
Bamboo Lignin Fractions with In Vitro Tyrosinase Inhibition Activity Downregulate Melanogenesis in B16F10 Cells via PKA/CREB Signaling Pathway



Abstract
:
1. Introduction
2. Results
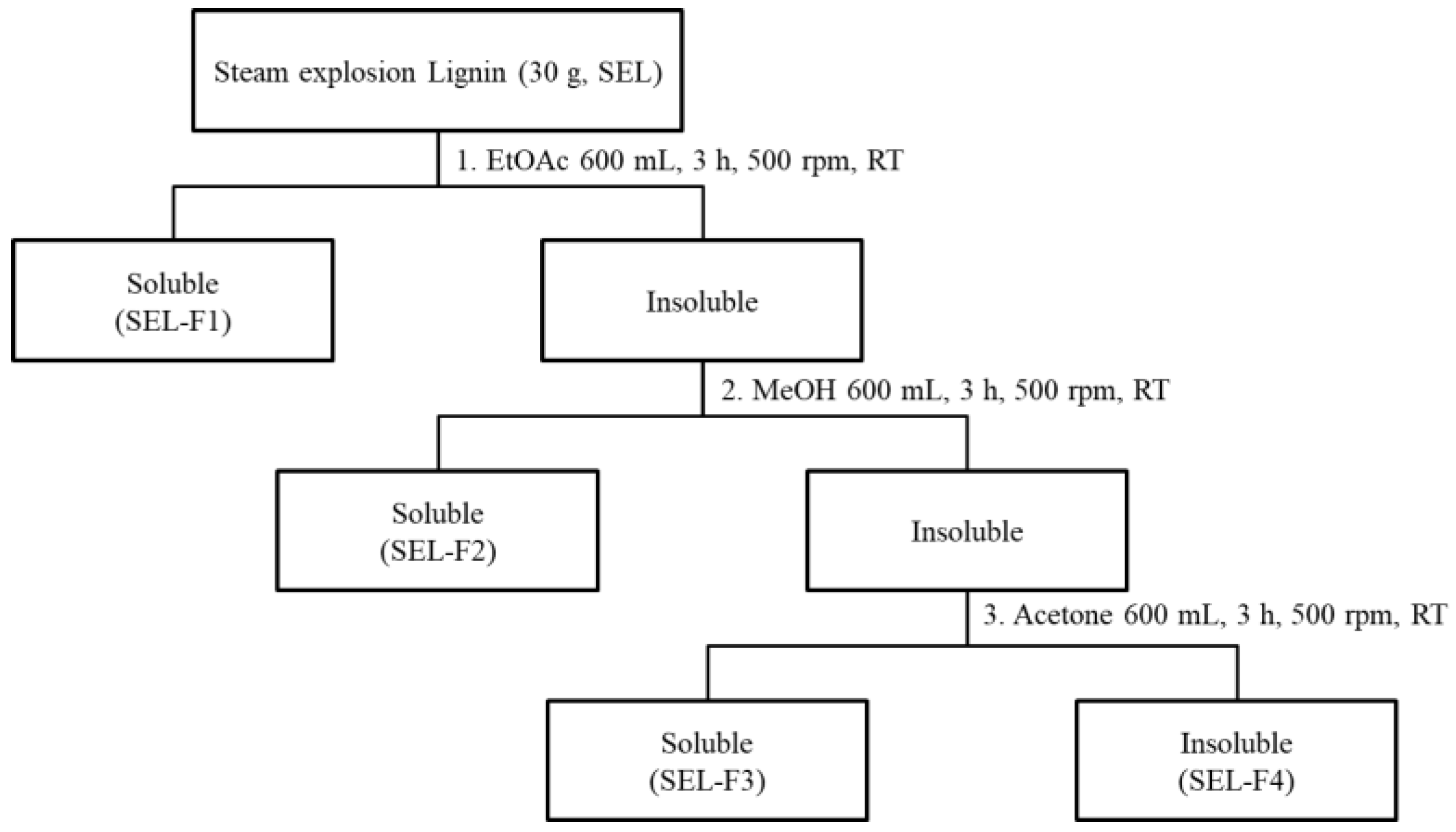
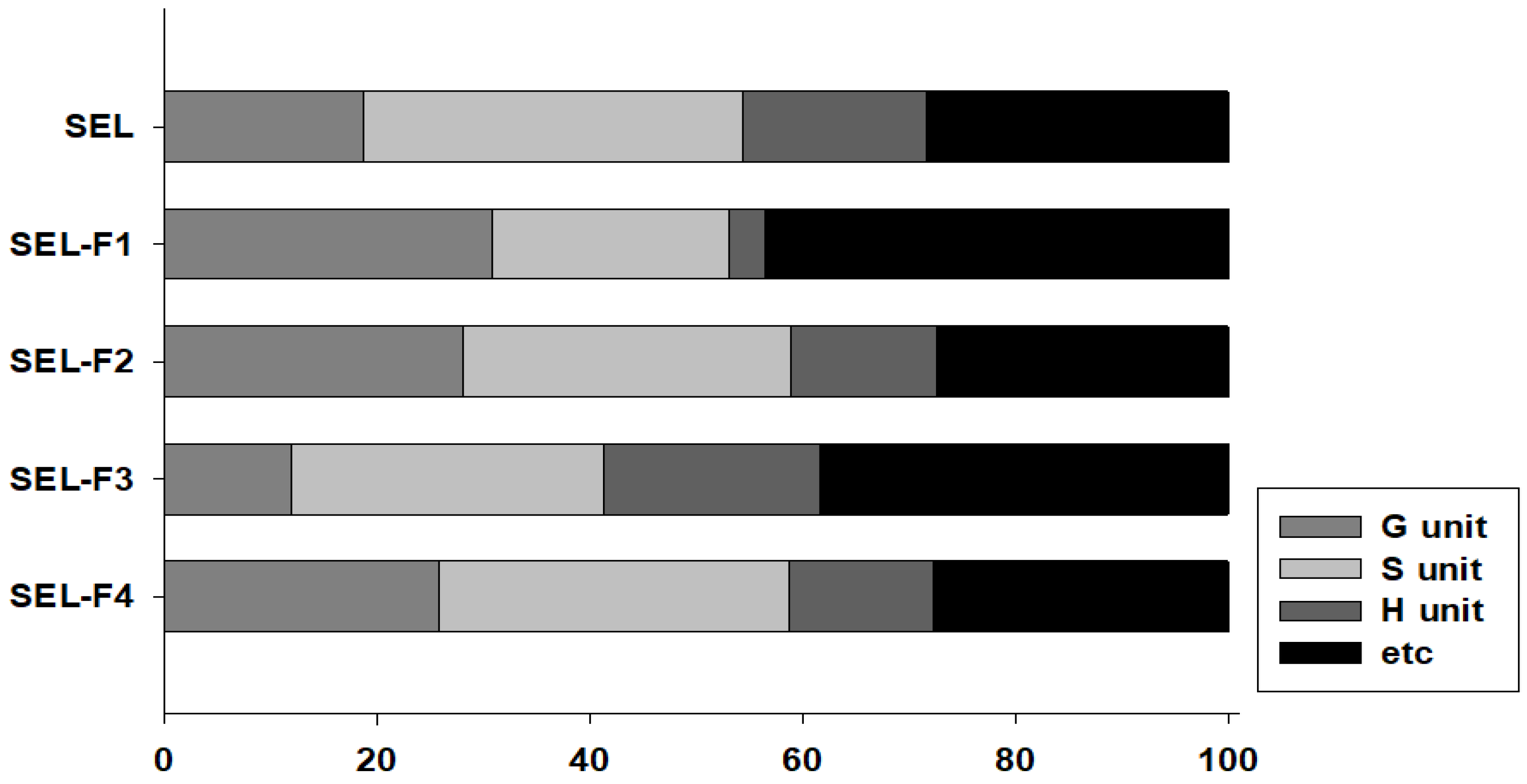
2.1. Fractionation and Characterization of Steam-Exploded Bamboo Lignin
2.1.1. Separation Yield and Molecular Weight
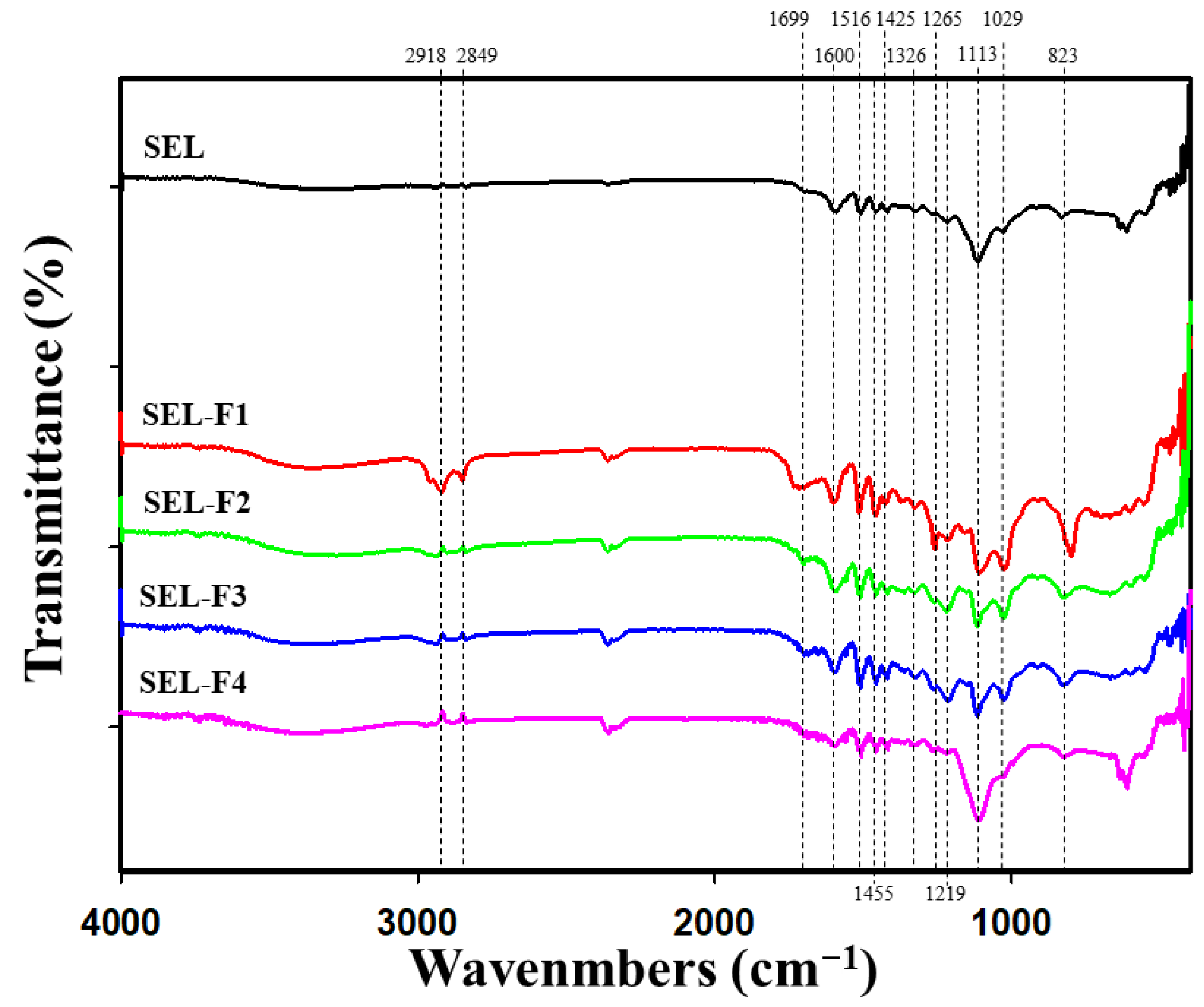
2.1.2. FT-IR
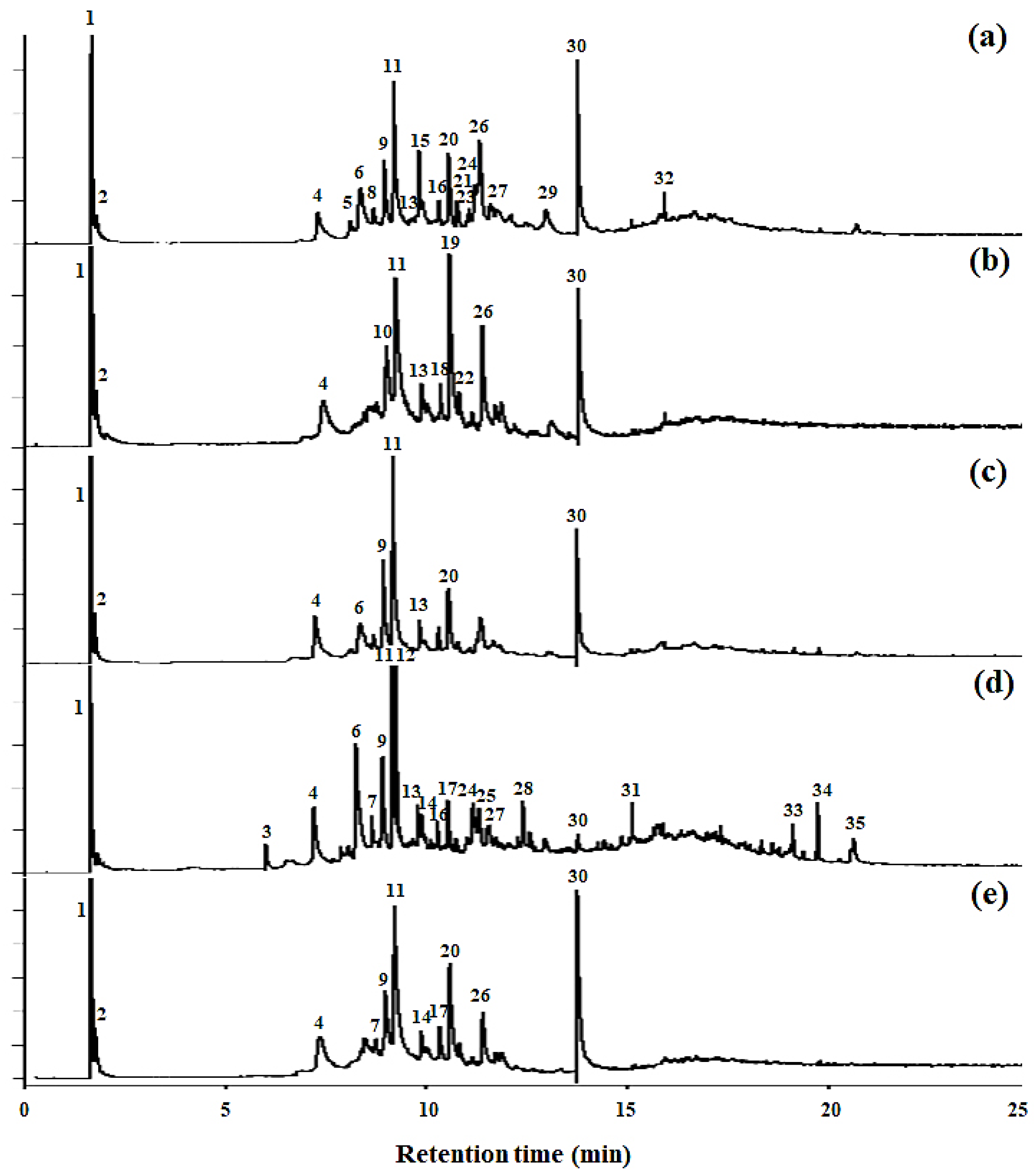
2.1.3. Py-GC/MS
2.1.4. NMR Analysis
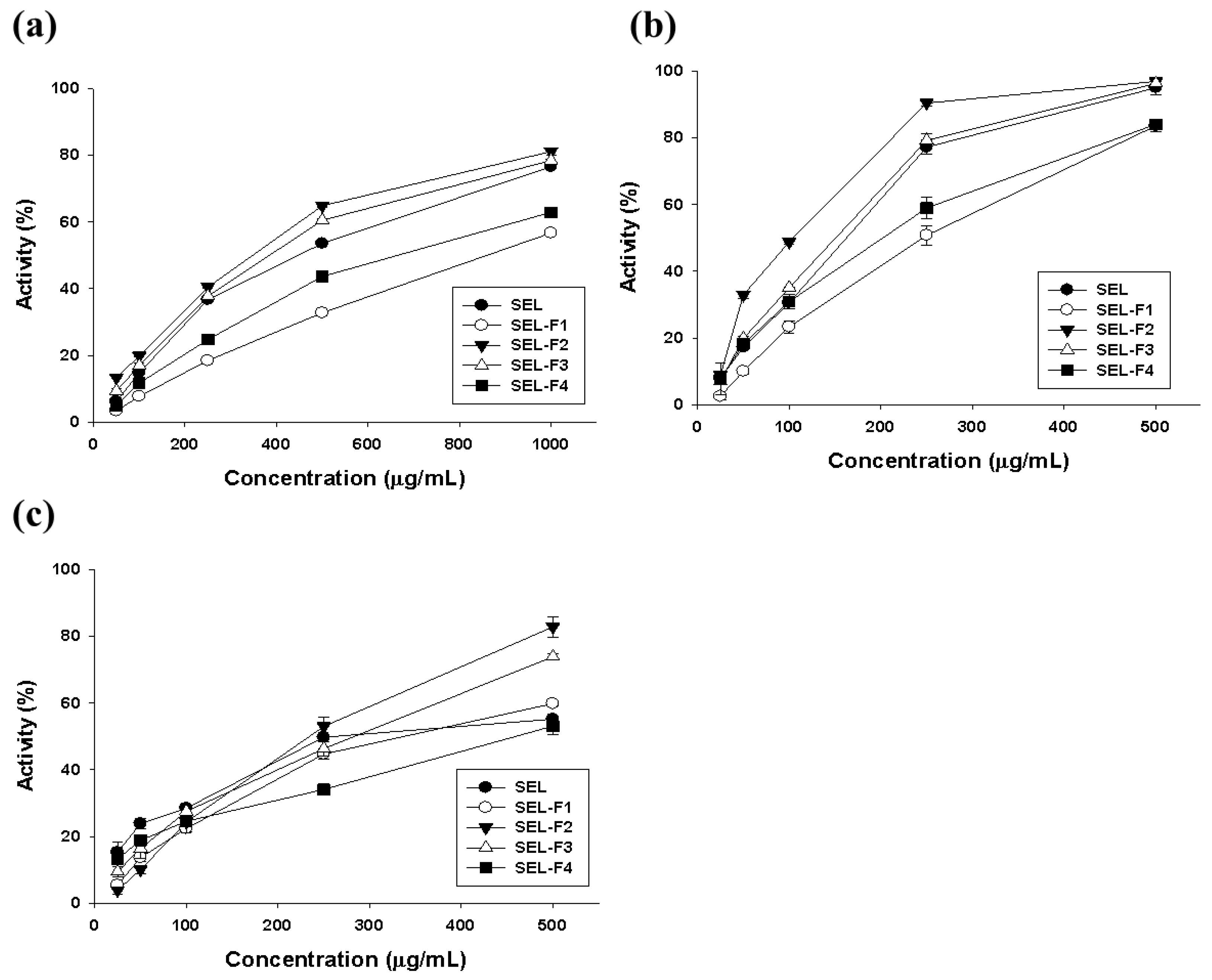
2.2. Antioxidant Activity of Lignin Fractions
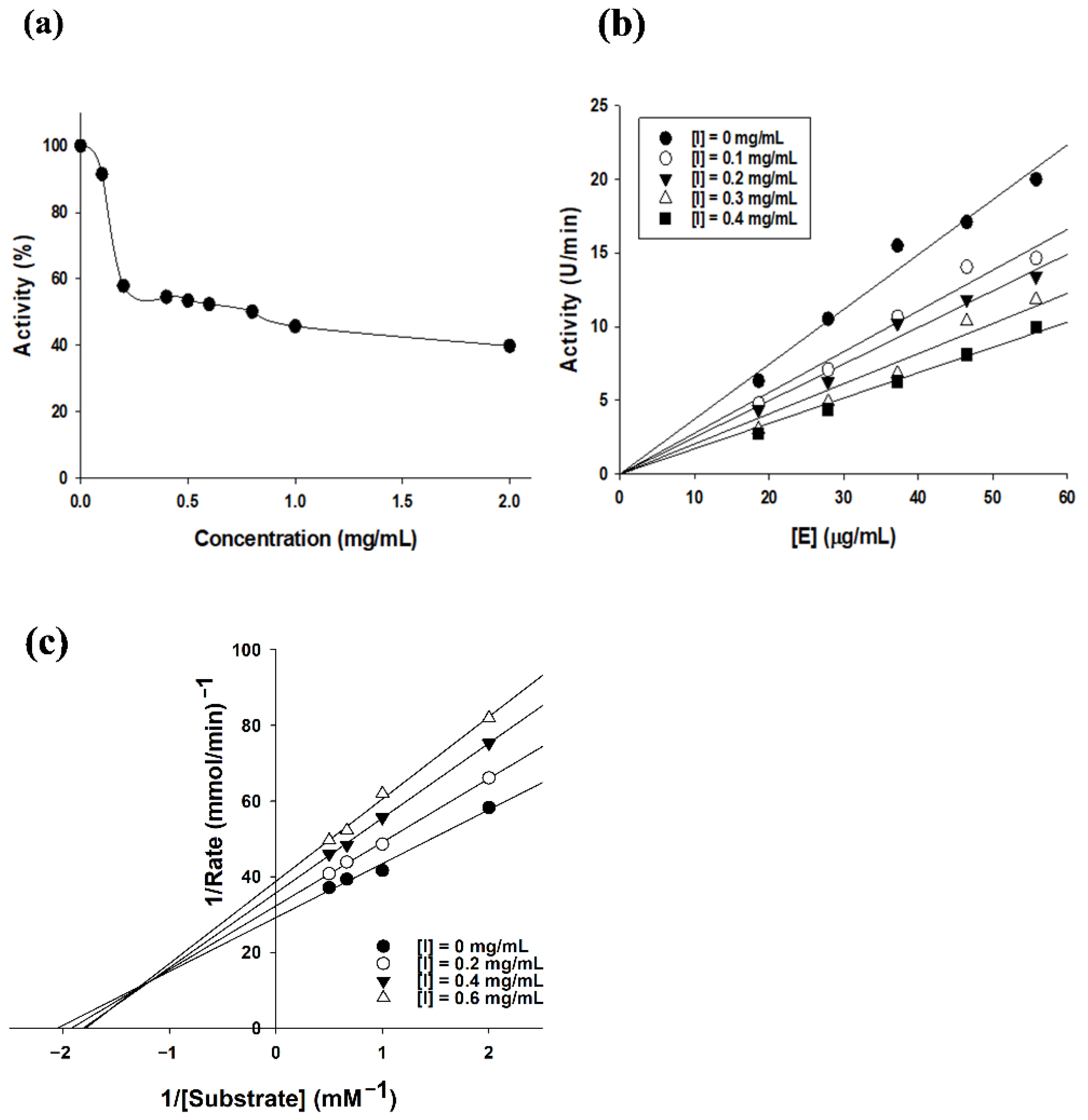
2.3. Tyrosinase Inhibition Kinetics of SEL-F2
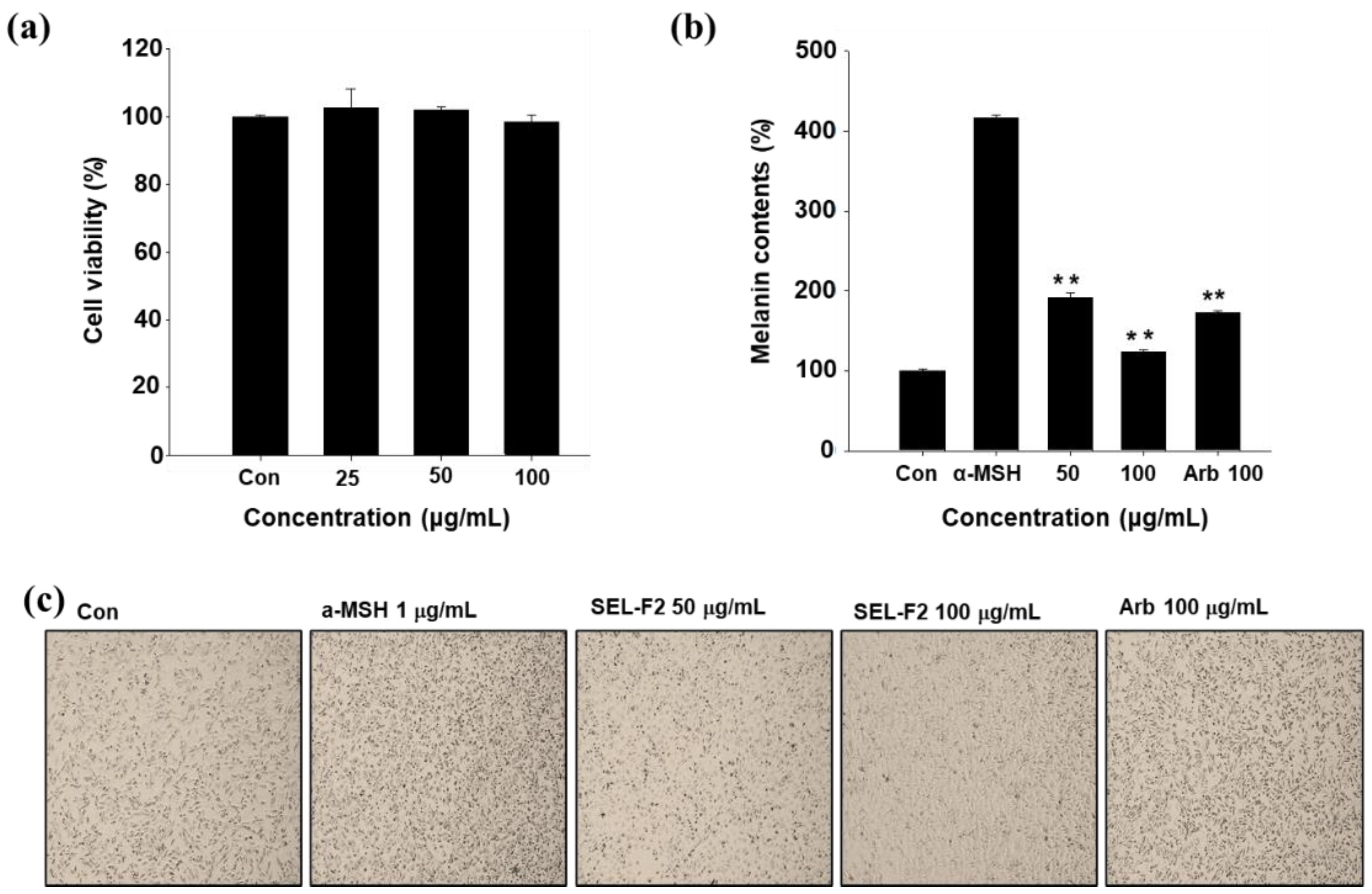
2.4. Inhibition of Melanin Synthesis in B16F10 Cell Line
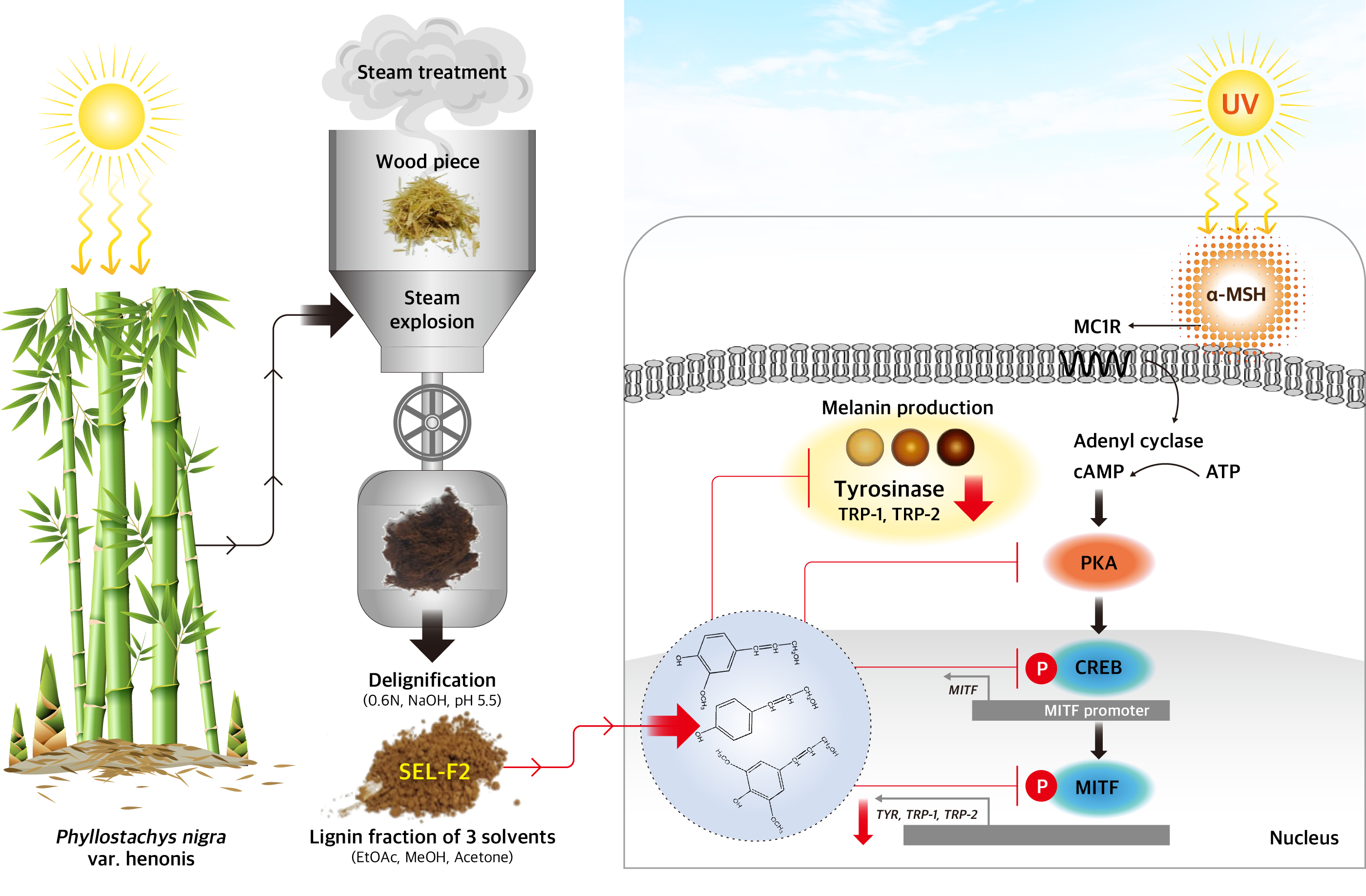
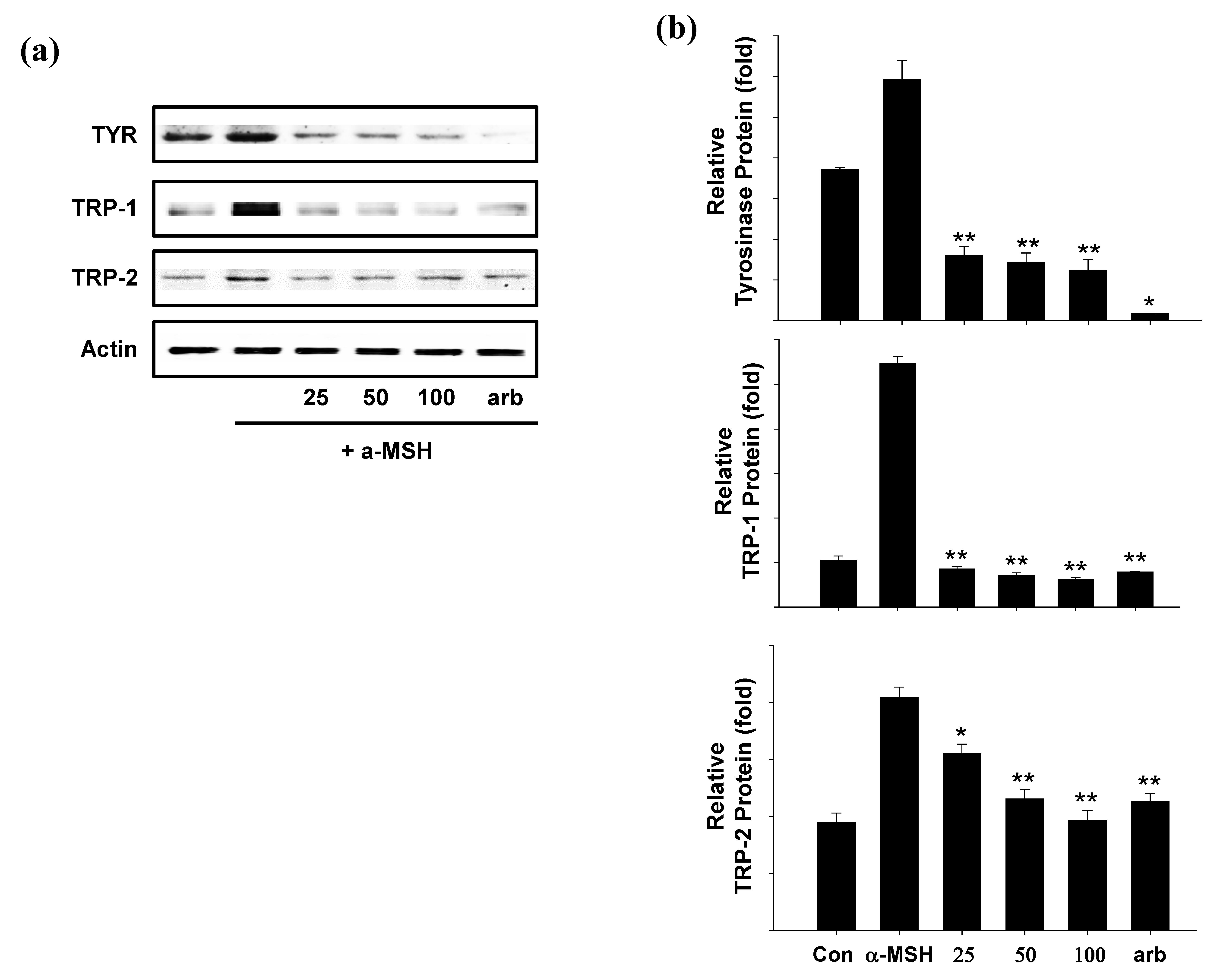
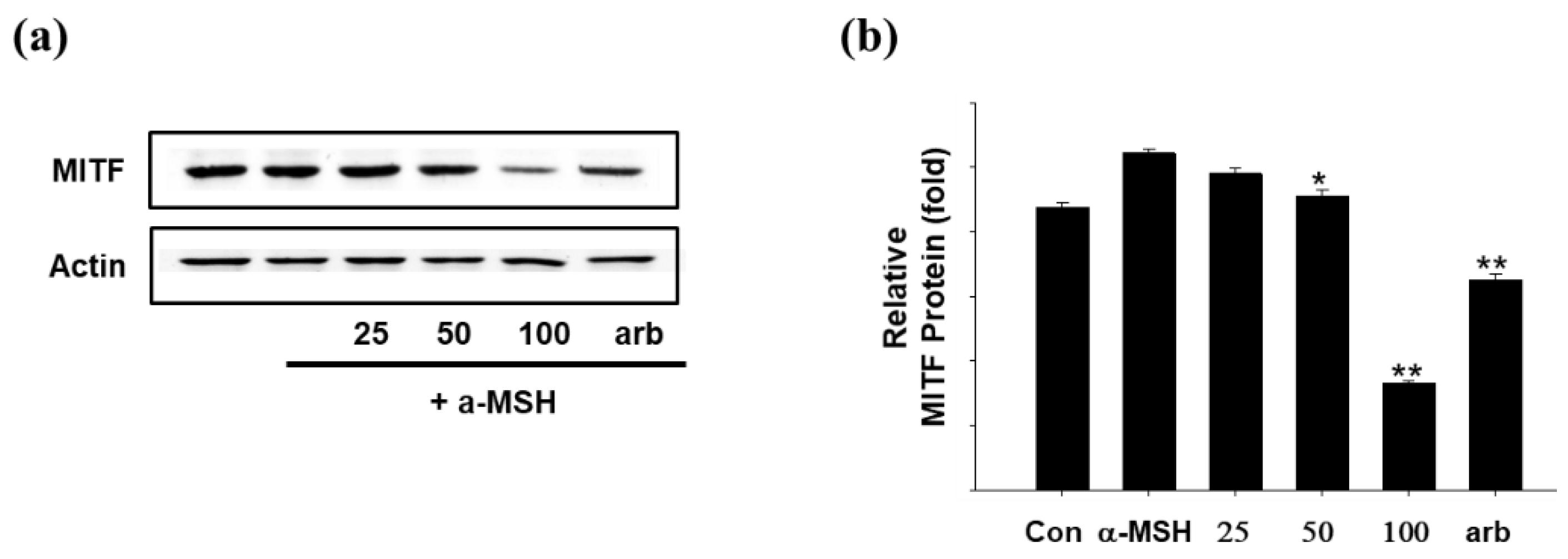
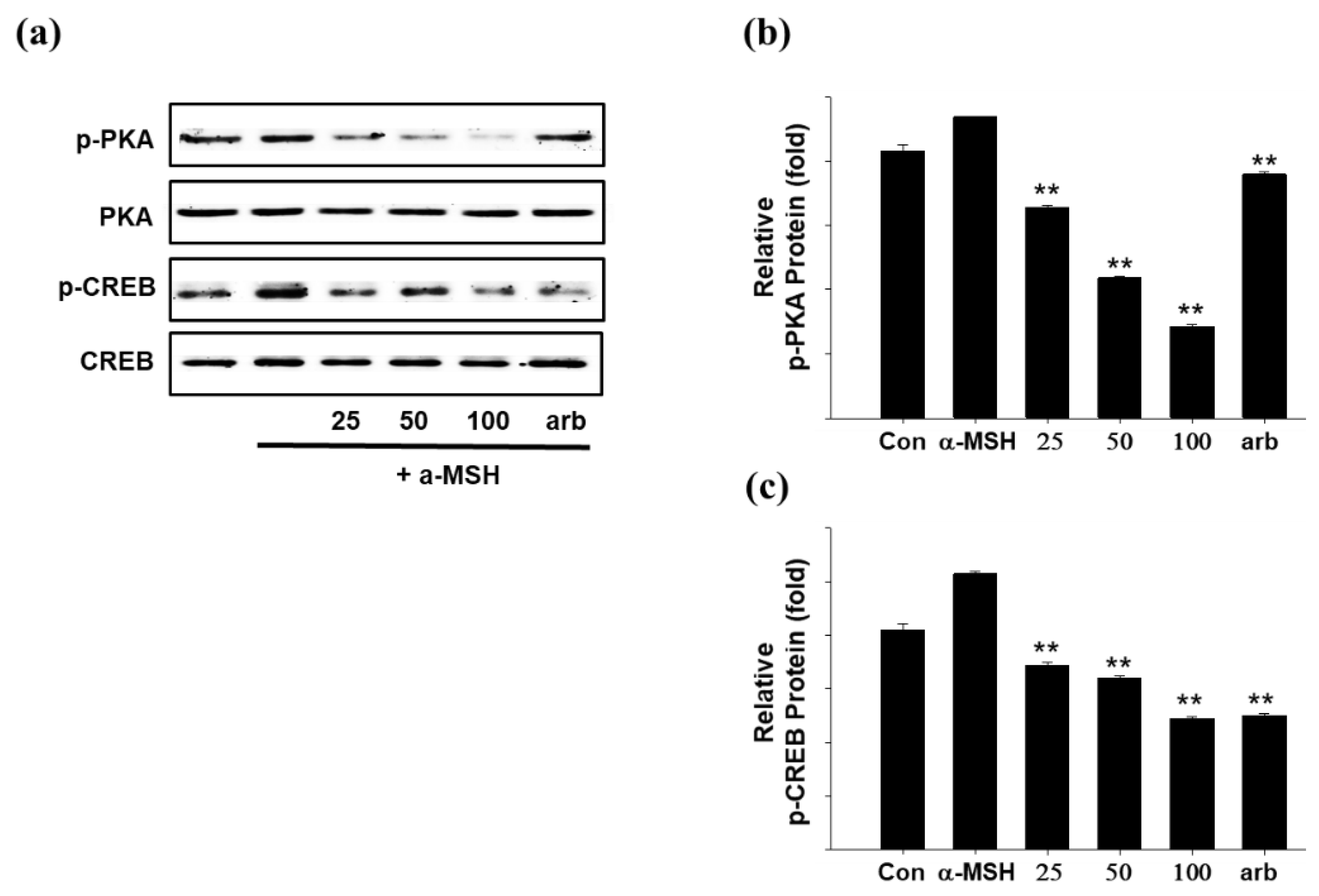
2.5. Effect of SEL-F2 on Anti-Melanogenesis-Related Proteins in B16F10 Cells
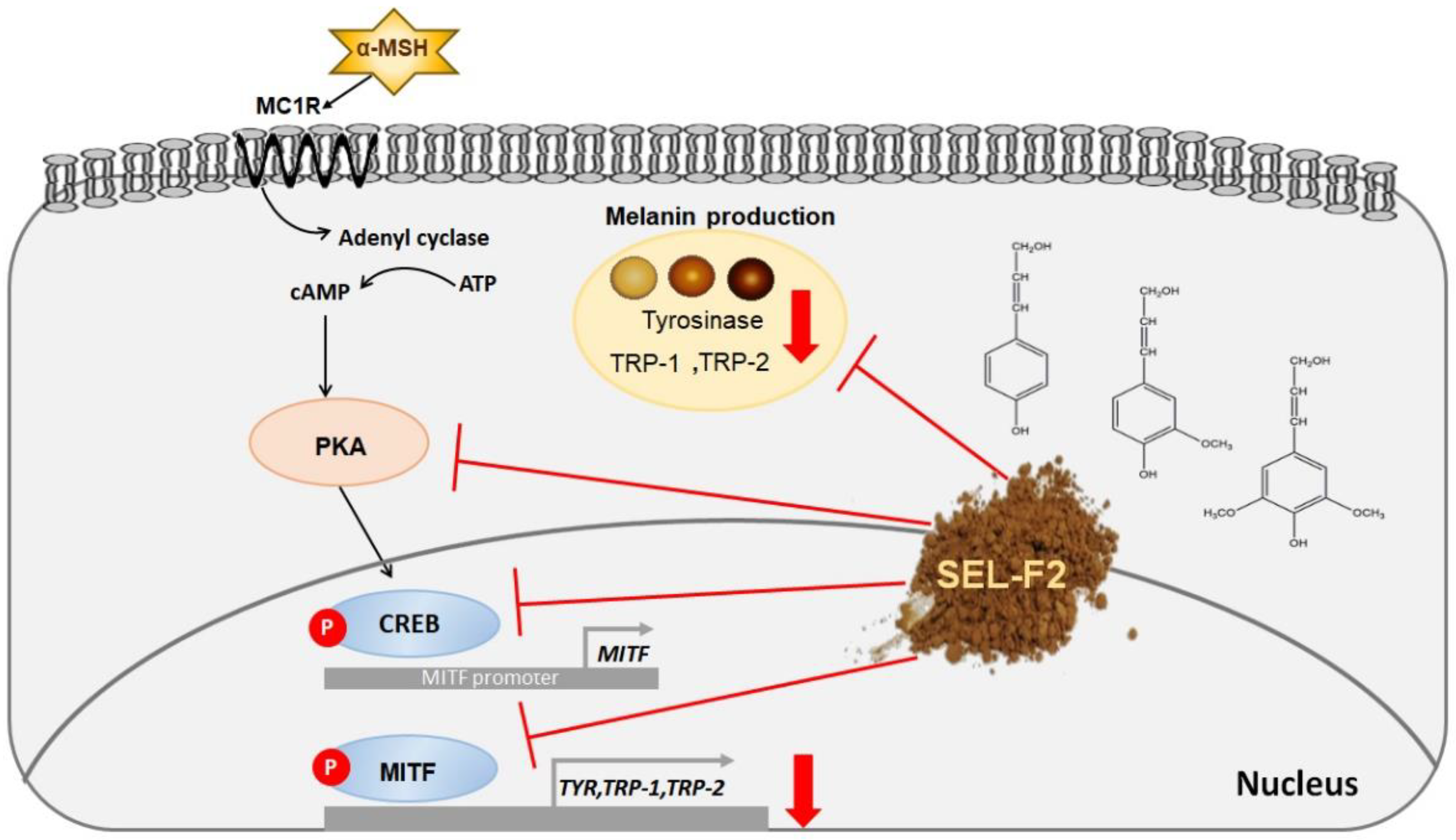
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Bamboo Biomass and Lignin Preparation
4.3. GPC Analysis
4.4. Physico-Chemical Characterization of Lignin
4.5. Antioxidant Activity Assay
4.6. Total Polyphenol Content
4.7. Anti-Melanogenic Activity Assay
4.7.1. Tyrosinase Inhibition Assay and Enzyme Kinetics Assay
4.7.2. MTT Assay for SEL-F2
4.7.3. Melanin Contents in B16F10 Cell Line
4.7.4. Immunoblot Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qian, W.; Liu, W.; Zhu, D.; Cao, Y.; Tang, A.; Gong, G.; Su, H. Natural skin-whitening compounds for the treatment of melanogenesis. Exp. Ther. Med. 2020, 20, 173–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.H.; Shin, H.J. Anti-melanogenesis effect of quercetin. Cosmetics 2016, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- Maddaleno, A.S.; Camargo, J.; Mitjans, M.; Vinardell, M.P. Melanogenesis and Melasma Treatment. Cosmetics 2021, 8, 82. [Google Scholar] [CrossRef]
- Scurlock, J.M.; Dayton, D.C.; Hames, B. Bamboo: An overlooked biomass resource? Biomass Bioenergy 2000, 19, 229–244. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Dai, C.; Liu, R.; Lian, C.; Yuan, J.; Fang, C.; Fei, B. Influence of cell wall structure on the fracture behavior of bamboo (Phyllostachys edulis) fibers. Ind. Crops Prod. 2020, 155, 112787. [Google Scholar] [CrossRef]
