matsuda, K.; Numata, T.; Sugawara, T.; Yoshizaki, T.; Arae, K.; Morita, H.; et al. IL-31 Is Crucial for Induction of Pruritus, but Not Inflammation, in Contact Hypersensitivity. Sci. Rep. 2018, 8, 6639. [Google Scholar] [CrossRef]
- Mesa, R.A.; Niblack, J.; Wadleigh, M.; Verstovsek, S.; Camoriano, J.; Barnes, S.; Tan, A.D.; Atherton, P.J.; Sloan, J.A.; Tefferi, A. The Burden of Fatigue and Quality of Life in Myeloproliferative Disorders (MPDs). Cancer 2007, 109, 68–76. [Google Scholar] [CrossRef]
- Diehn, F.; Tefferi, A. Pruritus in Polycythaemia Vera: Prevalence, Laboratory Correlates and Management. Br. J. Haematol. 2001, 115, 619–621. [Google Scholar] [CrossRef]
- Nattkemper, L.; Martinez-Escala, M.; Gelman, A.; Singer, E.; Rook, A.; Guitart, J.; Yosipovitch, G. Cutaneous T-Cell Lymphoma and Pruritus: The Expression of IL-31 and Its Receptors in the Skin. Acta Derm. Venereol. 2016, 96, 894–898. [Google Scholar] [CrossRef]
- Lange, M.; Gleń, J.; Zabłotna, M.; Nedoszytko, B.; Sokołowska-Wojdyło, M.; Rębała, K.; Ługowska-Umer, H.; Niedoszytko, M.; Górska, A.; Sikorska, M.; et al. Interleukin-31 Polymorphisms and Serum IL-31 Level in Patients with Mastocytosis: Correlation with Clinical Presentation and Pruritus. Acta Derm. Venereol. 2017, 97, 47–53. [Google Scholar] [CrossRef]
- Ohmatsu, H.; Sugaya, M.; Suga, H.; Morimura, S.; Miyagaki, T.; Kai, H.; Kagami, S.; Fujita, H.; Asano, Y.; Tada, Y.; et al. Serum IL-31 Levels Are Increased in Patients with Cutaneous T-Cell Lymphoma. Acta Derm. Venereol. 2012, 92, 282–283. [Google Scholar] [CrossRef] [PubMed]
- Miyagaki, T.; Sugaya, M.; Suga, H.; Ohmatsu, H.; Fujita, H.; Asano, Y.; Tada, Y.; Kadono, T.; Sato, S. Increased CCL18 Expression in Patients with Cutaneous T-Cell Lymphoma: Association with Disease Severity and Prognosis. J. Eur. Acad. Dermatol. Venereol. 2013, 27, e60–e67. [Google Scholar] [CrossRef] [PubMed]
- Singer, E.M.; Shin, D.B.; Nattkemper, L.A.; Benoit, B.M.; Klein, R.S.; Didigu, C.A.; Loren, A.W.; Dentchev, T.; Wysocka, M.; Yosipovitch, G.; et al. IL-31 Is Produced by the Malignant T-Cell Population in Cutaneous T-Cell Lymphoma and Correlates with CTCL Pruritus. J. Investig. Dermatol. 2013, 133, 2783–2785. [Google Scholar] [CrossRef] [PubMed]
- Malek, M.; Gleń, J.; Rębała, K.; Kowalczyk, A.; Sobjanek, M.; Nowicki, R.; Ruckemann-Dziurdzińska, K.; Sokołowska-Wojdyło, M. Il-31 Does Not Correlate to Pruritus Related to Early Stage Cutaneous T-Cell Lymphomas but Is Involved in Pathogenesis of the Disease. Acta Derm. Venereol. 2015, 95, 283–288. [Google Scholar] [CrossRef]
- Möbs, M.; Gryzik, S.; Haidar, A.; Humme, D.; Beyer, M.; Vandersee, S. Analysis of the IL-31 Pathway in Mycosis Fungoides and Sézary Syndrome. Arch. Dermatol. Res. 2015, 307, 479–485. [Google Scholar] [CrossRef]
- Abreu, M.; Miranda, M.; Castro, M.; Fernandes, I.; Cabral, R.; Santos, A.H.; Fonseca, S.; Rodrigues, J.; Leander, M.; Lau, C.; et al. IL-31 and IL-8 in Cutaneous T-Cell Lymphoma: Looking for Their Role in Itch. Adv. Hematol. 2021, 2021, 5582581. [Google Scholar] [CrossRef]
- Hartmann, K.; Wagner, N.; Rabenhorst, A.; Pflanz, L.; Leja, S.; Förster, A.; Gehring, M.; Kapp, A.; Raap, U. Serum IL-31 Levels Are Increased in a Subset of Patients with Mastocytosis and Correlate with Disease Severity in Adult Patients. J. Allergy Clin. Immunol. 2013, 132, 232–235.e4. [Google Scholar] [CrossRef]
