
Diet-Induced Hypercholesterolemia Leads to Cardiac Dysfunction and Alterations in the Myocardial Proteome

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Eight Weeks of Cholesterol-Enriched Diet in Rats Resulted in Elevated Plasma Lipid Levels
2.2. Cholesterol-Enriched-Diet-Induced Diastolic Dysfunction in the Heart
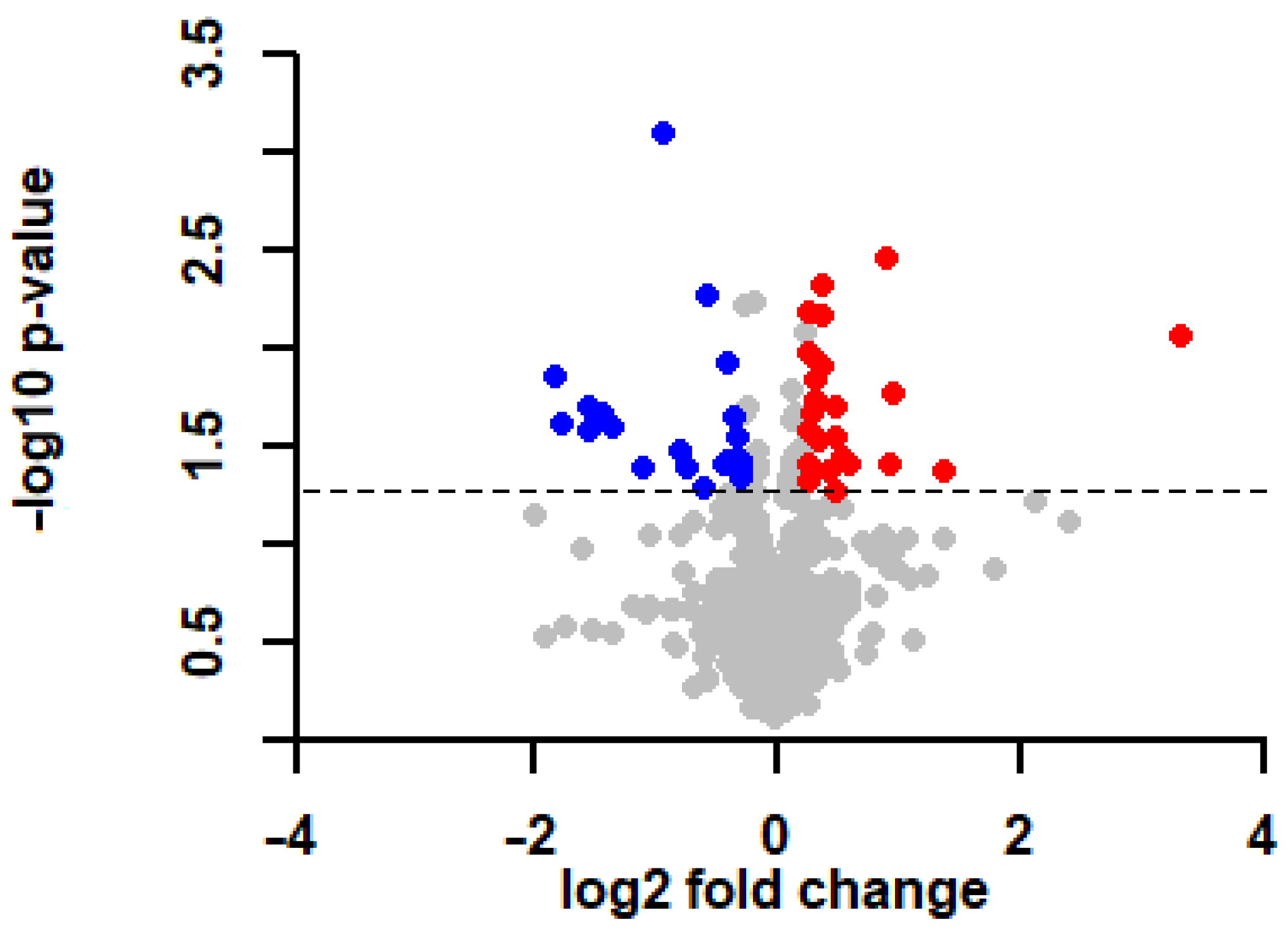
2.3. General Proteomic Characterization of the Left Ventricle of Hypercholesterolemic Rats
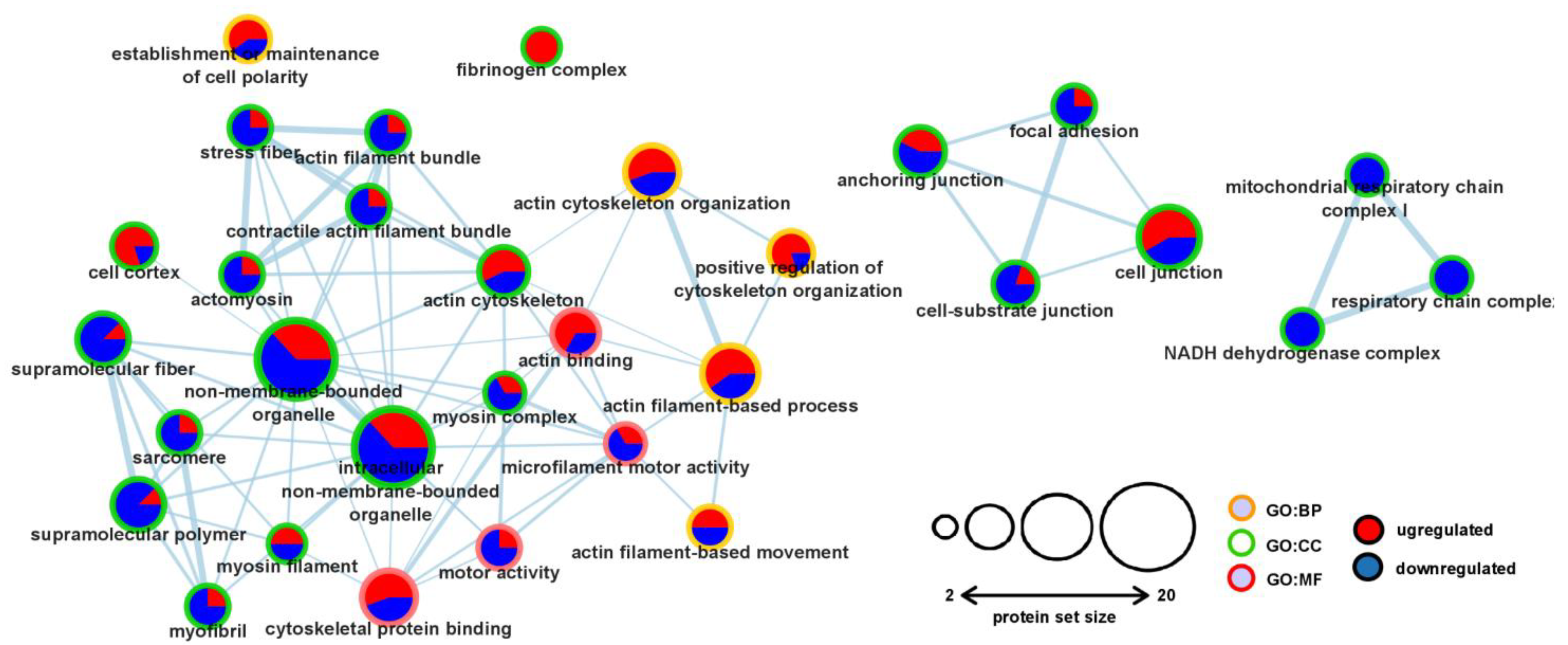
2.4. Pathway Enrichment Analysis of the Significantly Altered Proteins Revealed Changes in the Contractile and Cytoskeletal Systems
2.5. Functional Interaction Analysis of the Differentially Expressed Proteins
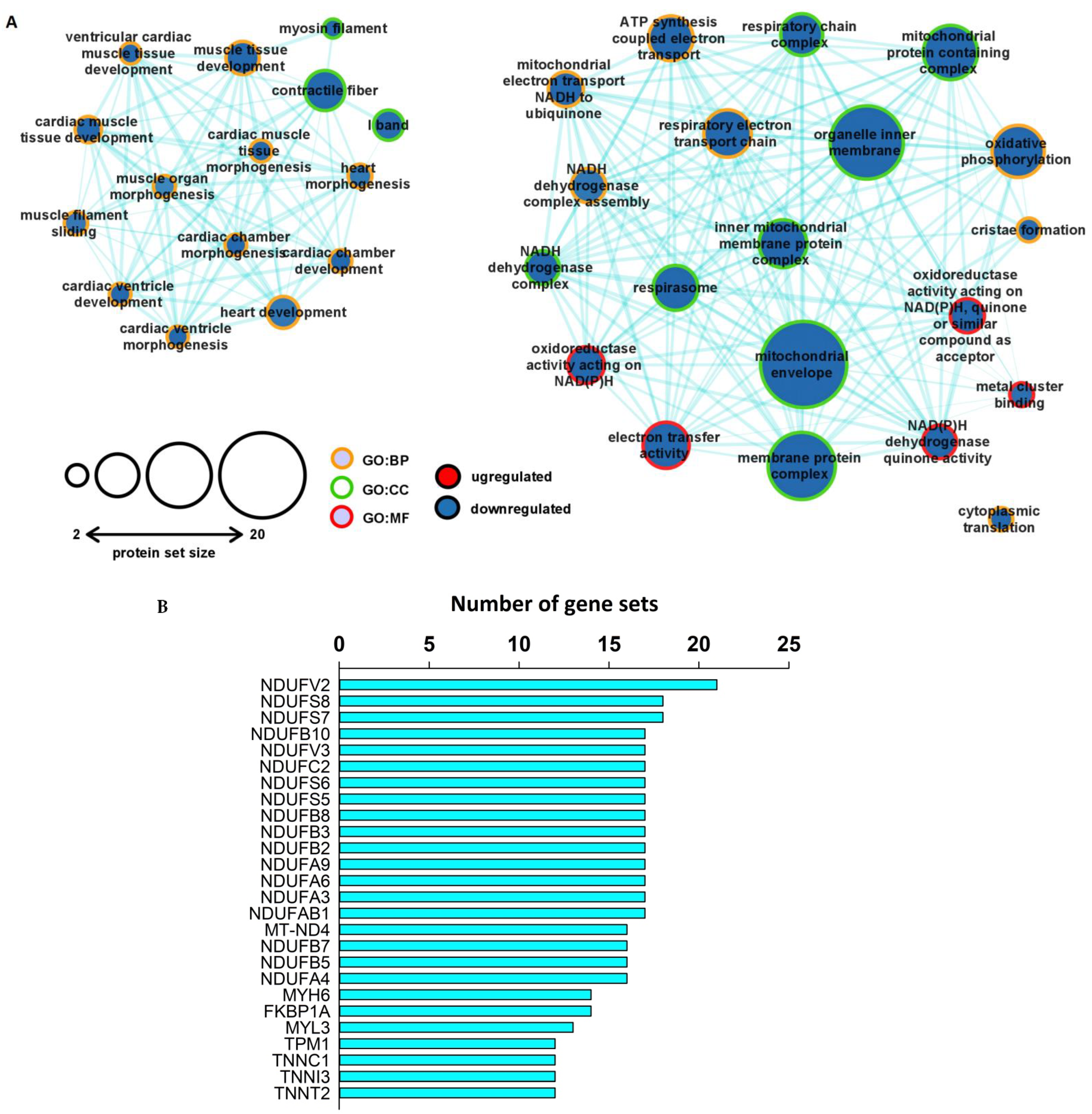
2.6. Protein-Specific Gene Set Enrichment Analysis Revealed Downregulated Expression Patterns of Mitochondrial and Contractile Proteins in the Unfiltered, Whole Left Ventricular Proteome
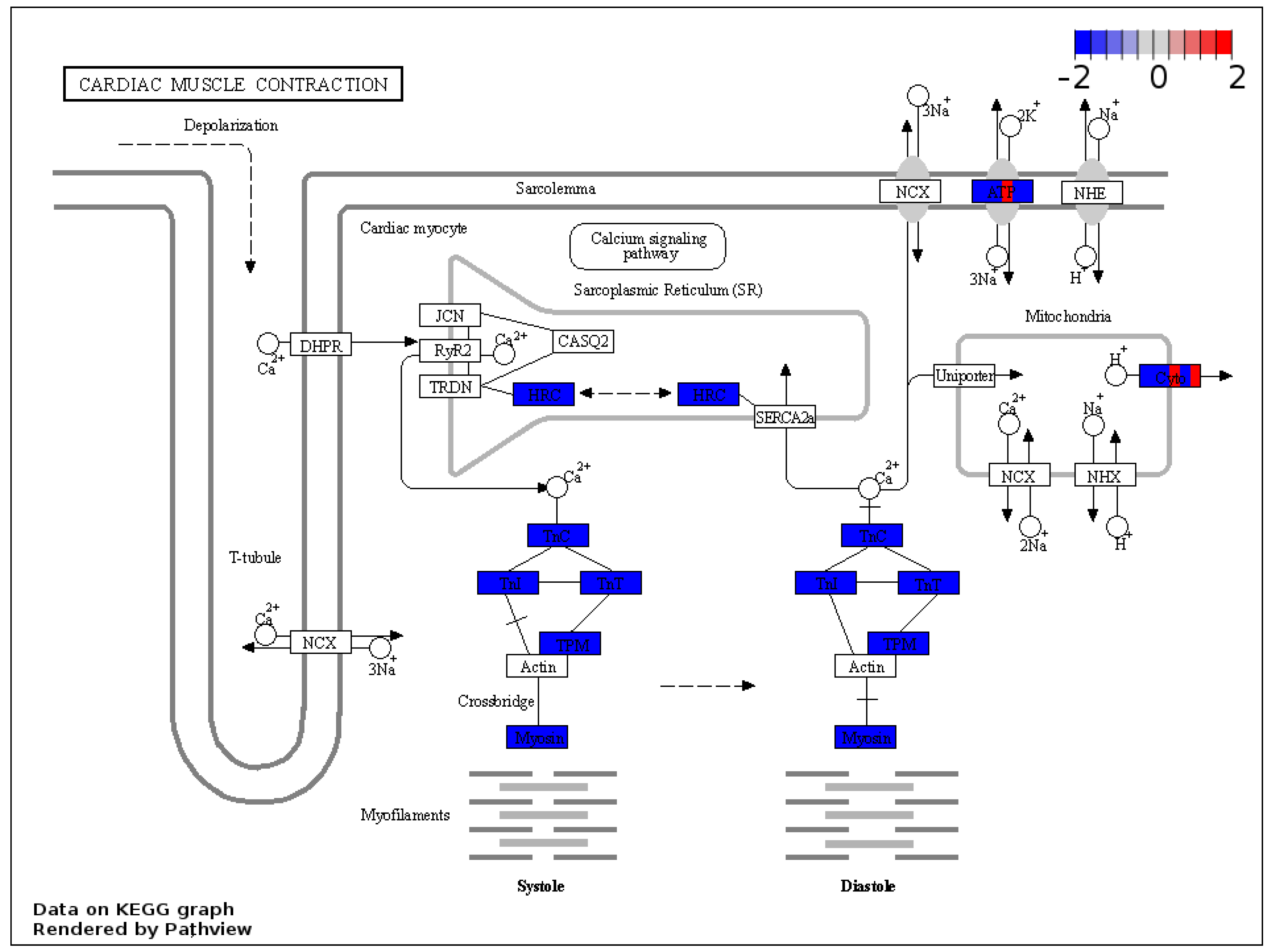
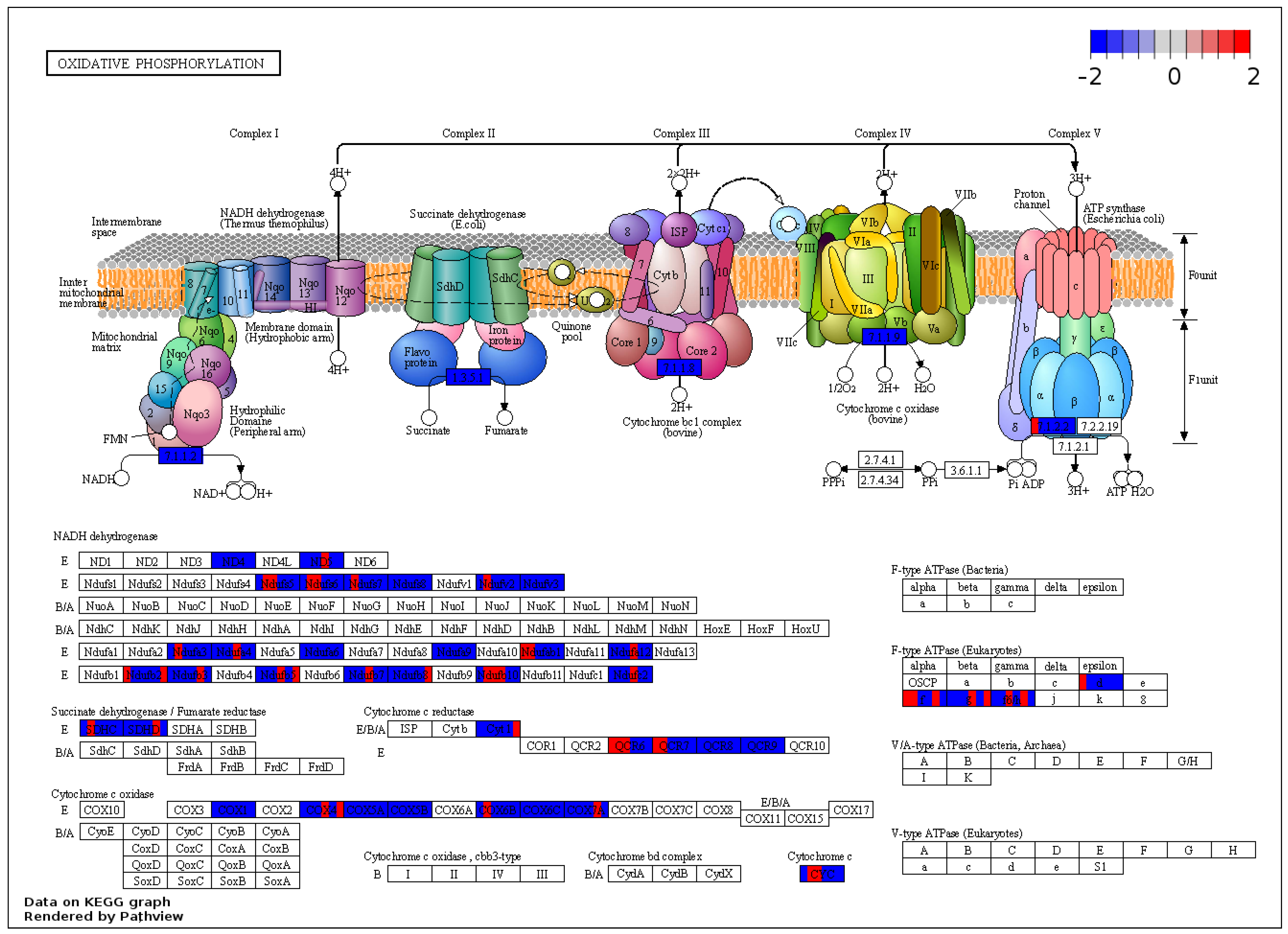
2.7. KEGG Analysis of the Output of GSEA
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Setup
4.3. Plasma Lipid Measurement
4.4. Transthoracic Echocardiography
4.5. Protein Extraction
4.6. Protein Digestion
4.7. LC-MS Analysis
4.8. Proteomic Data Analysis
4.9. Pathway Enrichment Analysis
4.10. Functional Protein–Protein Interaction and Network Clustering Analysis
4.11. Gene Set Enrichment Analysis of the Proteomic Dataset
4.12. KEGG Analysis and Visualization of Core Enrichment Proteins with Pathview
4.13. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Timmis, A.; Townsend, N.; Gale, C.P.; Torbica, A.; Lettino, M.; Petersen, S.E.; Mossialos, E.A.; Maggioni, A.P.; Kazakiewicz, D.; May, H.T.; et al. European Society of Cardiology: Cardiovascular Disease Statistics 2019. Eur. Heart J. 2020, 41, 12–85. [Google Scholar] [CrossRef] [PubMed]
- Gidding, S.S.; Allen, N.B. Cholesterol and Atherosclerotic Cardiovascular Disease: A Lifelong Problem. J. Am. Heart Assoc. 2019, 8, e012924. [Google Scholar] [CrossRef] [PubMed]