- Jiang, L.; Belwal, T.; Huang, H.; Ge, Z.; Limwachiranon, J.; Zhao, Y.; Li, L.; Ren, G.; Luo, Z. Extraction and characterization of phenolic compounds from bamboo shoot shell under optimized ultrasonic-assisted conditions: A potential source of nutraceutical compounds. Food Bioproc. Technol. 2019, 12, 1741–1755. [Google Scholar] [CrossRef]
- Ververis, C.; Georghiou, K.; Christodoulakis, N.; Santas, P.; Santas, R. Fiber dimensions, lignin and cellulose content of various plant materials and their suitability for paper production. Ind. Crops Prod. 2004, 19, 245–254. [Google Scholar] [CrossRef]
- Meng, X.; Bhagia, S.; Wang, Y.; Zhou, Y.; Pu, Y.; Dunlap, J.R.; Shuai, L.; Ragauskas, A.J.; Yoo, C.G. Effects of the advanced organosolv pretreatment strategies on structural properties of woody biomass. Ind. Crops Prod. 2020, 146, 112144. [Google Scholar] [CrossRef]
- Yang, S.H.; Shin, H.J. Lignin as bio-industrial materials: Focused on cosmetic ingredients. KSBB J. 2018, 33, 215–226. [Google Scholar] [CrossRef]
- Azadi, P.; Inderwildi, O.R.; Farnood, R.; King, D.A. Liquid fuels, hydrogen and chemicals from lignin: A critical review. Renew. Sust. Energ. Rev. 2013, 21, 506–523. [Google Scholar] [CrossRef]
- Ekielski, A.; Mishra, P.K. Lignin for bioeconomy: The present and future role of technical lignin. Int. J. Mol. Sci. 2020, 22, 63. [Google Scholar] [CrossRef]
- Gordobil, O.; Herrera, R.; Yahyaoui, M.; İlk, S.; Kaya, M.; Labidi, J. Potential use of Kraft and Organosolv lignins as a natural additive for healthcare products. RSC Adv. 2018, 8, 24525–24533. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Wang, G.; Xia, Y.; Sui, W.; Si, C. Functionality study of lignin as a tyrosinase inhibitor: Influence of lignin heterogeneity on anti-tyrosinase activity. Int. J. Biol. Macromol 2019, 128, 107–113. [Google Scholar] [CrossRef]
- Wang, G.; Xia, Y.; Sui, W.; Si, C. Lignin as a novel tyrosinase inhibitor: Effects of sources and isolation processes. ACS Sustain. Chem. Eng. 2018, 6, 9510–9518. [Google Scholar] [CrossRef]
