
Far-Red Light Coordinates the Diurnal Changes in the Transcripts Related to Nitrate Reduction, Glutathione Metabolism and Antioxidant Enzymes in Barley

 , ,
, , 

Abstract
:1. Introduction
2. Results
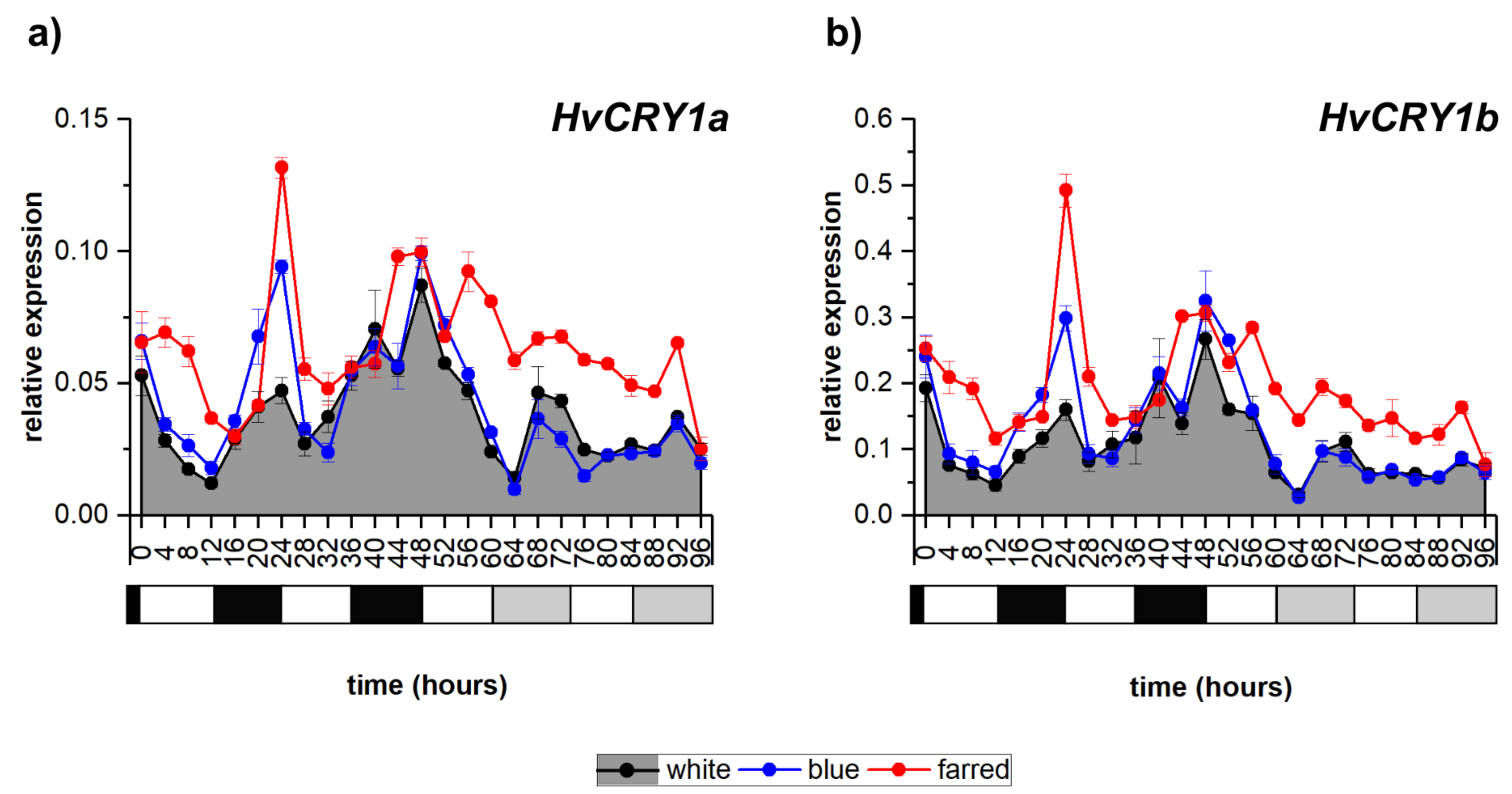
2.1. Effect of Light Spectrum on Photoreceptor-Coding Genes
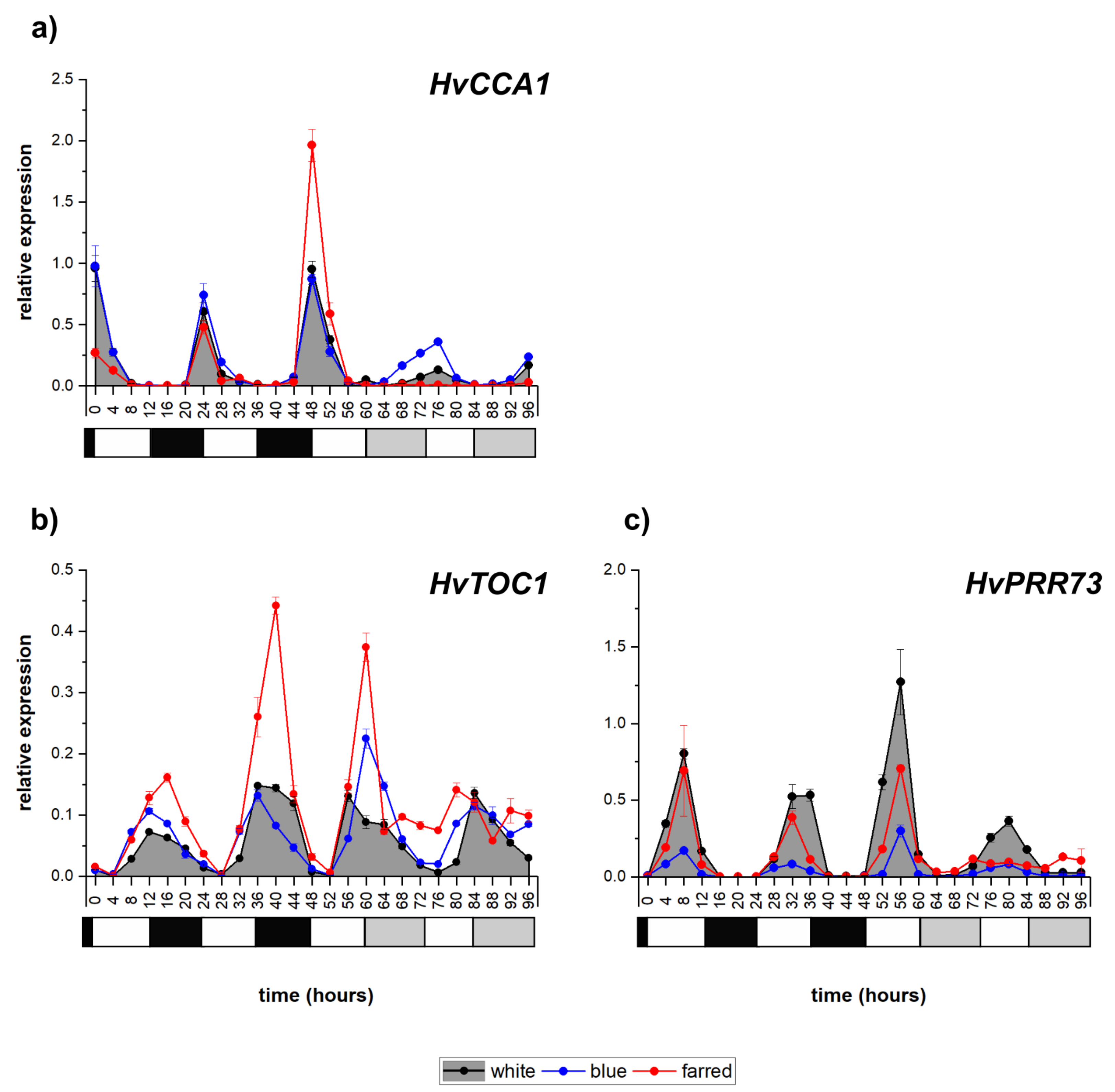
2.2. Expression of Circadian Clock Genes
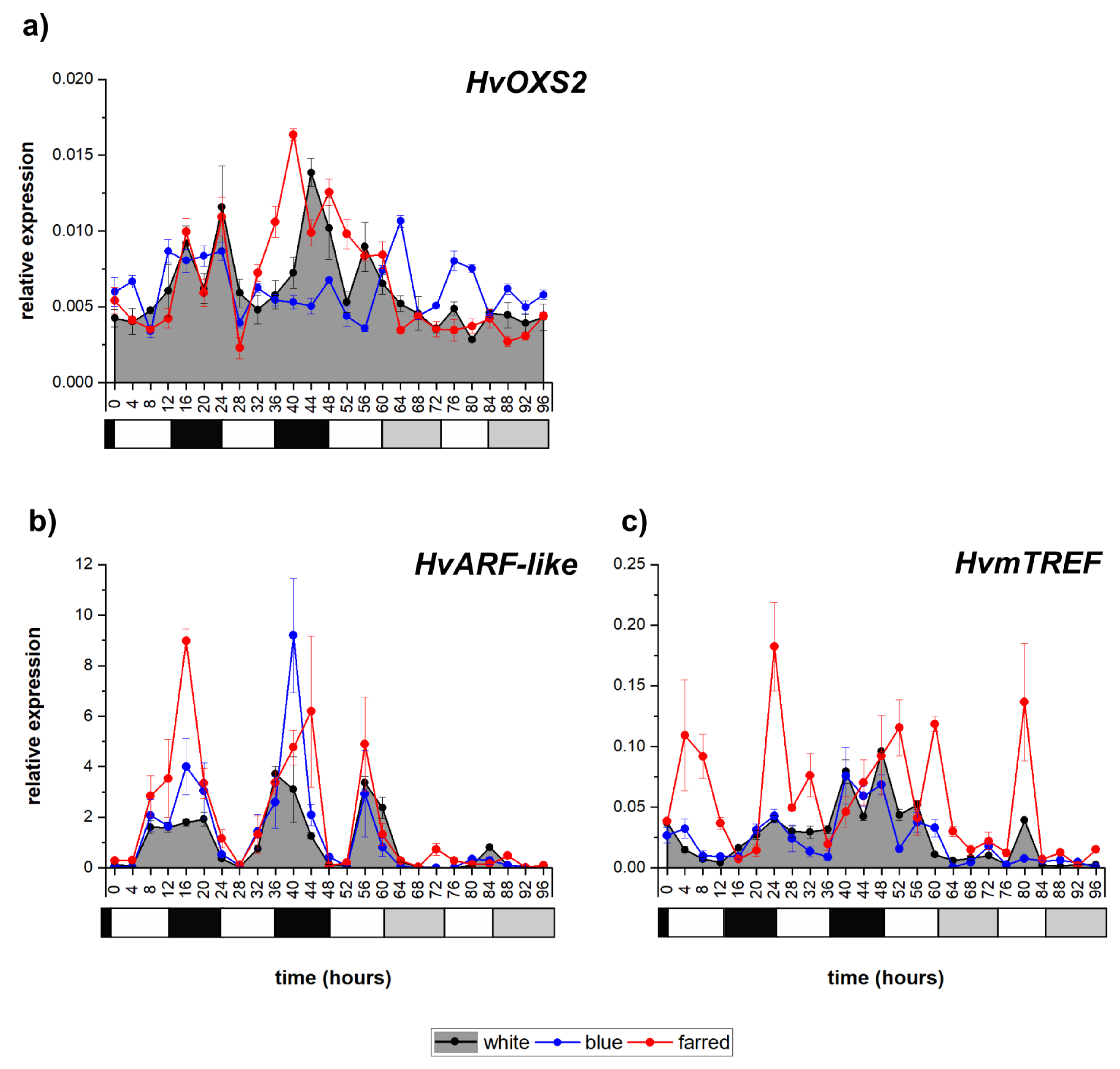
2.3. Analysis of Transcription Factors Related to Redox Regulation
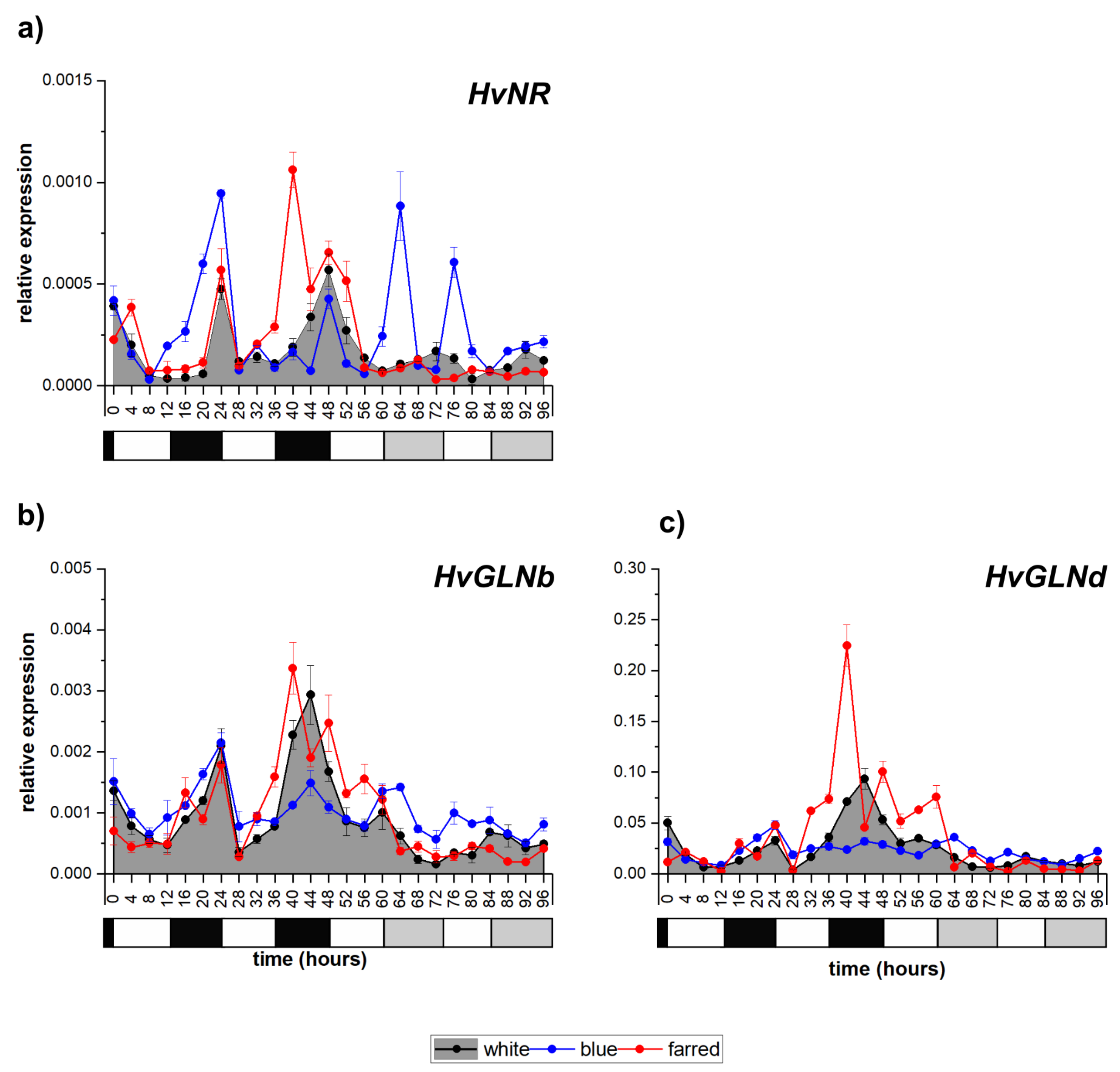
2.4. Influence of Light Spectrum on Genes Related to Nitrate Reduction
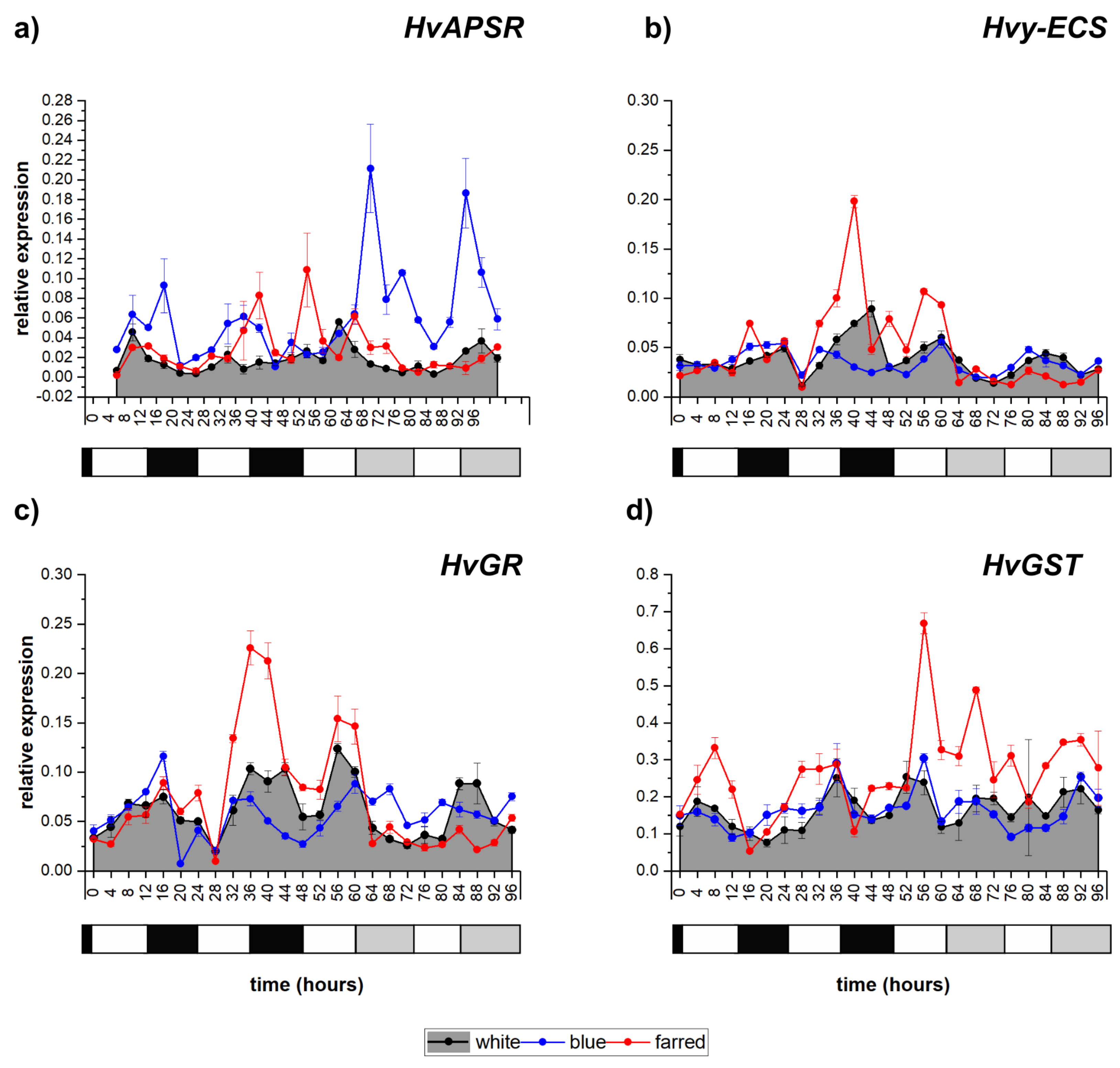
2.5. Diurnal Oscillations in Transcripts Related to Glutathione Metabolism
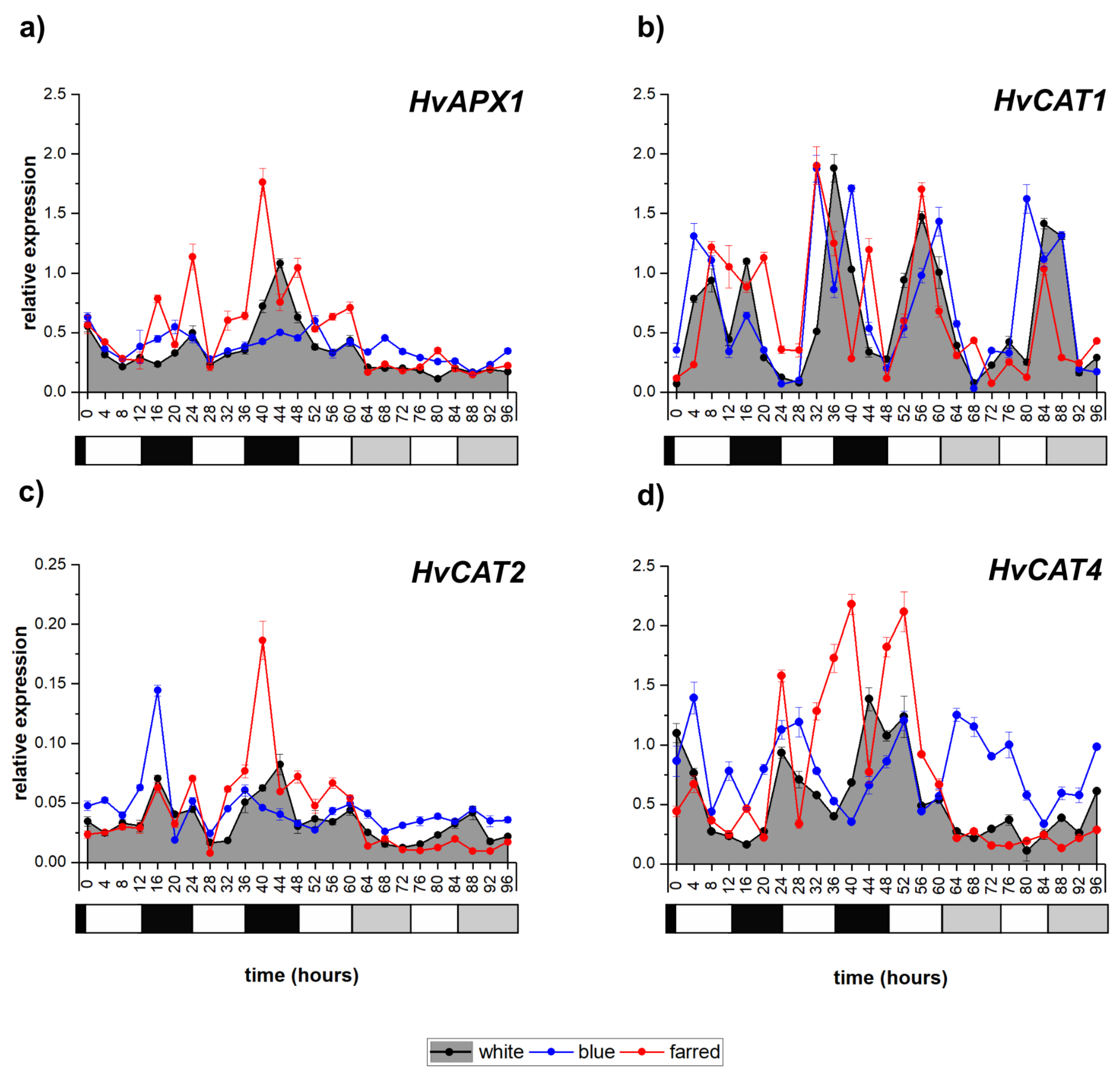
2.6. Expression of Antioxidant-Enzyme-Coding Genes
2.7. Effect of Spectrum on the Amount and Redox State of Thiols
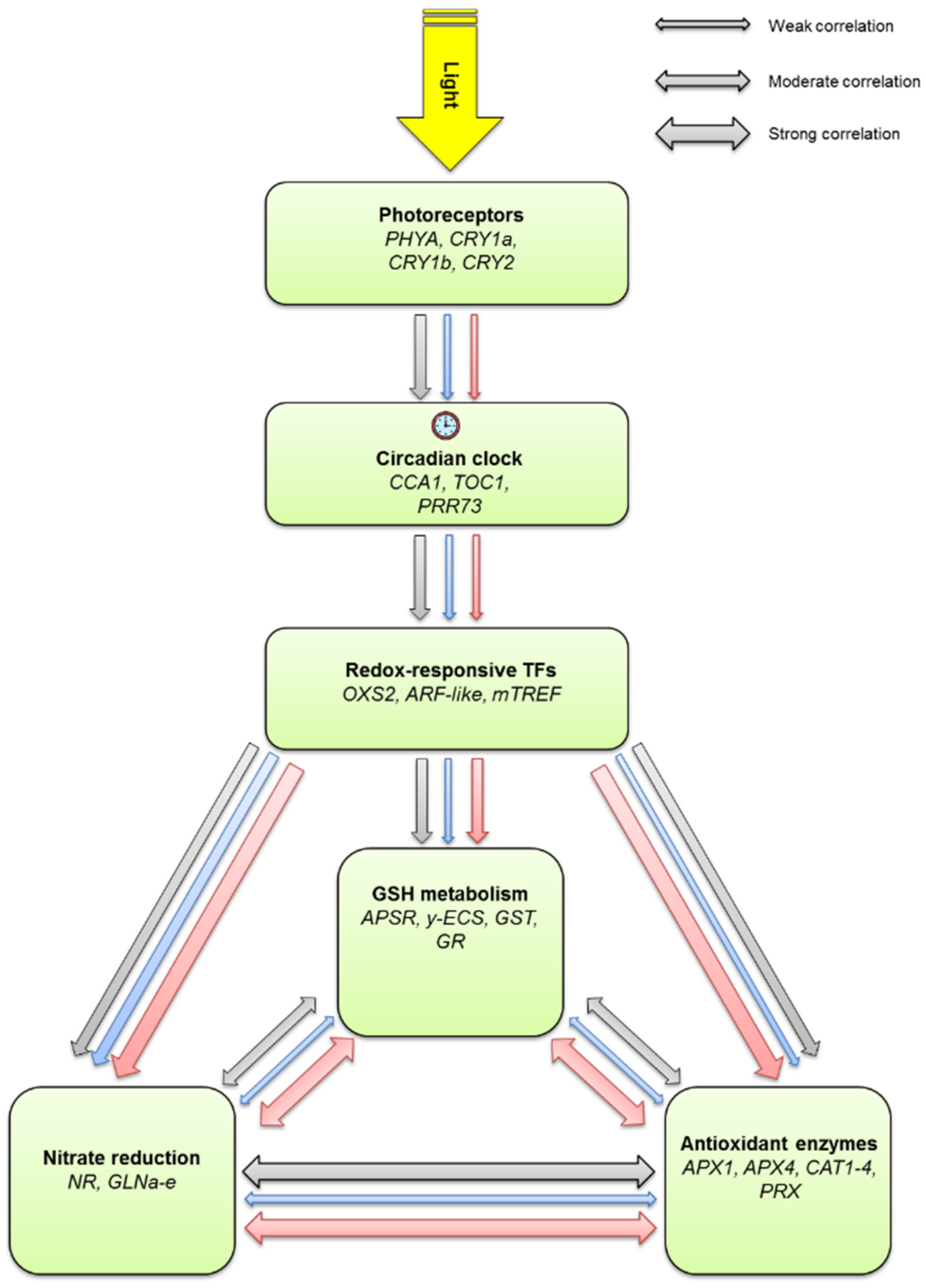
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Light Treatment and Sampling
4.3. Gene Expression Studies
4.4. Determination of Thiol Content
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franklin, K.A.; Whitelam, G.C. Light-Quality Regulation of Freezing Tolerance in Arabidopsis Thaliana. Nat. Genet. 2007, 39, 1410–1413. [Google Scholar] [CrossRef] [PubMed]
- Novák, A.; Boldizsár, Á.; Ádám, É.; Kozma-Bognár, L.; Majláth, I.; Båga, M.; Tóth, B.; Chibbar, R.; Galiba, G. Light-Quality and Temperature-Dependent CBF14 Gene Expression Modulates Freezing Tolerance in Cereals. EXBOTJ 2016, 67, 1285–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gierczik, K.; Novák, A.; Ahres, M.; Székely, A.; Soltész, A.; Boldizsár, Á.; Gulyás, Z.; Kalapos, B.; Monostori, I.; Kozma-Bognár, L.; et al. Circadian and Light Regulated Expression of CBFs and Their Upstream Signalling Genes in Barley. IJMS 2017, 18, 1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallé, Á.; Czékus, Z.; Bela, K.; Horváth, E.; Ördög, A.; Csiszár, J.; Poór, P. Plant Glutathione Transferases and Light. Front. Plant Sci. 2019, 9, 1944. [Google Scholar] [CrossRef] [Green Version]
- Somers, D.E.; Devlin, P.F.; Kay, S.A. Phytochromes and Cryptochromes in the Entrainment of the Arabidopsis Circadian Clock. Science 1998, 282, 1488–1490. [Google Scholar] [CrossRef] [Green Version]
- Cashmore, A.R.; Jarillo, J.A.; Wu, Y.-J.; Liu, D. Cryptochromes: Blue Light Receptors for Plants and Animals. Science 1999, 284, 760–765. [Google Scholar] [CrossRef]
- Chiang, C.; Olsen, J.E.; Basler, D.; Bånkestad, D.; Hoch, G. Latitude and Weather Influences on Sun Light Quality and the Relationship to Tree Growth. Forests 2019, 10, 610. [Google Scholar] [CrossRef] [Green Version]