- Musolino, C.; Allegra, A.; Mannucci, C.; Russo, S.; Alonci, A.; Maisano, V.; Calapai, G.; Gangemi, S. Possible Role of Interleukin-31/33 Axis in Imatinib Mesylate-Associated Skin Toxicity. Turk. J. Haematol. 2015, 32, 168–171. [Google Scholar] [CrossRef]
- Naumnik, W.; Naumnik, B.; Niewiarowska, K.; Ossolinska, M.; Chyczewska, E. Novel Cytokines: IL-27, IL-29, IL-31 and IL-33. Can They Be Useful in Clinical Practice at the Time Diagnosis of Lung Cancer? Exp. Oncol. 2012, 34, 348–353. [Google Scholar]
- Zeng, X.; Zhang, Z.; Gao, Q.-Q.; Wang, Y.-Y.; Yu, X.-Z.; Zhou, B.; Xi, M.-R. Clinical Significance of Serum Interleukin-31 and Interleukin-33 Levels in Patients of Endometrial Cancer: A Case Control Study. Dis. Markers 2016, 2016, 9262919. [Google Scholar] [CrossRef]
- Zeng, X.; Li, J.; Kang, L.-N.; Xi, M.-R.; Liao, G.-D. Potential Clinical Value of Interleukin-31 and Interleukin-33 with Their Receptors Expression as Diagnostic and Predictive Factors in Endometrial Cancer: A Case-Control Study. Int. J. Clin. Exp. Pathol. 2020, 13, 1324–1332. [Google Scholar]
- Kan, T.; Feldman, E.; Timaner, M.; Raviv, Z.; Shen-Orr, S.; Aronheim, A.; Shaked, Y. IL-31 Induces Antitumor Immunity in Breast Carcinoma. J. ImmunoTherapy Cancer 2020, 8, e001010. [Google Scholar] [CrossRef]
- Wang, X.; Lin, F.; Li, J.; Wang, H. A Comprehensive Risk Assessment Model for Ovarian Cancer Patients with Phospho-STAT3 and IL-31 as Immune Infiltration Relevant Genes. OncoTargets Ther. 2020, 13, 5617–5628. [Google Scholar] [CrossRef]
- He, Y.; Zhang, X.; Pan, W.; Tai, F.; Liang, L.; Shi, J. Interleukin-31 Receptor α Is Required for Basal-Like Breast Cancer Progression. Front. Oncol. 2020, 10, 816. [Google Scholar] [CrossRef]
- Liu, C.; Wang, Y.; Song, H.; Li, Q.; Zhang, Y.; Chen, P.; Song, Y.; Su, M.; Huang, Q.; Wang, M.; et al. Genetic Association of Interleukin-31 Gene Polymorphisms with Epithelial Ovarian Cancer in Chinese Population. Dis. Markers 2018, 2018, 3503858. [Google Scholar] [CrossRef]
- Lan, Z.; Wang, Y.; Yu, X.; Song, H.; Li, Q.; You, D.; Yuan, M.; Zeng, X.; Zhou, B.; Song, Y.; et al. Interleukin-31 Single Nucleotide Polymorphisms Are Significantly Associated with Endometrial Cancer in Chinese Han Women. Int. J. Clin. Exp. Pathol. 2018, 11, 894–903. [Google Scholar]
- Li, Q.; Tang, T.; Zhang, P.; Liu, C.; Pu, Y.; Zhang, Y.; Song, H.; Wang, Y.; Song, Y.; Su, M.; et al. Correlation of IL-31 Gene Polymorphisms with Susceptibility and Clinical Recurrence of Bladder Cancer. Fam. Cancer 2018, 17, 577–585. [Google Scholar] [CrossRef]
- Hashimoto, T.; Satoh, T.; Yokozeki, H. Pruritus in Ordinary Scabies: IL-31 from Macrophages Induced by Overexpression of Thymic Stromal Lymphopoietin and Periostin. Allergy 2019, 74, 1727–1737. [Google Scholar] [CrossRef]
- Gu, X.; Wei, C.; Zhu, X.; Lu, F.; Sheng, B.; Zang, X. Effect of Interleukin-31 on Septic Shock through Regulating Inflammasomes and Interleukin-1β. Exp. Ther. Med. 2018, 16, 171–177. [Google Scholar] [CrossRef]
- Watany, M.M.; Elmazny, M.I.; Nasif, E.M. Interleukin-31 Interaction with Inflammasome: A Promising Diagnostic and Prognostic Panel for Early Sepsis Identification in Critically Ill Patients. Cytokine 2020, 131, 155102. [Google Scholar] [CrossRef]
- Ming, D.; Yu, X.; Guo, R.; Deng, Y.; Li, J.; Lin, C.; Su, M.; Lin, Z.; Su, Z. Elevated TGF-Β1/IL-31 Pathway Is Associated with the Disease Severity of Hepatitis B Virus–Related Liver Cirrhosis. Viral Immunol. 2015, 28, 209–216. [Google Scholar] [CrossRef]