- Sun, S.N.; Li, M.F.; Yuan, T.Q.; Xu, F.; Sun, R.C. Sequential extractions and structural characterization of lignin with ethanol and alkali from bamboo (Neosinocalamus affinis). Ind. Crops Prod. 2012, 37, 51–60. [Google Scholar] [CrossRef]
- Wang, G.; Chen, H. Fractionation of alkali-extracted lignin from steam-exploded stalk by gradient acid precipitation. Sep. Purif. Technol. 2013, 105, 98–105. [Google Scholar] [CrossRef]
- Yang, H.; Li, J.; Xu, J.; Mo, L. The critical analysis of catalytic steam explosion pretreatment of corn stalk, lignin degradation, recovery, and characteristic variations. BioResources 2017, 12, 344–361. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.C.; Wen, J.L.; Xu, F.; Sun, R.C. Organosolv-and alkali-soluble hemicelluloses degraded from Tamarix austromongolica: Characterization of physicochemical, structural features and thermal stability. Polym. Degrad. Stab. 2011, 96, 1478–1488. [Google Scholar] [CrossRef]
- Sun, S.L.; Wen, J.L.; Ma, M.G.; Sun, R.C.; Jones, G.L. Structural features and antioxidant activities of degraded lignins from steam exploded bamboo stem. Ind. Crops Prod. 2014, 56, 128–136. [Google Scholar] [CrossRef]
- Rho, S.; Chung, H.S.; Kang, M.; Lee, E.; Cho, C.; Kim, H.; Park, S.; Kim, H.Y.; Hong, M.; Shin, M. Inhibition of production of reactive oxygen species and gene expression profile by treatment of ethanol extract of Moutan Cortex Radicis in oxidative stressed PC12 cells. Biol. Pharm. Bull. 2005, 28, 661–666. [Google Scholar] [CrossRef] [Green Version]
- Salanti, A.; Zoia, L.; Orlandi, M.; Zanini, F.; Elegir, G. Structural characterization and antioxidant activity evaluation of lignins from rice husk. J. Agric. Food Chem. 2010, 58, 10049–10055. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Ugartondo, V.; Mitjans, M.; Vinardell, M.P. Comparative antioxidant and cytotoxic effects of lignins from different sources. Bioresour. Technol. 2008, 99, 6683–6687. [Google Scholar] [CrossRef]
- Lee, S.Y.; Baek, N.; Nam, T.G. Natural, semisynthetic and synthetic tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2016, 31, 1–13. [Google Scholar] [CrossRef]