- Hotta, C.T.; Gardner, M.J.; Hubbard, K.E.; Baek, S.J.; Dalchau, N.; Suhita, D.; Dodd, A.N.; Webb, A.A.R. Modulation of Environmental Responses of Plants by Circadian Clocks: Circadian Modulation of Plant Environmental Responses. Plant Cell Environ. 2007, 30, 333–349. [Google Scholar] [CrossRef]
- Covington, M.F.; Maloof, J.N.; Straume, M.; Kay, S.A.; Harmer, S.L. Global Transcriptome Analysis Reveals Circadian Regulation of Key Pathways in Plant Growth and Development. Genome Biol. 2008, 9, R130. [Google Scholar] [CrossRef] [Green Version]
- Campoli, C.; Shtaya, M.; Davis, S.J.; Von Korff, M. Expression Conservation within the Circadian Clock of a Monocot: Natural Variation at Barley Ppd-H1 Affects Circadian Expression of Flowering Time Genes, but Not Clock Orthologs. BMC Plant Biol. 2012, 12, 97. [Google Scholar] [CrossRef] [Green Version]
- Franklin, K.A.; Toledo-Ortiz, G.; Pyott, D.E.; Halliday, K.J. Interaction of Light and Temperature Signalling. J. Exp. Bot. 2014, 65, 2859–2871. [Google Scholar] [CrossRef] [Green Version]
- Grundy, J.; Stoker, C.; Carré, I.A. Circadian Regulation of Abiotic Stress Tolerance in Plants. Front. Plant Sci. 2015, 6, 648. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Tobin, E.M. Constitutive Expression of the CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) Gene Disrupts Circadian Rhythms and Suppresses Its Own Expression. Cell 1998, 93, 1207–1217. [Google Scholar] [CrossRef] [Green Version]
- Alabadi, D. Reciprocal Regulation Between TOC1 and LHY/CCA1 Within the Arabidopsis Circadian Clock. Science 2001, 293, 880–883. [Google Scholar] [CrossRef]
- Nohales, M.A.; Kay, S.A. Molecular Mechanisms at the Core of the Plant Circadian Oscillator. Nat. Struct. Mol. Biol. 2016, 23, 1061–1069. [Google Scholar] [CrossRef]
- Murakami, M.; Tago, Y.; Yamashino, T.; Mizuno, T. Comparative Overviews of Clock-Associated Genes of Arabidopsis Thaliana and Oryza Sativa. Plant Cell Physiol. 2007, 48, 110–121. [Google Scholar] [CrossRef]
- Murakami, M.; Ashikari, M.; Miura, K.; Yamashino, T.; Mizuno, T. The Evolutionarily Conserved OsPRR Quintet: Rice Pseudo-Response Regulators Implicated in Circadian Rhythm. Plant Cell Physiol. 2003, 44, 1229–1236. [Google Scholar] [CrossRef] [Green Version]
- Decousset, L.; Griffiths, S.; Dunford, R.P.; Pratchett, N.; Laurie, D.A. Development of STS Markers Closely Linked to the Ppd-H1 Photoperiod Response Gene of Barley (Hordeum vulgare L.). Theor. Appl. Genet. 2000, 101, 1202–1206. [Google Scholar] [CrossRef]
- Turner, A. The Pseudo-Response Regulator Ppd-H1 Provides Adaptation to Photoperiod in Barley. Science 2005, 310, 1031–1034. [Google Scholar] [CrossRef]
