
Soybean Meal-Dependent Acute Intestinal Inflammation Delays Osteogenesis in Zebrafish Larvae


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
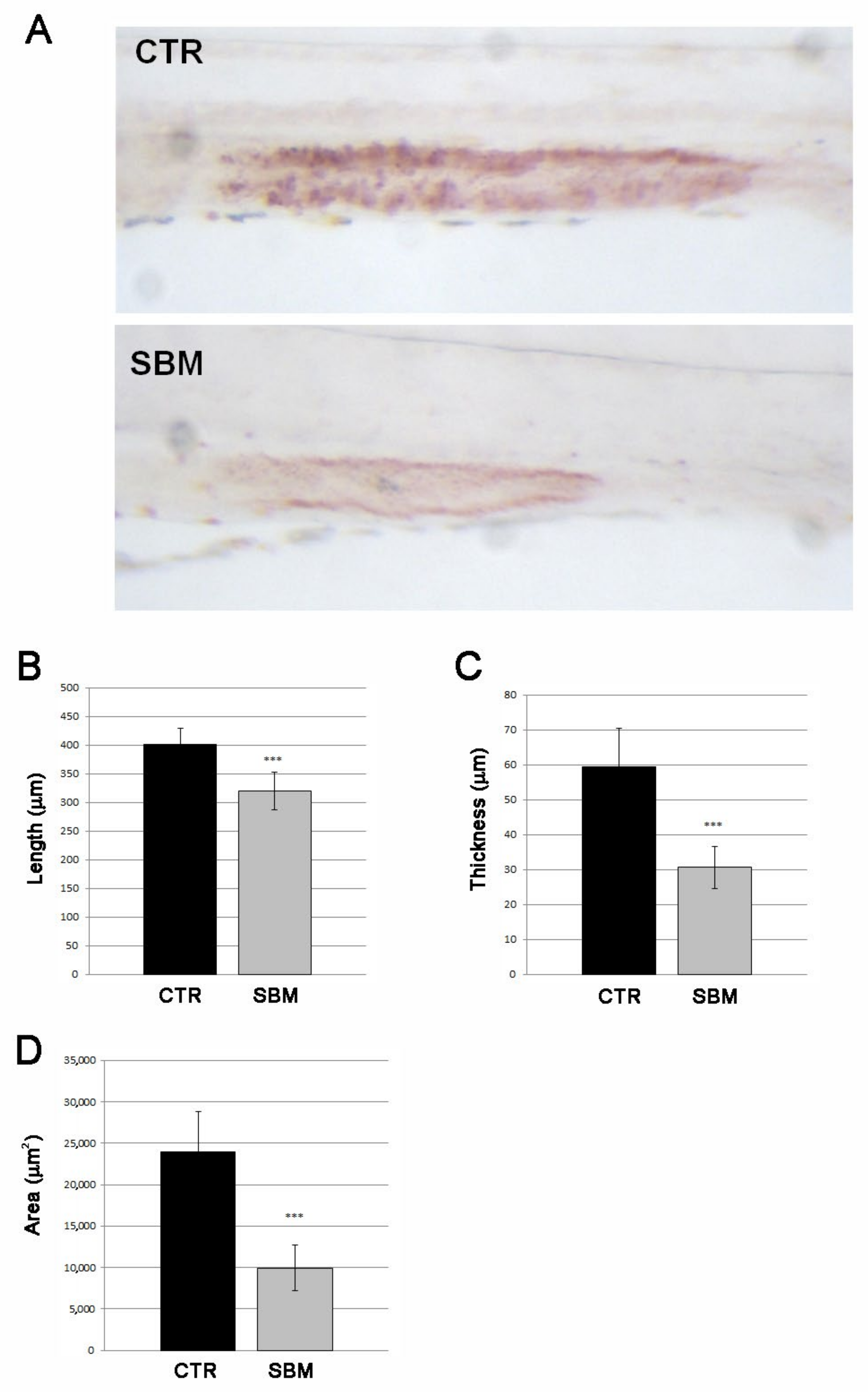
2.1. SBM-Diet Induced Alterations in Mid-Intestine of Zebrafish Larvae
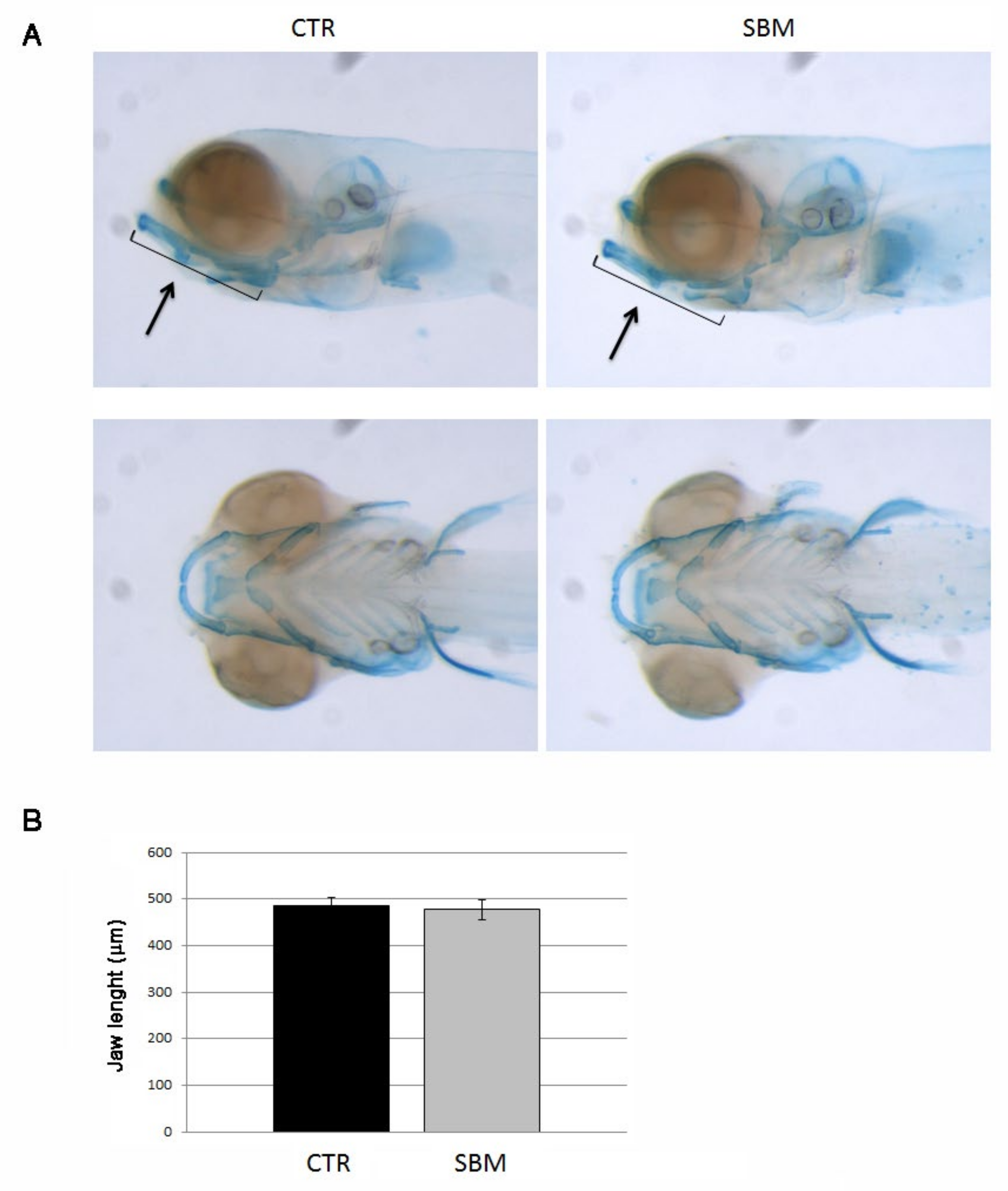
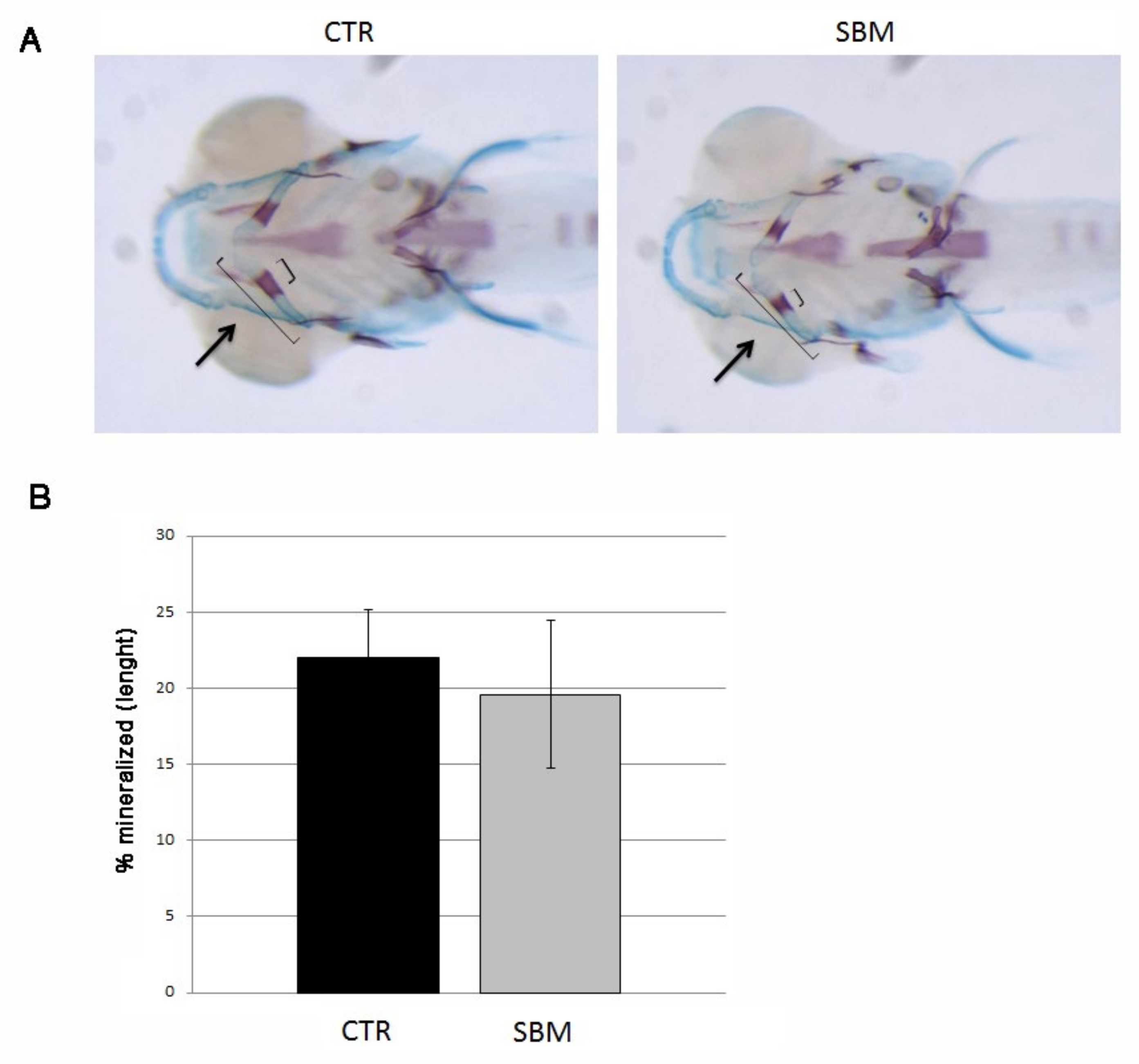
2.2. Chondrogenesis as Well as Endochondral Ossification Were Not Affected in the Head under SBM-Diet
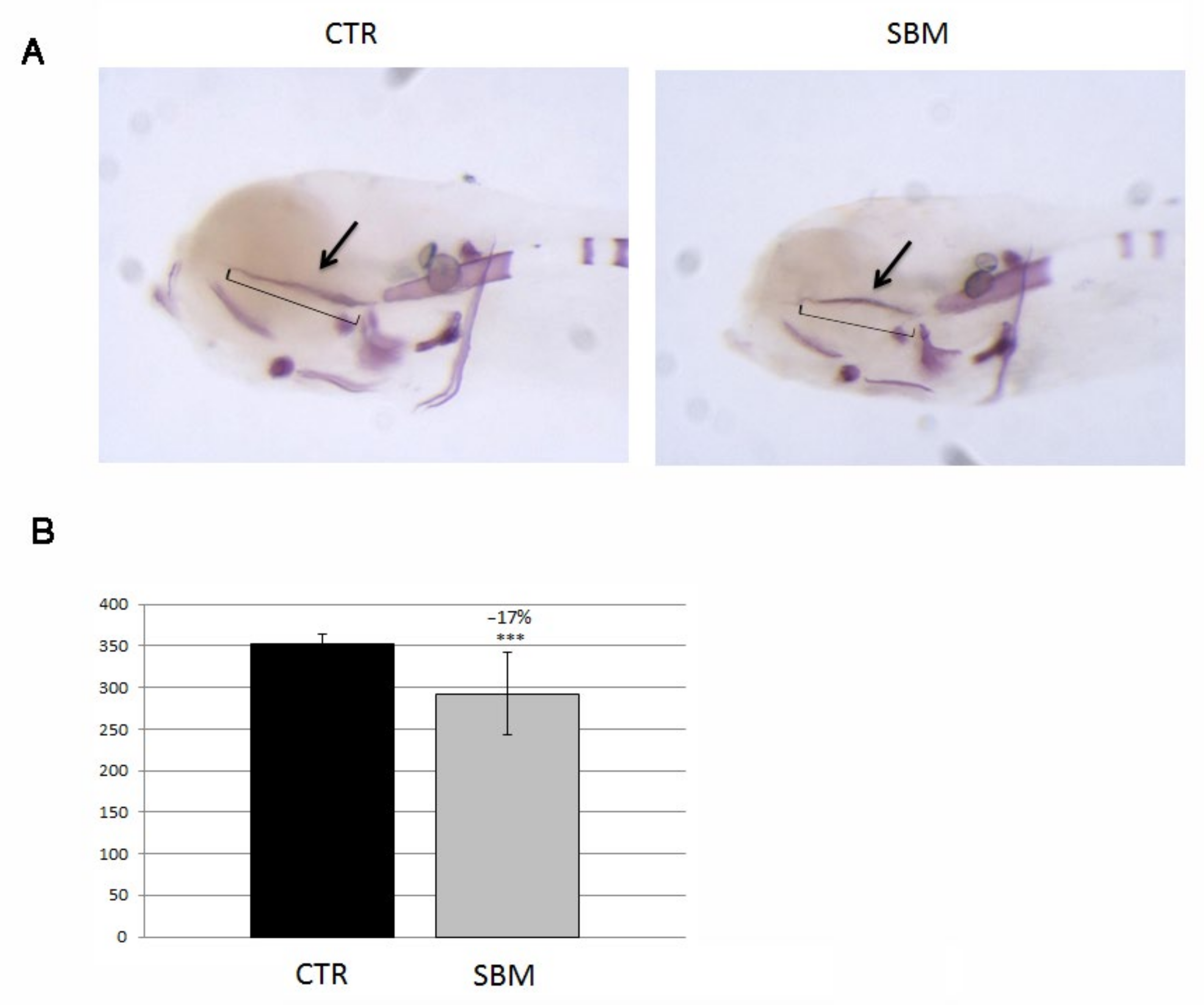
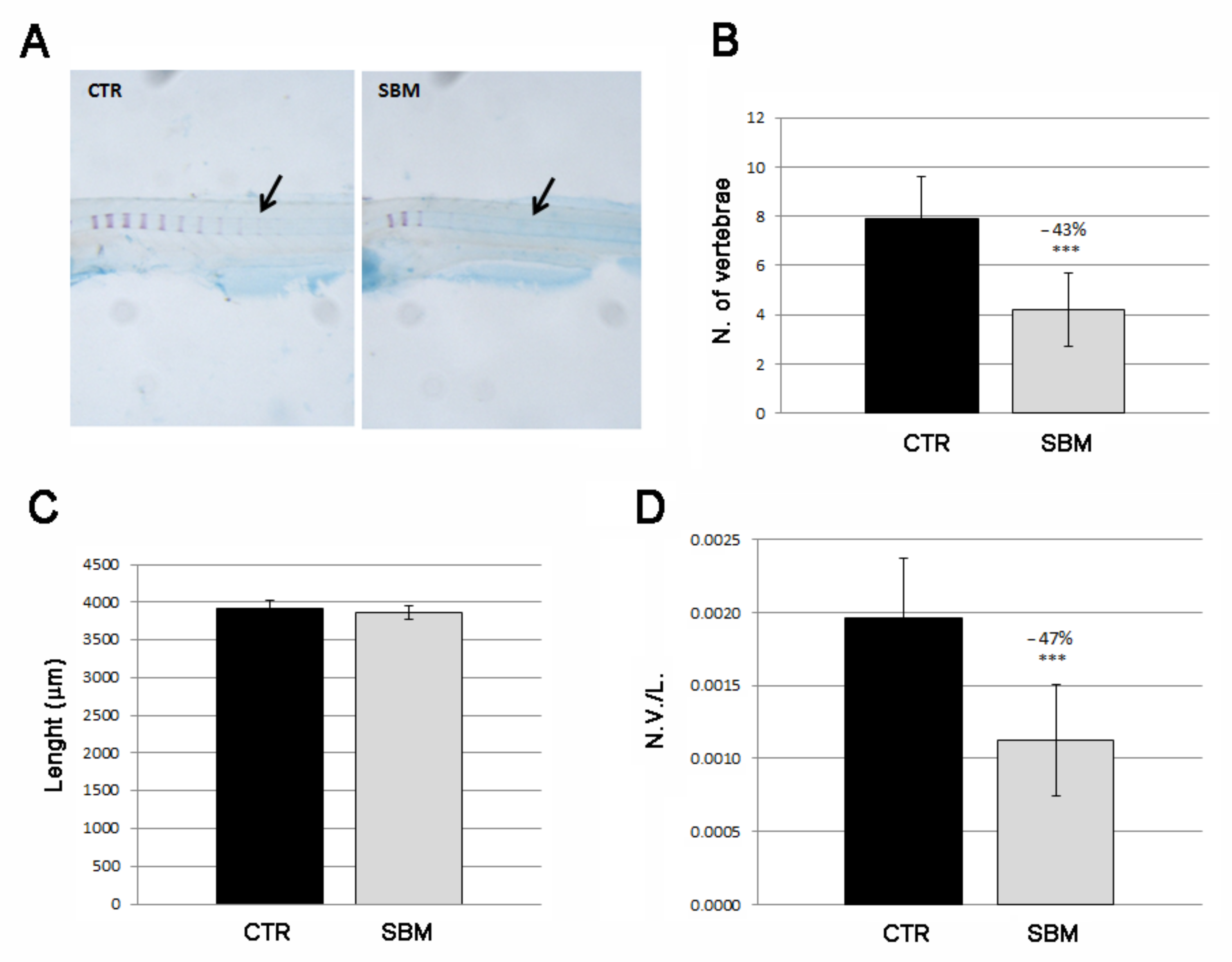
2.3. Intramembranous Ossification Was Delayed in Head and Trunk of SBM Larvae
3. Discussion
4. Materials and Methods
4.1. Ethic Statement
4.2. Animals
4.3. Treatments
4.4. Histochemical Analysis
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Deane, C.S.; Bass, J.J.; Crossland, H.; Phillips, B.E.; Atherton, P.J. Animal, Plant, Collagen and Blended Dietary Proteins: Effects on Musculoskeletal Outcomes. Nutrients 2020, 12, 2670. [Google Scholar] [CrossRef]
- Russell, L.A.; Balart, M.T.; Serrano, P.; Armstrong, D.; Pinto-Sanchez, M.I. The complexities of approaching nutrition in inflammatory bowel disease: Current recommendations and future directions. Nutr. Rev. 2022, 80, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; D’Arcy, M.; Barnes, E.L.; Freedman, N.D.; Engels, E.A.; Song, M. Associations of Inflammatory Bowel Disease and Subsequent Cancers in a Population-Based Study of Older Adults in the United States. JNCI Cancer Spectr. 2021, 6, pkab096. [Google Scholar] [CrossRef] [PubMed]
- Irwin, R.; Lee, T.; Young, V.B.; Parameswaran, N.; McCabe, L.R. Colitis induced bone loss is gender dependent and associated with increased inflammation. Inflamm. Bowel Dis. 2013, 19, 1586–1597. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.J.; Li, H.; Wu, D.T.; Zhuang, Q.G.; Li, H.B.; Geng, F.; Gan, R.Y. Recent development in zebrafish model for bioactivity and safety evaluation of natural products. Crit. Rev. Food Sci. Nutr. 2021, 1, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Tavares, B.; Santos Lopes, S. The importance of Zebrafish in biomedical research. Acta Médica Portuguesa 2013, 26, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Stoddard, M.; Huang, C.; Enyedi, B.; Niethammer, P. Live imaging of leukocyte recruitment in a zebrafish model of chemical liver injury. Sci. Rep. 2019, 9, 28. [Google Scholar] [CrossRef]
- Kimmel, C.B.; DeLaurier, A.; Ullman, B.; Down, J.; McFadden, M. Modes of developmental outgrowth and shaping of a craniofacial bone in zebrafish. PLoS ONE 2010, 5, e9475. [Google Scholar] [CrossRef] [Green Version]
- Bird, N.C.; Mabee, P.M. Developmental morphology of the axial skeleton of the zebrafish, Danio rerio (Ostariophysi: Cyprinidae). Dev. Dyn. 2003, 228, 337–357. [Google Scholar] [CrossRef]
- Bensimon-Brito, A.; Cardeira, J.; Cancela, M.L.; Huysseune, A.; Witten, P.E. Distinct patterns of notochord mineralization in zebrafish coincide with the localization of Osteocalcin isoform 1 during early vertebral centra formation. BMC Dev. Biol. 2012, 12, 28. [Google Scholar] [CrossRef] [Green Version]
- Brugman, S. The zebrafish as a model to study intestinal inflammation. Dev. Comp. Immunol. 2016, 64, 82–92. [Google Scholar] [CrossRef] [Green Version]
- López Nadal, A.; Peggs, D.; Wiegertjes, G.F.; Brugman, S. Exposure to Antibiotics Affects Saponin Immersion-Induced Immune Stimulation and Shift in Microbial Composition in Zebrafish Larvae. Front. Microbiol. 2018, 9, 2588. [Google Scholar] [CrossRef]
- Oehlers, S.H.; Flores, M.V.; Okuda, K.S.; Hall, C.J.; Crosier, K.E.; Crosier, P.S. A chemical enterocolitis model in zebrafish larvae that is dependent on microbiota and responsive to pharmacological agents. Dev. Dyn. 2011, 240, 288–298. [Google Scholar] [CrossRef]