- Choi, M.H.; Jo, H.G.; Yang, J.H.; Ki, S.H.; Shin, H.J. Antioxidative and anti-melanogenic activities of bamboo stems (Phyllostachys nigra variety Henosis) via PKA/CREB-mediated MITF downregulation in B16F10 melanoma cells. Int. J. Mol. Sci. 2018, 19, 409. [Google Scholar] [CrossRef] [Green Version]
- Ishii, N.; Ryu, M.; Suzuki, Y.A. Lactoferrin inhibits melanogenesis by down-regulating MITF in melanoma cells and normal melanocytes. Biochem. Cell Biol. 2017, 95, 119–125. [Google Scholar] [CrossRef]
- Hsu, J.Y.; Lin, H.H.; Li, T.S.; Tseng, C.Y.; Wong, Y.; Chen, J.H. Anti-melanogenesis effects of lotus seedpod in vitro and in vivo. Nutrients 2020, 12, 3535. [Google Scholar] [CrossRef]
- Sadeghifar, H.; Ragauskas, A. Lignin as a UV light blocker—A review. Polymers 2020, 12, 1134. [Google Scholar] [CrossRef]
- Qian, Y.; Qiu, X.; Zhu, S. Sunscreen performance of lignin from different technical resources and their general synergistic effect with synthetic sunscreens. ACS Sustain. Chem. Eng. 2016, 4, 4029–4035. [Google Scholar] [CrossRef]
- Deng, Y.; Feng, X.; Zhou, M.; Qian, Y.; Yu, H.; Qiu, X. Investigation of aggregation and assembly of alkali lignin using iodine as a probe. Biomacromolecules 2011, 12, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zheng, L.; Xu, Q.; Xu, L.; Wang, D.; Li, J.; Luc, G.; Huang, C.; Wang, Y. Unveiling the structural properties of water-soluble lignin from gramineous biomass by autohydrolysis and its functionality as a bioactivator (anti-inflammatory and antioxidative). Int. J. Biol. Macromol. 2021, 191, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Bos, J.D.; Meinardi, M.M. The 500 Dalton rule for the skin penetration of chemical compounds and drugs. Exp. Dermatol. 2000, 9, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.N.; Cao, X.F.; Xu, F.; Sun, R.C.; Jones, G.L. Structural features and antioxidant activities of lignins from steam-exploded bamboo (Phyllostachys pubescens). J. Agric. Food Chem. 2014, 62, 5939–5947. [Google Scholar] [CrossRef]
- Li, M.F.; Sun, S.N.; Xu, F.; Sun, R.C. Sequential solvent fractionation of heterogeneous bamboo organosolv lignin for value-added application. Sep. Purif. Technol. 2012, 101, 18–25. [Google Scholar] [CrossRef]