- Yu, X.; Guo, R.; Ming, D.; Deng, Y.; Su, M.; Lin, C.; Li, J.; Lin, Z.; Su, Z. The Transforming Growth Factor Β1/Interleukin-31 Pathway Is Upregulated in Patients with Hepatitis B Virus-Related Acute-on-Chronic Liver Failure and Is Associated with Disease Severity and Survival. Clin. Vaccine Immunol. 2015, 22, 484–492. [Google Scholar] [CrossRef]
- Perrigoue, J.G.; Zaph, C.; Guild, K.; Du, Y.; Artis, D. IL-31-IL-31R Interactions Limit the Magnitude of Th2 Cytokine-Dependent Immunity and Inflammation Following Intestinal Helminth Infection. J. Immunol. 2009, 182, 6088–6094. [Google Scholar] [CrossRef]
- Huang, X.; Grüner, B.; Lechner, C.J.; Kern, P.; Soboslay, P.T. Distinctive Cytokine, Chemokine, and Antibody Responses in Echinococcus Multilocularis-Infected Patients with Cured, Stable, or Progressive Disease. Med. Microbiol. Immunol. 2014, 203, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Kasutani, K.; Fujii, E.; Ohyama, S.; Adachi, H.; Hasegawa, M.; Kitamura, H.; Yamashita, N. Anti-IL-31 Receptor Antibody Is Shown to Be a Potential Therapeutic Option for Treating Itch and Dermatitis in Mice. Br. J. Pharmacol. 2014, 171, 5049–5058. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, A.J.; Fleck, T.J.; Humphrey, W.R.; Galvan, B.A.; Aleo, M.M.; Mahabir, S.P.; Tena, J.; Greenwood, K.G.; McCall, R.B. IL-31-induced Pruritus in Dogs: A Novel Experimental Model to Evaluate Anti-pruritic Effects of Canine Therapeutics. Vet. Dermatol. 2016, 27, 34-e10. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.P.; Rosychuk, R.A.W.; Contreras, E.T.; Schissler, J.R.; Simpson, A.C. A Retrospective Analysis of the Use of Lokivetmab in the Management of Allergic Pruritus in a Referral Population of 135 Dogs in the Western USA. Vet. Dermatol. 2018, 29, 489-e164. [Google Scholar] [CrossRef] [PubMed]
- Silverberg, J.I.; Pinter, A.; Pulka, G.; Poulin, Y.; Bouaziz, J.-D.; Wollenberg, A.; Murrell, D.F.; Alexis, A.; Lindsey, L.; Ahmad, F.; et al. Phase 2B Randomized Study of Nemolizumab in Adults with Moderate-to-Severe Atopic Dermatitis and Severe Pruritus. J. Allergy Clin. Immunol. 2020, 145, 173–182. [Google Scholar] [CrossRef]
- Kabashima, K.; Furue, M.; Hanifin, J.M.; Pulka, G.; Wollenberg, A.; Galus, R.; Etoh, T.; Mihara, R.; Nakano, M.; Ruzicka, T. Nemolizumab in Patients with Moderate-to-Severe Atopic Dermatitis: Randomized, Phase II, Long-Term Extension Study. J. Allergy Clin. Immunol. 2018, 142, 1121–1130.e7. [Google Scholar] [CrossRef]
- Oweis, A.O.; AL-Qarqaz, F.; Bodoor, K.; Heis, L.; Alfaqih, M.A.; Almomani, R.; Obeidat, M.A.; Alshelleh, S.A. Elevated Interleukin 31 Serum Levels in Hemodialysis Patients Are Associated with Uremic Pruritus. Cytokine 2021, 138, 155369. [Google Scholar] [CrossRef]
- Hashimoto, T.; Sakai, K.; Sanders, K.; Yosipovitch, G.; Akiyama, T. Antipruritic Effects of Janus Kinase Inhibitor Tofacitinib in a Mouse Model of Psoriasis. Acta Derm. Venereol. 2019, 99, 298–303. [Google Scholar] [CrossRef]
- Nemoto, O.; Furue, M.; Nakagawa, H.; Shiramoto, M.; Hanada, R.; Matsuki, S.; Imayama, S.; Kato, M.; Hasebe, I.; Taira, K.; et al. The First Trial of CIM331, a Humanized Antihuman Interleukin-31 Receptor A Antibody, in Healthy Volunteers and Patients with Atopic Dermatitis to Evaluate Safety, Tolerability and Pharmacokinetics of a Single Dose in a Randomized, Double-Blind, Placebo-Co. Br. J. Dermatol. 2016, 174, 296–304. [Google Scholar] [CrossRef]