- Francis, G.; Kerem, Z.; Makkar, H.P.S.; Becker, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef]
- Kumar, V.; Hossain, M.S.; Ragaza, J.A.; Rubio Benito, M. The potential impacts of soy protein on fish gut health. In Soybean for Human Consumption and Animal Feed; IntechOpen: London, UK, 2020; Chapter 5. [Google Scholar] [CrossRef]
- Baeverfjord, G.; Krogdahl, A. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L.; distal intestine: A comparison with the intestines of fasted fish. J. Fish Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Krogdahl, Å.; Bakke-Mckellep, A.; Baeverfjiord, G. Effects of graded levels of standard soybean meal on intestinal structure, mucosal enzyme activities, and pancreatic response in Atlantic salmon (Salmon salar L.). Aquac. Nutr. 2003, 9, 361–371. [Google Scholar] [CrossRef]
- Urán, P.A.; Gonçalves, A.A.; Taverne-Thiele, J.J.; Schrama, J.W.; Verreth, J.A.; Rombout, J.H. Soybean meal induces intestinal inflammation in common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 2008, 25, 751–760. [Google Scholar] [CrossRef]
- Hedrera, M.I.; Galdames, J.A.; Jimenez-Reyes, M.F.; Reyes, A.E.; Avendaño-Herrera, R.; Romero, J.; Feijóo, C.G. Soybean meal induces intestinal inflammation in zebrafish larvae. PLoS ONE 2013, 8, e69983. [Google Scholar] [CrossRef] [Green Version]
- Carnovali, M.; Valli, R.; Banfi, G.; Porta, G.; Mariotti, M. Soybean Meal-Dependent Intestinal Inflammation Induces Different Patterns of Bone-Loss in Adult Zebrafish Scale. Biomedicines 2021, 9, 393. [Google Scholar] [CrossRef]
- Wallace, K.N.; Akhter, S.; Smith, E.M.; Lorent, K.; Pack, M. Intestinal growth and differentiation in zebrafish. Mech. Dev. 2005, 122, 157–173. [Google Scholar] [CrossRef]
- Chuang, L.S.; Morrison, J.; Hsu, N.; Labrias, P.R.; Nayar, S.; Chen, E.; Villaverde, N.; Facey, J.A.; Boschetti, G.; Giri, M.; et al. Zebrafish modeling of intestinal injury, bacterial exposures and medications defines epithelial in vivo responses relevant to human inflammatory bowel disease. Dis. Model Mech. 2019, 12, dmm037432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs) -Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walldorf, J.; Krummenerl, A.; Engler, K.; Busch, J.; Dollinger, M.M.; Seufferlein, T.; Albert, J.G. Health care for osteoporosis in inflammatory bowel disease: Unmet needs in care of male patients? J. Crohn’s Colitis 2013, 7, 