- Csont, T.; Bereczki, E.; Bencsik, P.; Fodor, G.; Görbe, A.; Zvara, A.; Csonka, C.; Puskás, L.G.; Sántha, M.; Ferdinandy, P. Hypercholesterolemia increases myocardial oxidative and nitrosative stress thereby leading to cardiac dysfunction in apoB-100 transgenic mice. Cardiovasc. Res. 2007, 76, 100–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varga, Z.V.; Kupai, K.; Szűcs, G.; Gáspár, R.; Pálóczi, J.; Faragó, N.; Zvara, A.; Puskás, L.G.; Rázga, Z.; Tiszlavicz, L.; et al. MicroRNA-25-dependent up-regulation of NADPH oxidase 4 (NOX4) mediates hypercholesterolemia-induced oxidative/nitrative stress and subsequent dysfunction in the heart. J. Mol. Cell. Cardiol. 2013, 62, 111–121. [Google Scholar] [CrossRef]
- Pluijmert, N.J.; den Haan, M.C.; van Zuylen, V.L.; Steendijk, P.; de Boer, H.C.; van Zonneveld, A.J.; Fibbe, W.E.; Schalij, M.J.; Quax, P.H.A.; Atsma, D.E. Hypercholesterolemia affects cardiac function, infarct size and inflammation in APOE*3-Leiden mice following myocardial ischemia-reperfusion injury. PLoS ONE 2019, 14, e0217582. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.S.; Li, T.D.; Zeng, Z.H. Mechanisms underlying direct actions of hyperlipidemia on myocardium: An updated review. Lipids Health Dis. 2020, 19, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Csonka, C.; Sárközy, M.; Pipicz, M.; Dux, L.; Csont, T. Modulation of Hypercholesterolemia-Induced Oxidative/Nitrative Stress in the Heart. Oxid. Med. Cell. Longev. 2016, 2016, 3863726. [Google Scholar] [CrossRef]
- Csonka, C.; Murlasits, Z.; Ferdinandy, P.; Csont, T. Ischemic stress adaptation of the myocardium in the disease states: Role of hyperlipidemia. In Advances in Cardiomyocyte Research; Nánási, P., Ed.; Transworld Research Network: Kerala, India, 2009; pp. 245–265. [Google Scholar]
- McCommis, K.S.; McGee, A.M.; Laughlin, M.H.; Bowles, D.K.; Baines, C.P. Hypercholesterolemia increases mitochondrial oxidative stress and enhances the MPT response in the porcine myocardium: Beneficial effects of chronic exercise. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2011, 301, R1250–R1258. [Google Scholar] [CrossRef] [Green Version]
- Csonka, C.; Kupai, K.; Bencsik, P.; Görbe, A.; Pálóczi, J.; Zvara, A.; Puskás, L.G.; Csont, T.; Ferdinandy, P. Cholesterol-enriched diet inhibits cardioprotection by ATP-sensitive K+ channel activators cromakalim and diazoxide. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H405–H413. [Google Scholar] [CrossRef] [Green Version]