- Cedeno-Laurent, F.; Singer, E.M.; Wysocka, M.; Benoit, B.M.; Vittorio, C.C.; Kim, E.J.; Yosipovitch, G.; Rook, A.H. Improved Pruritus Correlates with Lower Levels of IL-31 in CTCL Patients under Different Therapeutic Modalities. Clin. Immunol. 2015, 158, 1–7. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Author/Citation | Target Cells | Mediators/Pathway | Effects |
|---|---|---|---|---|
| AD | Wong et al. [16] | Eosinophils—fibroblasts interaction | IL-33, IL-6, CXCL1, CXCL10, CCL2, CCL5 | Skin inflammation |
| Kasraie et al. [26] | macrophages | STAT-1 and 5 | Itch | |
| Tang et al. [31] | keratinocytes | Histamine, TSLP, substance P | Itch, changes in skin pH, trans-epidermal water loss | |
| PSO | Nattkemper et al. [42] | - | TRP melastatin 8, TRP vanilloid 3, phospholipase C, IL-36α/γ | Itch |
| CHRONIC ITCH | Arai et al. [47] | Dorsal-root ganglia | - | Perpetration of the itching stimulus |
| Andoh et al. [50] | Mouse keratinocytes | LTB4 | - | |
| ASTHMA | Moaaz et al. [55] | Bronchial epithelium | MAPK, EGF, VEGF, CCL2 | Bronchial inflammation |
| Neuper et al. [61] | Bronchial epithelium of IL-31RA-/-mice | - | Lower-leukocyte infiltration, reduced mucus and epithelial thickening | |
| AR | Baumann et al. [64] | Nasal epithelium | CCL17, CCL22, CCL1 | Inflammation in the nasal epithelium |
| Liu et al. [66] | NECs PBMCs | MUC5AC, ECP IL-4, IL-5, IL-13 | Enhanced Th2 response and eosinophils activation | |
| CTCL | Singer et al. [83] | PBMCs | - | Itch |
| LUNG CANCER | Naumnik et al. [89] | Bronchial and alveolar epithelial cells, pulmonary fibroblasts, and macrophages | EGF, VEGF | Increased cancer progression |
| MURINE BREAST CANCER | Kan et al. [92] | CTLs | Decreased levels of IL-10 | Increase of total CTLs, antitumor activity |
| OVARIAN and BREAST CANCER | Wang et al. [93] He et al. [94] | - - | JNK-STAT - | Tumor invasion and metastasis |
| SEPSIS | Watany et al. [100] | - | Down-regulation of NLRP3, NLRP1 | Pyroptosis, release of IL-1beta, IL-18 and Gasdermin-D, decrease in TNFalpha |
| HBV | Ming et al., Yu et al., [101,102] | - | TGF- β1 | Liver fibrosis, progression to cirrhosis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borgia, F.; Custurone, P.; Li Pomi, F.; Cordiano, R.; Alessandrello, C.; Gangemi, S. IL-31: State of the Art for an Inflammation-Oriented Interleukin. Int. J. Mol. Sci. 2022, 23, 6507. https://doi.org/10.3390/ijms23126507
Borgia F, Custurone P, Li Pomi F, Cordiano R, Alessandrello C, Gangemi S. IL-31: State of the Art for an Inflammation-Oriented Interleukin. International Journal of Molecular Sciences. 2022; 23(12):6507. https://doi.org/10.3390/ijms23126507
Chicago/Turabian StyleBorgia, Francesco, Paolo Custurone, Federica Li Pomi, Raffaele Cordiano, Clara Alessandrello, and Sebastiano Gangemi. 2022. "IL-31: State of the Art for an Inflammation-Oriented Interleukin" International Journal of Molecular Sciences 23, no. 12: 6507. https://doi.org/10.3390/ijms23126507
APA StyleBorgia, F., Custurone, P., Li Pomi, F., Cordiano, R., Alessandrello, C., & Gangemi, S. (2022). IL-31: State of the Art for an Inflammation-Oriented Interleukin. International Journal of Molecular Sciences, 23(12), 6507. https://doi.org/10.3390/ijms23126507