- Miwa, K.; Serikawa, M.; Suzuki, S.; Kondo, T.; Oyama, T. Conserved Expression Profiles of Circadian Clock-Related Genes in Two Lemna Species Showing Long-Day and Short-Day Photoperiodic Flowering Responses. Plant Cell Physiol. 2006, 47, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Serikawa, M.; Miwa, K.; Kondo, T.; Oyama, T. Functional Conservation of Clock-Related Genes in Flowering Plants: Overexpression and RNA Interference Analyses of the Circadian Rhythm in the Monocotyledon Lemna Gibba. Plant Physiol. 2008, 146, 1952–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Rowe, S.C.; Harmon, F.G. Coordination of the Maize Transcriptome by a Conserved Circadian Clock. BMC Plant Biol. 2010, 10, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filichkin, S.A.; Breton, G.; Priest, H.D.; Dharmawardhana, P.; Jaiswal, P.; Fox, S.E.; Michael, T.P.; Chory, J.; Kay, S.A.; Mockler, T.C. Global Profiling of Rice and Poplar Transcriptomes Highlights Key Conserved Circadian-Controlled Pathways and Cis-Regulatory Modules. PLoS ONE 2011, 6, e16907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foyer, C.H.; Noctor, G. Oxidant and Antioxidant Signalling in Plants—Reevaluation of the Concept. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Kocsy, G.; Tari, I.; Vanková, R.; Zechmann, B.; Gulyás, Z.; Poór, P.; Galiba, G. Redox Control of Plant Growth and Development. Plant Sci. 2013, 211, 77–91. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. REACTIVE OXYGEN SPECIES: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Boldt, R.; Scandalios, J.G. Influence of UV-Light on the Expression of the Cat2 and Cat3 Catalase Genes in Maize. Free Radic. Biol. Med. 1997, 23, 505–514. [Google Scholar] [CrossRef]
- Zhong, H.H.; McClung, C.R. The Circadian Clock Gates Expression of Two Arabidopsis Catalase Genes to Distinct and Opposite Circadian Phases. Mol. Gen. Genet. 1996, 251, 196–203. [Google Scholar] [CrossRef]
- Horling, F.; Lamkemeyer, P.; König, J.; Finkemeier, I.; Kandlbinder, A.; Baier, M.; Dietz, K.-J. Divergent Light-, Ascorbate-, and Oxidative Stress-Dependent Regulation of Expression of the Peroxiredoxin Gene Family in Arabidopsis. Plant Physiol. 2003, 131, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Lai, A.G.; Doherty, C.J.; Mueller-Roeber, B.; Kay, S.A.; Schippers, J.H.M.; Dijkwel, P.P. CIRCADIAN CLOCK-ASSOCIATED 1 Regulates ROS Homeostasis and Oxidative Stress Responses. Proc. Natl. Acad. Sci. USA 2012, 109, 17129–17134. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.S.; Green, E.W.; Zhao, Y.; Van Ooijen, G.; Olmedo, M.; Qin, X.; Xu, Y.; Pan, M.; Valekunja, U.K.; Feeney, K.A.; et al. Peroxiredoxins Are Conserved Markers of Circadian Rhythms. Nature 2012, 485, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Kubo, A.; Saji, H.; Tanaka, K.; Kondo, N. Expression of Arabidopsis Cytosolic Ascorbate Peroxidase Gene in Response to Ozone or Sulfur Dioxide. Plant Mol. Biol. 1995, 29, 479–489. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Jwa, N.-S.; Iwahashi, H.; Rakwal, R. Importance of Ascorbate Peroxidases OsAPX1 and OsAPX2 in the Rice Pathogen Response Pathways and Growth and Reproduction Revealed by Their Transcriptional Profiling. Gene 2003, 322, 93–103. [Google Scholar] [CrossRef]
- Meyer, A.J. The Integration of Glutathione Homeostasis and Redox Signaling. J. Plant Physiol. 2008, 165, 1390–1403. [Google Scholar] [CrossRef]
- Anjum, N.A.; Umar, S.; Chan, M.T. Ascorbate-Glutathione Pathway and Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar] [CrossRef]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant Responses to Stresses: Role of Ascorbate Peroxidase in the Antioxidant Protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef] [Green Version]
- Kocsy, G.; Owttrim, G.; Brander, K.; Brunold, C. Effect of Chilling on the Diurnal Rhythm of Enzymes Involved in Protection against Oxidative Stress in a Chilling-Tolerant and a Chilling-Sensitive Maize Genotype. Physiol. Plant. 1997, 99, 249–254. [Google Scholar] [CrossRef]
- Kopriva, S.; Muheim, R.; Koprivova, A.; Trachsel, N.; Catalano, C.; Suter, M.; Brunold, C. Light Regulation of Assimilatory Sulphate Reduction in Arabidopsis Thaliana. Plant J. 1999, 20, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Kopriva, S.; Rennenberg, H. Control of Sulphate Assimilation and Glutathione Synthesis: Interaction with N and C Metabolism. J. Exp. Bot. 2004, 55, 1831–1842. [Google Scholar] [CrossRef] [PubMed]
- Hesse, H.; Nikiforova, V.; Gakiere, B.; Hoefgen, R. Molecular Analysis and Control of Cysteine Biosynthesis: Integration of Nitrogen and Sulphur Metabolism. J. Exp. Bot. 2004, 55, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Lillo, C. Light Regulation of Nitrate Reductase in Green Leaves of Higher Plants. Physiol. Plant. 1994, 90, 616–620. [Google Scholar] [CrossRef]
- Lillo, C.; Meyer, C.; Ruoff, P. The Nitrate Reductase Circadian System. The Central Clock Dogma Contra Multiple Oscillatory Feedback Loops: Figure 1. Plant Physiol. 2001, 125, 1554–1557. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, R.F.; Campos, M.L.; Azevedo, R.A. The role of phytochromes in stress tolerance. In Salt Stress in Plants: Signalling, Omics and Adaptations; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2013; pp. 283–299. ISBN 978-1-4614-6108-1. [Google Scholar]