- Puskás, L.G.; Nagy, Z.B.; Giricz, Z.; Onody, A.; Csonka, C.; Kitajka, K.; Hackler, L., Jr.; Zvara, A.; Ferdinandy, P. Cholesterol diet-induced hyperlipidemia influences gene expression pattern of rat hearts: A DNA microarray study. FEBS Lett. 2004, 562, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Kocsis, G.F.; Csont, T.; Varga-Orvos, Z.; Puskas, L.G.; Murlasits, Z.; Ferdinandy, P. Expression of genes related to oxidative/nitrosative stress in mouse hearts: Effect of preconditioning and cholesterol diet. Med. Sci. Monit. 2010, 16, BR32–BR39. [Google Scholar] [PubMed]
- Sárközy, M.; Zvara, Á.; Gyémánt, N.; Fekete, V.; Kocsis, G.F.; Pipis, J.; Szűcs, G.; Csonka, C.; Puskás, L.G.; Ferdinandy, P.; et al. Metabolic syndrome influences cardiac gene expression pattern at the transcript level in male ZDF rats. Cardiovasc. Diabetol. 2013, 12, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Walker, K.E.; Hanley, F.; Narula, J.; Houser, S.R.; Tulenko, T.N. Cardiac systolic and diastolic dysfunction after a cholesterol-rich diet. Circulation 2004, 109, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Mu, Y.; Han, W.; Wang, C. Association of hypercholesterolemia and cardiac function evaluated by speckle tracking echocardiography in a rabbit model. Lipids Health Dis. 2014, 13, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, K.; Ciotola, M.; Sasso, F.C.; Cozzolino, D.; Saccomanno, F.; Assaloni, R.; Ceriello, A.; Giugliano, D. Effect of a single high-fat meal on endothelial function in patients with the metabolic syndrome: Role of tumor necrosis factor-α. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 274–279. [Google Scholar] [CrossRef]
- Talini, E.; Di Bello, V.; Bianchi, C.; Palagi, C.; Delle Donne, M.G.; Penno, G.; Nardi, C.; Canale, M.L.; Del Prato, S.; Mariani, M.; et al. Early impairment of left ventricular function in hypercholesterolemia and its reversibility after short term treatment with rosuvastatin A preliminary echocardiographic study. Atherosclerosis 2008, 197, 346–354. [Google Scholar] [CrossRef]
- Saracoglu, E.; Kılıç, S.; Vuruşkan, E.; Düzen, I.; Çekici, Y.; Kuzu, Z.; Yıldırım, A.; Küçükosmanoğlu, M.; Çetin, M. Prediction of subtle left ventricular systolic dysfunction in homozygous and heterozygous familial hypercholesterolemia: Genetic analyses and speckle tracking echocardiography study. Echocardiography 2018, 35, 1289–1299. [Google Scholar] [CrossRef]
- Muthuramu, I.; Mishra, M.; Aboumsallem, J.P.; Postnov, A.; Gheysens, O.; De Geest, B. Cholesterol lowering attenuates pressure overload-induced heart failure in mice with mild hypercholesterolemia. Aging 2019, 11, 6872–6891. [Google Scholar] [CrossRef]