- Wang, G.; Chen, H. Carbohydrate elimination of alkaline-extracted lignin liquor by steam explosion and its methylolation for substitution of phenolic adhesive. Ind. Crops Prod. 2014, 53, 93–101. [Google Scholar] [CrossRef]
- Joyeux, M.; Mortier, F.; Fleurentin, J. Screening of antiradical, antilipoperoxidant and hepatoprotective effects of nine plant extracts used in Caribbean folk medicine. Phytother. Res. 1995, 9, 228–230. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolourising assay. Free Radic. Biol. Med. 1999, 26, 1231–1271. [Google Scholar] [CrossRef]
- Kim, Y.W.; Wu, Y.; Choi, M.H.; Shin, H.J.; Li, J. Alginate-derived elicitors enhance β-glucan content and antioxidant activities in culinary and medicinal mushroom, Sparassis latifolia. J. Fungi 2020, 6, 92. [Google Scholar] [CrossRef]
- Dinis, T.C.P.; Madeira, V.M.C.; Almeida, L.M. Action of phenolic derivates (acetoaminophen, salycilate, and 5-aminosalycilate) as inhibitors of membrane lipid peroxidation and as peroxyl radical scavengers. Arch. Biochem. Biophys. 1994, 315, 161–169. [Google Scholar] [CrossRef]
- Gülçin, İ. Antioxidant activity of caffeic acid (3, 4-dihydroxycinnamic acid). Toxicology 2006, 217, 213–220. [Google Scholar]
- Ribarova, F.; Atanassova, M. Total phenolics and flavonoids in Bulgarian fruits and vegetables. J. Univ. Chem. Technol. Metall. 2005, 40, 255–260. [Google Scholar]
- Choi, M.H.; Yang, S.H.; Kim, D.S.; Kim, N.D.; Shin, H.J.; Liu, K. Novel quercetin derivative of 3, 7-dioleylquercetin shows less toxicity and highly potent tyrosinase inhibition activity. Int. J. Mol. Sci. 2021, 22, 4264. [Google Scholar] [CrossRef]
- Fan, M.; Zhang, G.; Hu, X.; Xu, X.; Gong, D. Quercetin as a tyrosinase inhibitor: Inhibitory activity, conformational change and mechanism. Food Res. Int. 2017, 100, 226–233. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Q.; Song, K.; Xie, J. Inhibitory effects of salicylic acid family compounds on the diphenolase activity of mushroom tyrosinase. Food Chem. 2006, 95, 579–584. [Google Scholar] [CrossRef]
- Hridya, H.; Amrita, A.; Sankari, M.; Doss, C.G.P.; Gopalakrishnan, M.; Gopalakrishnan, C.; Siva, R. Inhibitory effect of brazilein on tyrosinase and melanin synthesis: Kinetics and in silico approach. Int. J. Biol. Macromol. 2015, 81, 228–234. [Google Scholar] [CrossRef]