- Consentino, L.; Lambert, S.; Martino, C.; Jourdan, N.; Bouchet, P.-E.; Witczak, J.; Castello, P.; El-Esawi, M.; Corbineau, F.; D’Harlingue, A.; et al. Blue-Light Dependent Reactive Oxygen Species Formation by Arabidopsis Cryptochrome May Define a Novel Evolutionarily Conserved Signaling Mechanism. New Phytol. 2015, 206, 1450–1462. [Google Scholar] [CrossRef]
- Jourdan, N.; Martino, C.F.; El-Esawi, M.; Witczak, J.; Bouchet, P.-E.; D’Harlingue, A.; Ahmad, M. Blue-Light Dependent ROS Formation by Arabidopsis Cryptochrome-2 May Contribute toward Its Signaling Role. Plant Signal. Behav. 2015, 10, e1042647. [Google Scholar] [CrossRef] [Green Version]
- Gavassi, M.A.; Monteiro, C.C.; Campos, M.L.; Melo, H.C.; Carvalho, R.F. Phytochromes Are Key Regulators of Abiotic Stress Responses in Tomato. Sci. Hortic. 2017, 222, 126–135. [Google Scholar] [CrossRef] [Green Version]
- El-Esawi, M.; Arthaut, L.-D.; Jourdan, N.; D’Harlingue, A.; Link, J.; Martino, C.F.; Ahmad, M. Blue-Light Induced Biosynthesis of ROS Contributes to the Signaling Mechanism of Arabidopsis Cryptochrome. Sci. Rep. 2017, 7, 13875. [Google Scholar] [CrossRef]
- D’Amico-Damião, V.; Carvalho, R.F. Cryptochrome-Related Abiotic Stress Responses in Plants. Front. Plant Sci. 2018, 9, 1897. [Google Scholar] [CrossRef]
- Dutta Gupta, S.; Agarwal, A. Influence of LED Lighting on In Vitro Plant Regeneration and Associated Cellular Redox Balance. In Light Emitting Diodes for Agriculture; Dutta Gupta, S., Ed.; Springer: Singapore, 2017; pp. 273–303. ISBN 978-981-10-5806-6. [Google Scholar]
- Shohael, A.M.; Ali, M.B.; Yu, K.W.; Hahn, E.J.; Islam, R.; Paek, K.Y. Effect of Light on Oxidative Stress, Secondary Metabolites and Induction of Antioxidant Enzymes in Eleutherococcus Senticosus Somatic Embryos in Bioreactor. Process Biochem. 2006, 41, 1179–1185. [Google Scholar] [CrossRef]
- Baque, M.A.; Hahn, E.-J.; Paek, K.-Y. Induction Mechanism of Adventitious Root from Leaf Explants of Morinda Citrifolia as Affected by Auxin and Light Quality. In Vitro Cell. Dev. Biol. Plant 2010, 46, 71–80. [Google Scholar] [CrossRef]
- Bartoli, C.G.; Tambussi, E.A.; Diego, F.; Foyer, C.H. Control of Ascorbic Acid Synthesis and Accumulation and Glutathione by the Incident Light Red/Far Red Ratio in Phaseolus vulgaris Leaves. FEBS Lett. 2009, 583, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Cameron, J.C.; Pakrasi, H.B. Essential Role of Glutathione in Acclimation to Environmental and Redox Perturbations in the Cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 2010, 154, 1672–1685. [Google Scholar] [CrossRef] [Green Version]