- Rubinstein, J.; Pelosi, A.; Vedre, A.; Kotaru, P.; Abela, G.S. Hypercholesterolemia and myocardial function evaluated via tissue doppler imaging. Cardiovasc. Ultrasound 2009, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Nagueh, S.F.; Smiseth, O.A.; Appleton, C.P.; Byrd, B.F., 3rd; Dokainish, H.; Edvardsen, T.; Flachskampf, F.A.; Gillebert, T.C.; Klein, A.L.; Lancellotti, P.; et al. Recommendations for the Evaluation of Left Ventricular Diastolic Function by Echocardiography: An Update from the American Society of Echocardiography and the European Association of Cardiovascular Imaging. J. Am. Soc. Echocardiogr. Off. Publ. Am. Soc. Echocardiogr. 2016, 29, 277–314. [Google Scholar] [CrossRef] [Green Version]
- Demján, V.; Kiss, T.; Siska, A.; Szabó, M.R.; Sárközy, M.; Földesi, I.; Csupor, D.; Csont, T. Effect of Stellaria media Tea on Lipid Profile in Rats. Evid.-Based Complement. Altern. Med. Ecam 2020, 2020, 5109328. [Google Scholar] [CrossRef] [Green Version]
- Hołda, M.K.; Stachowicz, A.; Suski, M.; Wojtysiak, D.; Sowińska, N.; Arent, Z.; Palka, N.; Podolec, P.; Kopeć, G. Myocardial proteomic profile in pulmonary arterial hypertension. Sci. Rep. 2020, 10, 14351. [Google Scholar] [CrossRef]
- Szűcs, G.; Sója, A.; Péter, M.; Sárközy, M.; Bruszel, B.; Siska, A.; Földesi, I.; Szabó, Z.; Janáky, T.; Vígh, L.; et al. Prediabetes Induced by Fructose-Enriched Diet Influences Cardiac Lipidome and Proteome and Leads to Deterioration of Cardiac Function prior to the Development of Excessive Oxidative Stress and Cell Damage. Oxid. Med. Cell. Longev. 2019, 2019, 3218275. [Google Scholar] [CrossRef]
- Yang, M.; Yan, J.; Wu, A.; Zhao, W.; Qin, J.; Pogwizd, S.M.; Wu, X.; Yuan, S.; Ai, X. Alterations of housekeeping proteins in human aged and diseased hearts. Pflügers Arch.-Eur. J. Physiol. 2021, 473, 351–362. [Google Scholar] [CrossRef]
- Michaelson, J.; Hariharan, V.; Huang, H. Hyperglycemic and Hyperlipidemic Conditions Alter Cardiac Cell Biomechanical Properties. Biophys. J. 2014, 106, 2322–2329. [Google Scholar] [CrossRef] [Green Version]
- Varela, R.; Rauschert, I.; Romanelli, G.; Alberro, A.; Benech, J.C. Hyperglycemia and hyperlipidemia can induce morphophysiological changes in rat cardiac cell line. Biochem. Biophys. Rep. 2021, 26, 100983. [Google Scholar] [CrossRef]