- Hosoi, J.; Abe, E.; Suda, T.; Kuroki, T. Regulation of melanin synthesis of B16 mouse melanoma cells by 1α, 25-dihydroxyvitamin D3 and retinoic acid. Cancer Res. 1985, 45, 1474–1478. [Google Scholar]
- Yang, J.H.; Choi, M.H.; Yang, S.H.; Cho, S.S.; Park, S.J.; Shin, H.J.; Ki, S.H. Potent anti-Inflammatory and antiadipogenic properties of bamboo (Sasa coreana Nakai) leaves extract and its major constituent flavonoids. J. Agric. Food Chem. 2017, 65, 6665–6673. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Yield (%) | GPC | ||
|---|---|---|---|---|
| Mw | Mn | PDI | ||
| SEL | - | 4118 ± 7.76 | 2342 ± 24.90 | 1.759 ± 0.02 |
| SEL-F1 | 0.27 | 3512 ± 25.72 | 2110 ± 8.16 | 1.664 ± 0.02 |
| SEL-F2 | 36.43 | 3914 ± 12.66 | 2251 ± 17.56 | 1.739 ± 0.01 |
| SEL-F3 | 1.97 | 3230 ± 40.82 | 2046 ± 19.19 | 1.579 ± 0.03 |
| SEL-F4 | 60.33 | 3980 ± 16.33 | 2244 ± 13.06 | 1.774 ± 0.02 |
| No. | Compound | SEL | SEL-F1 | SEL-F2 | SEL-F3 | SEL-F4 |
|---|---|---|---|---|---|---|
| 1 | Carbon dioxide | 18.85 | 9.33 | 15.49 | 21.11 | 7.89 |
| 2 | Acetaldehyde | 3.86 | - | 3.95 | 3.39 | 2.38 |
| 3 | 1-Bromo-2-methyl-cyclohexane | - | 1.57 | - | - | - |
| 4 | 2-Methoxy-phenol | 8.09 | 8.68 | 8.26 | 7.55 | 3.87 |
| 5 | 4-Methoxy-2-methylphenol | - | - | - | - | 1.86 |
| 6 | 2,3-Dihydro-benzofuran | - | 14.96 | 7.95 | - | 10.24 |
| 7 | 1,5-Heptandien-3-yne | 4.5 | 1.19 | - | - | - |
| 8 | 2-Methyl-benzaldehyde | - | - | - | - | 2.27 |
| 9 | 2-Methoxy-4-vinylphenol | 8.32 | 7.06 | 10.33 | - | 7.96 |
| 10 | 3-Methoxyacetophenone | - | - | - | 8.7 | - |
| 11 | 2,6-Dimethoxy-phenol | 21.18 | 14.56 | 22.45 | 18.65 | 14.68 |
| 12 | Propanoic acid | - | 6.49 | - | - | - |
| 13 | 4,5-Dimethoxy-2-methylphenol | - | 1.98 | 1.75 | 1.73 | 5.49 |
| 14 | 3-Methyl-2-propylcyclopent-2en-1-one | 1.18 | 3.49 | - | - | - |
| 15 | 1,5-Heptadien-3-yne | - | - | - | - | 3.59 |
| 16 | 1,1’-Ethylidenbis-benzene | - | 2.03 | - | - | 3.21 |
| 17 | 2,4-Dimethoxybenzoic acid | 2.34 | 3.72 | - | - | - |
| 18 | 4-Ethyl-2,6-dimethoxy-phenol | - | - | - | 2.16 | - |
| 19 | 4-Hydroxy-2mercaptopteridine | - | - | - | 13.87 | - |
| 20 | 4-Ethenyl-2,6-dimethoxy-phenol | 8.97 | - | 8.47 | - | 6.24 |