- Devlin, P.F.; Kay, S.A. Cryptochromes Are Required for Phytochrome Signaling to the Circadian Clock but Not for Rhythmicity. Plant Cell 2000, 12, 2499–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanovsky, M.J.; Kay, S.A. Living by the Calendar: How Plants Know When to Flower. Nat. Rev. Mol. Cell Biol. 2003, 4, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Salomé, P.A.; Mcclung, C.R. What Makes the Arabidopsis Clock Tick on Time? A Review on Entrainment. Plant Cell Environ. 2005, 28, 21–38. [Google Scholar] [CrossRef]
- Deng, W.; Clausen, J.; Boden, S.; Oliver, S.N.; Casao, M.C.; Ford, B.; Anderssen, R.S.; Trevaskis, B. Dawn and Dusk Set States of the Circadian Oscillator in Sprouting Barley (Hordeum vulgare) Seedlings. PLoS ONE 2015, 10, e0129781. [Google Scholar] [CrossRef] [Green Version]
- Yakir, E.; Hilman, D.; Harir, Y.; Green, R.M. Regulation of Output from the Plant Circadian Clock. FEBS J. 2007, 274, 335–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, M.; Johnson, J.M.; Hieno, A.; Tokizawa, M.; Nomoto, M.; Tada, Y.; Godfrey, R.; Obokata, J.; Sherameti, I.; Yamamoto, Y.Y.; et al. High REDOX RESPONSIVE TRANSCRIPTION FACTOR1 Levels Result in Accumulation of Reactive Oxygen Species in Arabidopsis Thaliana Shoots and Roots. Mol. Plant 2015, 8, 1253–1273. [Google Scholar] [CrossRef] [Green Version]
- Leustek, T. Sulfate Metabolism. Arab. Book 2002, 1, e0017. [Google Scholar] [CrossRef]
- Lillo, C.; Appenroth, K.-J. Light Regulation of Nitrate Reductase in Higher Plants: Which Photoreceptors Are Involved? Plant Biol. 2001, 3, 455–465. [Google Scholar] [CrossRef]
- Lillo, C. Diurnal Variations of Nitrite Reductase, Glutamine Synthetase, Glutamate Synthase, Alanine Aminotransferase and Aspartate Aminotransferase in Barley Leaves. Physiol. Plant 1984, 61, 214–218. [Google Scholar] [CrossRef]
- Schupp, R.; Rennenberg, H. Diurnal Changes in the Glutathione Content of Spruce Needles (Picea abies L.). Plant Sci. 1988, 57, 113–117. [Google Scholar] [CrossRef]
- Cao, J.; Gulyás, Z.; Kalapos, B.; Boldizsár, Á.; Liu, X.; Pál, M.; Yao, Y.; Galiba, G.; Kocsy, G. Identification of a Redox-Dependent Regulatory Network of MiRNAs and Their Targets in Wheat. J. Exp. Bot. 2019, 70, 85–99. [Google Scholar] [CrossRef]
- Toldi, D.; Gyugos, M.; Darkó, É.; Szalai, G.; Gulyás, Z.; Gierczik, K.; Székely, A.; Boldizsár, Á.; Galiba, G.; Müller, M.; et al. Light Intensity and Spectrum Affect Metabolism of Glutathione and Amino Acids at Transcriptional Level. PLoS ONE 2019, 14, e0227271. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.-W.; Liu, M.-J.; Chen, I.-C.; Huang, C.-H.; Chao, L.-Y.; Hsieh, H.-L. A Glutathione S-Transferase Regulated by Light and Hormones Participates in the Modulation of Arabidopsis Seedling Development. Plant Physiol. 2010, 154, 1646–1658. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Wang, W.; Karapetyan, S.; Mwimba, M.; Marqués, J.; Buchler, N.E.; Dong, X. Redox Rhythm Reinforces the Circadian Clock to Gate Immune Response. Nature 2015, 523, 472–476. [Google Scholar] [CrossRef]