901–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulte, C.M.; Dignass, A.U.; Goebell, H.; Röher, H.D.; Schulte, K.M. Genetic factors determine extent of bone loss in inflammatory bowel disease. Gastroenterology 2000, 119, m909–m920. [Google Scholar] [CrossRef]
- Shannon, H.C.; Ravid, K. Differentiation of mesenchymal stem cells to osteoblasts and chondrocytes: A focus on adenosine receptors. Expert Rev. Mol. Med. 2013, 15, e1. [Google Scholar]
- Gilbert, L.; He, X.; Farmer, P.; Boden, S.; Kozlowski, M.; Rubin, J.; Nanes, M.S. Inhibition of osteoblast differentiation by tumor necrosis factor-alpha. Endocrinology 2000, 141, 3956–3964. [Google Scholar] [CrossRef]
- Gilbert, L.; He, X.; Farmer, P.; Rubin, J.; Drissi, H.; van Wijnen, A.J.; Lian, J.B.; Stein, G.S.; Nanes, M.S. Expression of the osteoblast differentiation factor RUNX2 (Cbfa1/AML3/Pebp2alpha A) is inhibited by tumor necrosis factor-alpha. J. Biol. Chem. 2002, 277, 2695–2701. [Google Scholar] [CrossRef] [Green Version]
- McCabe, L.; Britton, R.A.; Parameswaran, N. Prebiotic and probiotic regulation of bone health: Role of the intestine and its microbiome. Curr. Osteoporos. Rep. 2015, 13, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Avella, M.A.; Place, A.; Du, S.J.; Williams, E.; Silvi, S.; Zohar, Y.; Carnevali, O. Lactobacillus