| 21 | 2,6-Dimethoxy-4-[(Z)-prop-1-enyl]phenol | - | 3.62 | 7.68 | - | 3.16 |
| 22 | 2,5-Dimethoxy-4-methyl-benzaldehyde | - | - | - | 2.65 | - |
| 24 | 4-Hydroxy-3,5-dimethoxy benzaldehyde | - | 5.25 | - | - | 2.59 |
| 25 | 2,6-Dimethoxy-4-(2-propenyl) phenol | 5.55 | - | - | 8.54 | 8.43 |
| 26 | 1-(3,5-Dimethoxy-4-oxidanyl-phenyl)ethanone | - | 3.04 | - | - | 1.22 |
| 27 | Acetamide | - | 2.7 | - | - | - |
| 28 | 1,5-Heptadien-3-yne | - | - | - | - | 3.53 |
| 29 | 4-4’-(1-Methylethylidene)bis-phenol | 17.15 | 1.34 | 13.66 | 11.65 | 10.33 |
| 30 | Bis(2-Methylhexyl) phthalate | - | 1.06 | - | - | - |
| 31 | 4,4’-Methylenebis(2,6-dimethoxy-phenol) | - | - | - | - | 1.02 |
| 32 | β-Progesterone | - | 1.89 | - | - | - |
| 33 | Friedelan-y-al | - | 3.51 | - | - | - |
| 34 | Syringaresinol | - | 2.52 | - | - | - |
| Assignments | δC/δH (ppm) | ||||
|---|---|---|---|---|---|
| SEL | SEL-F1 | SEL-F2 | SEL-F3 | SEL-F4 | |
| Cβ-Hβ in phenylcoumaran substructures | - | 51.6/3.56 | - | - | - |
| Cβ-Hβ in β-β’ resinol substructures | 54.2/3.04 | 53.2/3.76 | 54.23/3.04 | 54.2/3.05 | - |
| C-H in methoxyls | - | 56.2/3.74 | 56.5/3.75 | - | 56.0/3.97 |
| Cβ-Hβ in β-O-4β’ substructures | - | 59.5/3.56 | - | - | - |
| Cβ-Hβ in g-acylated β-O-4β’ substructures | - | 60.1/4.01 | - | - | - |
| Cβ-Hβ in p-hydroxycinnamyl alcohol end groups | - | 66.1/3.81 | - | 63.37/3.38 | - |
| Cβ-Hβ in β-β’ resinol substructures | 71.6/4.17 | - | 71.6/4.15 | 71.6/4.17 | - |
| Cβ-Hβ in phenylcoumaran substructures | - | 70.9/4.78 | 72.6/4.82 | 72.5/4.86 | 72.3/4.86 |
| Cβ-Hβ in β-β’ resinol substructures | 85.7/4.61 | - | 85.8/4.61 | 85.7/4.62 | - |
| Cβ Hβ in β-O-4β’ linked to a S unit | 86.4/4.11 | - | 86.5/4.11 | 86.5/4.12 | 86.3/4.12 |
| Cβ-Hβ in phenylcoumaran substructures | - | - | 87.6/5.43 | 87.6/5.44 | - |
| C2,6-H2,6 in syringyl units | 104.4/6.66 | - | 104.0/6.60 | 104.3/6.69 | - |
| C2,6-H2,6 in oxidized (C=O) S units | 107.0/7.30 | 107.0/7.21 | 107.1/7.29 | 107.4/7.21 | 107.1/7.18 |
| C2-H2 in guaiacyl units | 111.7/6.98 | - | - | 110.9/6.90 | 110.08/6.89 |
| C5-H5 in guaiacyl units | 116.1/6.77 | - | 116.1/6.76 | 116.1/6.79 | 116.12/6.76 |
| C6-H6 in guaiacyl units | 119.1/6.74 | - | 119.1/6.75 | 119.1/6.75 | - |