- De Montaigu, A.; Tóth, R.; Coupland, G. Plant Development Goes like Clockwork. Trends Genet. 2010, 26, 296–306. [Google Scholar] [CrossRef]
- Guilford, J.P. Creativity. Am. Psychol. 1950, 5, 444–454. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- He, H.; Van Breusegem, F.; Mhamdi, A. Redox-Dependent Control of Nuclear Transcription in Plants. J. Exp. Bot. 2018, 69, 3359–3372. [Google Scholar] [CrossRef]
- De Carvalho Oliveira, R.A.; De Andrade, A.S.; Imparato, D.O.; De Lima, J.G.S.; De Almeida, R.V.M.; Lima, J.P.M.S.; Pasquali, M.A.; De Bittencourt Pasquali, M.A.; Dalmolin, R.J.S. Analysis of Arabidopsis Thaliana Redox Gene Network Indicates Evolutionary Expansion of Class III Peroxidase in Plants. Sci. Rep. 2019, 9, 15741. [Google Scholar] [CrossRef]
- Eddy, S.R. A New Generation of Homology Search Tools Based on Probabilistic Inference. In Genome Informatics 2009, Proceedings of the 20th International Conference, Pacifico Yokohama, Japan, 14–16 December 2009; Imperial College Press: London, UK; World Scientific Publishing Co.: Yokohama, Japan, 2009; pp. 205–211. [Google Scholar]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam Protein Families Database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Gulyás, Z.; Boldizsár, Á.; Novák, A.; Szalai, G.; Pál, M.; Galiba, G.; Kocsy, G. Central Role of the Flowering Repressor ZCCT2 in the Redox Control of Freezing Tolerance and the Initial Development of Flower Primordia in Wheat. BMC Plant Biol. 2014, 14, 91. [Google Scholar] [CrossRef] [Green Version]
- Schafer, F.Q.; Buettner, G.R. Redox Environment of the Cell as Viewed through the Redox State of the Glutathione Disulfide/Glutathione Couple. Free Radic. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Morran, S.; Eini, O.; Pyvovarenko, T.; Parent, B.; Singh, R.; Ismagul, A.; Eliby, S.; Shirley, N.; Langridge, P.; Lopato, S. Improvement of stress tolerance of wheat and barley by modulation of expression of DREB/CBF factors. Plant Biotechnol. J. 2011, 9, 230–249. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | White | Far-Red | Blue | |
|---|---|---|---|---|
| Photoreceptors | HvPHYA | irregular | * <circadian | <diurnal |
| HvCRY1a | diurnal | <diurnal | =diurnal | |
| HvCRY1b | diurnal | <diurnal | =diurnal | |
| HvCRY2 | irregular | <diurnal | =diurnal | |
| Circadian clock | HvCCA1 | circadian | <diurnal | <circadian |
| HvTOC1 | circadian | <diurnal | <circadian | |
| HvPRR73 | circadian | >diurnal | >circadian | |
| Redox-responsive TFs | HvOXS2 | diurnal | =diurnal | <irregular |