- Ojamaa, K.; Kenessey, A.; Shenoy, R.; Klein, I. Thyroid hormone metabolism and cardiac gene expression after acute myocardial infarction in the rat. Am. J. Physiol.-Endocrinol. Metab. 2000, 279, E1319–E1324. [Google Scholar] [CrossRef]
- Razeghi, P.; Young, M.E.; Alcorn, J.L.; Moravec, C.S.; Frazier, O.H.; Taegtmeyer, H. Metabolic gene expression in fetal and failing human heart. Circulation 2001, 104, 2923–2931. [Google Scholar] [CrossRef] [Green Version]
- Nakao, K.; Minobe, W.; Roden, R.; Bristow, M.R.; Leinwand, L.A. Myosin heavy chain gene expression in human heart failure. J. Clin. Investig. 1997, 100, 2362–2370. [Google Scholar] [CrossRef]
- Krenz, M.; Robbins, J. Impact of beta-myosin heavy chain expression on cardiac function during stress. J. Am. Coll. Cardiol. 2004, 44, 2390–2397. [Google Scholar] [CrossRef] [Green Version]
- Herron, T.J.; McDonald, K.S. Small amounts of alpha-myosin heavy chain isoform expression significantly increase power output of rat cardiac myocyte fragments. Circ Res. 2002, 90, 1150–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sequeira, V.; Nijenkamp, L.L.A.M.; Regan, J.A.; van der Velden, J. The physiological role of cardiac cytoskeleton and its alterations in heart failure. Biochim. Biophys. Acta (BBA)-Biomembr. 2014, 1838, 700–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehler, E. Actin-associated proteins and cardiomyopathy-the ’unknown’ beyond troponin and tropomyosin. Biophys. Rev. 2018, 10, 1121–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Tian, R. Mitochondrial dysfunction in pathophysiology of heart failure. J. Clin. Investig. 2018, 128, 3716–3726. [Google Scholar] [CrossRef] [Green Version]
- Bhatti, J.S.; Bhatti, G.K.; Reddy, P.H. Mitochondrial dysfunction and oxidative stress in metabolic disorders—A step towards mitochondria based therapeutic strategies. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 1066–1077. [Google Scholar] [CrossRef]
- Castillo, R.L.; Herrera, E.A.; Gonzalez-Candia, A.; Reyes-Farias, M.; de la Jara, N.; Peña, J.P.; Carrasco-Pozo, C. Quercetin Prevents Diastolic Dysfunction Induced by a High-Cholesterol Diet: Role of Oxidative Stress and Bioenergetics in Hyperglycemic Rats. Oxid. Med. Cell. Longev. 2018, 2018, 7239123. [Google Scholar] [CrossRef]
- Elustondo, P.; Martin, L.A.; Karten, B. Mitochondrial cholesterol import. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2017, 1862, 90–101. [Google Scholar] [CrossRef]