| C2,6-H2,6 in H units | 127.7/7.15 | - | - | - | - |
| C3,5-H3,5 in p-coumaric acid | 115.4/6.51 | 116.2/6.79 | - | 116.4/6.31 | - |
| C2,6-H2,6 in p-coumaric acid | 130.0/7.41 | 130.6/7.50 | 129.6/7.37 | - | 130.7/7.41 |
| C7-H7 in p-coumaric acid | 141.3/7.32 | 144.7/7.47 | 140.2/7.27 | 144.2/7.49 | - |
| C8-H8 in p-coumaric acid | 120.3/6.27 | - | - | 116.5/6.28 | - |
| Sample | IC50 (μg/mL) | Fe2+ Chelate (%) | TPC (μg/mL) | ||
|---|---|---|---|---|---|
| DPPH | ABTS | Hydroxyl Radical | |||
| SEL | 439.06 ± 1.90 | 161.33 ± 4.33 | 249.92 ± 5.21 | 61.57 ± 2.53 | 512.23 ± 4.22 |
| SEL-F1 | 757.24 ± 10.81 | 241.89 ± 10.46 | 372.50 ± 1.27 | 49.51 ± 1.48 | 395.74 ± 8.82 |
| SEL-F2 | 466.02 ± 4.69 | 120.60 ± 2.16 | 232.46 ± 13.05 | 73.41 ± 3.22 | 578.58 ± 4.30 |
| SEL-F3 | 392.09 ± 3.08 | 154.40 ± 2.69 | 266.81 ± 15.88 | 65.02 ± 4.21 | 516.28 ± 6.87 |
| SEL-F4 | 567.12 ± 0.86 | 205.51 ± 12.86 | 429.95 ± 3.72 | 53.16 ± 3.36 | 448.33 ± 4.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, M.-H.; Yang, S.-H.; Park, W.-K.; Shin, H.-J. Bamboo Lignin Fractions with In Vitro Tyrosinase Inhibition Activity Downregulate Melanogenesis in B16F10 Cells via PKA/CREB Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 7462. https://doi.org/10.3390/ijms23137462
Choi M-H, Yang S-H, Park W-K, Shin H-J. Bamboo Lignin Fractions with In Vitro Tyrosinase Inhibition Activity Downregulate Melanogenesis in B16F10 Cells via PKA/CREB Signaling Pathway. International Journal of Molecular Sciences. 2022; 23(13):7462. https://doi.org/10.3390/ijms23137462
Chicago/Turabian StyleChoi, Moon-Hee, Seung-Hwa Yang, Won-Keun Park, and Hyun-Jae Shin. 2022. "Bamboo Lignin Fractions with In Vitro Tyrosinase Inhibition Activity Downregulate Melanogenesis in B16F10 Cells via PKA/CREB Signaling Pathway" International Journal of Molecular Sciences 23, no. 13: 7462. https://doi.org/10.3390/ijms23137462
APA StyleChoi, M. -H., Yang, S. -H., Park, W. -K., & Shin, H. -J. (2022). Bamboo Lignin Fractions with In Vitro Tyrosinase Inhibition Activity Downregulate Melanogenesis in B16F10 Cells via PKA/CREB Signaling Pathway. International Journal of Molecular Sciences, 23(13), 7462. https://doi.org/10.3390/ijms23137462