| HvARF-like | circadian | <circadian | <circadian | |
| HvmTREF | diurnal | <diurnal | =diurnal | |
| Nitrate reduction | HvNR | circadian | <diurnal | <circadian |
| HvGLNa | irregular | <irregular | <irregular | |
| HvGLNb | diurnal | =diurnal | <irregular | |
| HvGLNd | diurnal | <diurnal | =no | |
| HvGLNe | no | =no | <irregular | |
| Glutathione metabolism | HvAPSR | diurnal | <irregular | <circadian |
| Hvγ-ECS | diurnal | <irregular | >irregular | |
| HvGST | circadian | >circadian | =circadian | |
| HvGR | circadian | >diurnal | <irregular | |
| Antioxidant enzymes | HvAPX1 | diurnal | >diurnal | <diurnal |
| HvAPX4 | irregulal | >irregular | <no | |
| HvCAT1 | circadian | =circadian | =circadian | |
| HvCAT2 | diurnal | >irregular | >irregular | |
| HvCAT4 | diurnal | >irregular | >diurnal | |
| HvPRX | irregular | >irregular | >irregular | |
| Thiols | Cys | irregular | <irregular | >irregular |
| CySS | irregular | > irregular | <irregular | |
| ECySS/Cys | no | >no | <no | |
| GSH | irregular | <irregular | >irregular | |
| GSSG | irregular | =irregular | <irregular | |
| EGSSG/GSH | no | >no | <no |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balogh, E.; Kalapos, B.; Ahres, M.; Boldizsár, Á.; Gierczik, K.; Gulyás, Z.; Gyugos, M.; Szalai, G.; Novák, A.; Kocsy, G. Far-Red Light Coordinates the Diurnal Changes in the Transcripts Related to Nitrate Reduction, Glutathione Metabolism and Antioxidant Enzymes in Barley. Int. J. Mol. Sci. 2022, 23, 7479. https://doi.org/10.3390/ijms23137479
Balogh E, Kalapos B, Ahres M, Boldizsár Á, Gierczik K, Gulyás Z, Gyugos M, Szalai G, Novák A, Kocsy G. Far-Red Light Coordinates the Diurnal Changes in the Transcripts Related to Nitrate Reduction, Glutathione Metabolism and Antioxidant Enzymes in Barley. International Journal of Molecular Sciences. 2022; 23(13):7479. https://doi.org/10.3390/ijms23137479
Chicago/Turabian StyleBalogh, Eszter, Balázs Kalapos, Mohamed Ahres, Ákos Boldizsár, Krisztián Gierczik, Zsolt Gulyás, Mónika Gyugos, Gabriella Szalai, Aliz Novák, and Gábor Kocsy. 2022. "Far-Red Light Coordinates the Diurnal Changes in the Transcripts Related to Nitrate Reduction, Glutathione Metabolism and Antioxidant Enzymes in Barley" International Journal of Molecular Sciences 23, no. 13: 7479. https://doi.org/10.3390/ijms23137479
APA StyleBalogh, E., Kalapos, B., Ahres, M., Boldizsár, Á., Gierczik, K., Gulyás, Z., Gyugos, M., Szalai, G., Novák, A., & Kocsy, G. (2022). Far-Red Light Coordinates the Diurnal Changes in the Transcripts Related to Nitrate Reduction, Glutathione Metabolism and Antioxidant Enzymes in Barley. International Journal of Molecular Sciences, 23(13), 7479. https://doi.org/10.3390/ijms23137479