- Szabó, M.R.; Gáspár, R.; Pipicz, M.; Zsindely, N.; Diószegi, P.; Sárközy, M.; Bodai, L.; Csont, T. Hypercholesterolemia Interferes with Induction of miR-125b-1-3p in Preconditioned Hearts. Int. J. Mol. Sci. 2020, 21, 3744. [Google Scholar] [CrossRef]
- Pipicz, M.; Varga, Z.V.; Kupai, K.; Gáspár, R.; Kocsis, G.F.; Csonka, C.; Csont, T. Rapid ventricular pacing-induced postconditioning attenuates reperfusion injury: Effects on peroxynitrite, RISK and SAFE pathways. Br. J. Pharmacol. 2015, 172, 3472–3483. [Google Scholar] [CrossRef] [Green Version]
- Searle, B.C.; Pino, L.K.; Egertson, J.D.; Ting, Y.S.; Lawrence, R.T.; MacLean, B.X.; Villén, J.; MacCoss, M.J. Chromatogram libraries improve peptide detection and quantification by data independent acquisition mass spectrometry. Nat. Commun. 2018, 9, 5128. [Google Scholar] [CrossRef] [Green Version]
- Gessulat, S.; Schmidt, T.; Zolg, D.P.; Samaras, P.; Schnatbaum, K.; Zerweck, J.; Knaute, T.; Rechenberger, J.; Delanghe, B.; Huhmer, A.; et al. Prosit: Proteome-wide prediction of peptide tandem mass spectra by deep learning. Nat. Methods 2019, 16, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway enrichment analysis and visualization of omics data using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Doncheva, N.T.; Morris, J.H.; Gorodkin, J.; Jensen, L.J. Cytoscape StringApp: Network Analysis and Visualization of Proteomics Data. J. Proteome Res. 2019, 18, 623–632. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdóttir, H.; Tamayo, P.; Mesirov, J.P. Molecular signatures database (MSigDB) 3.0. Bioinformatics 2011, 27, 1739–1740. [Google Scholar] [CrossRef]
- Luo, W.; Brouwer, C. Pathview: An R/Bioconductor package for pathway-based data integration and visualization. Bioinformatics 2013, 29, 1830–1831. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Pant, G.; Bhavnasi, Y.K.; Blanchard, S.G., Jr.; Brouwer, C. Pathview Web: User friendly pathway visualization and data integration. Nucleic Acids Res. 2017, 45, W501–W508. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normochol | Hyperchol | |
|---|---|---|
| Body weight (g) | 485 ± 22 | 521 ± 17 |
| Tibia length (cm) | 4.20 ± 0.08 | 4.21 ± 0.05 |
| Left ventricular weight (mg) | 1242 ± 42 | 1230 ± 48 |
| Total cholesterol (mmol/L) | 1.52 ± 0.11 | 4.35 ± 0.21 * |
| Total triglyceride (mmol/L) | 0.44 ± 0.03 | 1.18 ± 0.08 * |
| Normochol | Hyperchol | p-Value | |
|---|---|---|---|
| AWTs (mm) | 3.86 ± 0.01 | 3.74 ± 0.13 | 0.471 |
| AWTd (mm) | 2.03 ± 0.10 | 2.17 ± 0.13 | 0.420 |
| IWTs (mm) | 3.88 ± 0.12 | 3.62 ± 0.13 | 0.154 |
| IWTd (mm) | 2.39 ± 0.13 | 2.30 ± 0.14 | 0.660 |
| PWTs (mm) | 3.71 ± 0.09 | 3.70 ± 0.22 | 0.938 |
| PWTd (mm) | 2.46 ± 0.20 | 2.43 ± 0.13 | 0.902 |
| SWTs (mm) | 3.81 ± 0.05 | 3.69 ± 0.13 | 0.442 |
| SWTd (mm) | 2.18 ± 0.06 | 2.27 ± 0.13 | 0.548 |
| LVESD (mm) | 2.39 ± 0.10 | 2.85 ± 0.28 | 0.155 |
| LVEDD (mm) | 6.32 ± 0.30 | 6.72 ± 0.25 | 0.343 |
| FS (%) | 62.06 ± 1.43 | 63.89 ± 4.06 | 0.679 |
| EF (%) | 93.61 ± 0.63 | 90.56 ± 2.18 | 0.209 |
| MV E velocity (m/s) | 1.28 ± 0.19 | 0.80 ± 0.14 | 0.067 |
| MV A velocity (m/s) | 0.95 ± 0.16 | 0.50 ± 0.16 | 0.076 |
| E/A | 1.39 ± 0.06 | 1.81 ± 0.17 * | 0.043 |
| e’ (m/s) | 0.06 ± 0.00 | 0.04 ± 0.00 * | 0.005 |
| E/e’ | 20.78 ± 3.38 | 20.08 ± 3.80 | 0.894 |
| E deceleration time | 79.00 ± 9.08 | 51.56 ± 6.12 * | 0.031 |
| Heart rate (1/min) | 350.33 ± 10.89 | 323.50 ± 5.00 * | 0.049 |
| UniProt ID | Gene Symbol | Protein Name | Fold Change |
|---|---|---|---|
| P09895 | Rpl5 | 60S ribosomal protein L5 | 2.60 |
| Q03626 | Mug1 | Murinoglobulin-1 | 1.96 |
| O35814 | Stip1 | Stress-induced-phosphoprotein 1 | 1.91 |
| P09006 | Serpina3n | Serine protease inhibitor A3N | 1.90 |
| P52873 | Pc | Pyruvate carboxylase | 1.54 |
| P02680 | Fgg | Fibrinogen gamma chain | 1.45 |
| P02564 | Myh7 | Myosin-7 | 1.42 |
| P06399 | Fga | Fibrinogen alpha chain | 1.42 |
| P01026 | C3 | Complement C3 | 1.35 |
| D3ZWC6 | Sntb1 | Syntrophin, basic 1 | 1.31 |
| P25113 | Pgam1 | Phosphoglycerate mutase 1 | 1.30 |
| P29147 | Bdh1 | D-beta-hydroxybutyrate dehydrogenase | 1.30 |
| Q68FP1 | Gsn | Gelsolin | 1.29 |
| P05545 | Serpina3k | Serine protease inhibitor A3K | 1.27 |
| Q5RKI0 | Wdr1 | WD repeat-containing protein 1 | 1.27 |
| P07335 | Ckb | Creatine kinase B-type | 1.25 |
| A0A0G2K542 | Ugp2 | UTP--glucose-1-phosphate uridylyltransferase | 1.22 |
| Q99PD4 | Arpc1a | Actin-related protein 2/3 complex subunit 1A | 1.22 |
| P50137 | Tkt | Transketolase | 1.22 |
| D4A5W5 | Recql4 | RecQ-like helicase 4 | 1.22 |
| P63102 | Ywhaz | 14-3-3 protein zeta/delta | 1.21 |
| P61589 | Rhoa | Transforming protein RhoA | 1.21 |
| Q08163 | Cap1 | Adenylyl cyclase-associated protein 1 | 1.21 |
| G3V885 | Myh6 | Myosin-6 | 0.83 |
| Q925Q9 | Sh3kbp1 | SH3 domain-containing kinase-binding protein 1 | 0.83 |
| F1LNF0 | Myh14 | Myosin heavy chain 14 | 0.83 |
| F1M7L9 | Uncharacterized protein | 0.82 | |
| P38650 | Dync1h1 | Cytoplasmic dynein 1 heavy chain 1 | 0.81 |
| Q925F0 | Smpx | Small muscular protein | 0.78 |
| O35115 | Fhl2 | Four and a half LIM domains protein 2 | 0.77 |
| P02401 | Rplp2 | 60S acidic ribosomal protein P2 | 0.77 |
| Q6PCU8 | Ndufv3 | NADH dehydrogenase [ubiquinone] flavoprotein 3 | 0.76 |
| P41123 | Rpl13 | 60S ribosomal protein L13 | 0.76 |
| Q5XIG9 | Mtfp1 | Mitochondrial fission process 1 | 0.74 |
| P02650 | Apoe | Apolipoprotein E | 0.68 |
| P62902 | Rpl31 | 60S ribosomal protein L31 | 0.66 |
| C0KUC6 | Lims1 | LIM and senescent cell antigen-like-containing domain protein | 0.61 |
| Q924S5 | Lonp1 | Lon protease homolog | 0.58 |
| P02466 | Col1a2 | Collagen alpha-2(I) chain | 0.52 |
| P60711 | Actb | Beta-actin | 0.47 |
| A0A0G2K1W9 | Ldhd | Lactate dehydrogenase D | 0.37 |
| M0RB63 | LOC684509 | NADH-ubiquinone oxidoreductase B9 subunit | 0.34 |
| A0A0G2KAA3 | Ndufa3 | NADH:ubiquinone oxidoreductase subunit A3 | 0.34 |
| P13697 | Me1 | NADP-dependent malic enzyme | 0.34 |
| Q9QZA6 | Cd151 | CD151 antigen | 0.28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szabó, M.R.; Pipicz, M.; Sárközy, M.; Bruszel, B.; Szabó, Z.; Csont, T. Diet-Induced Hypercholesterolemia Leads to Cardiac Dysfunction and Alterations in the Myocardial Proteome. Int. J. Mol. Sci. 2022, 23, 7387. https://doi.org/10.3390/ijms23137387
Szabó MR, Pipicz M, Sárközy M, Bruszel B, Szabó Z, Csont T. Diet-Induced Hypercholesterolemia Leads to Cardiac Dysfunction and Alterations in the Myocardial Proteome. International Journal of Molecular Sciences. 2022; 23(13):7387. https://doi.org/10.3390/ijms23137387
Chicago/Turabian StyleSzabó, Márton Richárd, Márton Pipicz, Márta Sárközy, Bella Bruszel, Zoltán Szabó, and Tamás Csont. 2022. "Diet-Induced Hypercholesterolemia Leads to Cardiac Dysfunction and Alterations in the Myocardial Proteome" International Journal of Molecular Sciences 23, no. 13: 7387. https://doi.org/10.3390/ijms23137387
APA StyleSzabó, M. R., Pipicz, M., Sárközy, M., Bruszel, B., Szabó, Z., & Csont, T. (2022). Diet-Induced Hypercholesterolemia Leads to Cardiac Dysfunction and Alterations in the Myocardial Proteome. International Journal of Molecular Sciences, 23(13), 7387. https://doi.org/10.3390/ijms23137387