rhamnosus accelerates zebrafish backbone calcification and gonadal differentiation through effects on the GnRH and IGF systems. PLoS ONE 2012, 7, e45572. [Google Scholar] [CrossRef]
- Carnevali, O.; Avella, M.A.; Gioacchini, G. General and comparative endocrinology. Gen. Comp. Endocrinol. 2013, 188, 297–302. [Google Scholar] [CrossRef]
- Maradonna, F.; Gioacchini, G.; Falcinelli, S.; Bertotto, D.; Radaelli, G.; Olivotto, I.; Carnevali, O. Probiotic supplementation promotes calcification in Danio rerio larvae: A molecular study. PLoS ONE 2013, 8, e83155. [Google Scholar] [CrossRef] [Green Version]
- Oehlers, S.H.; Flores, M.V.; Hall, C.J.; Okuda, K.S.; Sison, J.O.; Crosier, K.E.; Crosier, P.S. Chemically induced intestinal damage models in zebrafish larvae. Zebrafish 2013, 10, 184–193. [Google Scholar] [CrossRef]
- Bakke-McKellep, A.M.; Penn, M.H.; Salas, P.M. Effects of dietary soyabean meal, inulin and oxytetracycline on intestinal microbiota and epithelial cell stress, apoptosis and proliferation in the teleost Atlantic salmon (Salmo salar L.). Br. J. Nutr. 2007, 97, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Merrifield, D.L.; Dimitroglou, A.; Bradley, G.; Baker, R.T.; Davies, S.J. Soybean meal alters autochthonous microbial populations, microvilli morphology and compromises intestinal enterocyte integrity of rainbow trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis. 2009, 32, 755–766. [Google Scholar] [CrossRef]
- Westerfield, M. The zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio), 5th ed.; University of Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Herbomel, P.; Thisse, B.; Thisse, C. Zebrafish early macrophages colonize cephalic mesenchyme and developing brain, retina, and epidermis through a M-CSF receptor-dependent invasive process. Dev. Biol. 2001, 238, 274–288. [Google Scholar] [CrossRef] [Green Version]
- Matthews, M.; Varga, Z.M. Anesthesia and euthanasia in zebrafish. ILAR J. 2012, 53, 192–204. [Google Scholar] [CrossRef] [Green Version]
- Walker, M.B.; Kimmel, C.B. A two-color acid-free cartilage and bone stain for zebrafish larvae. Biotech. Histochem. 2007, 82, 23–28. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carnovali, M.; Banfi, G.; Porta, G.; Mariotti, M. Soybean Meal-Dependent Acute Intestinal Inflammation Delays Osteogenesis in Zebrafish Larvae. Int. J. Mol. Sci. 2022, 23, 7480. https://doi.org/10.3390/ijms23137480
Carnovali M, Banfi G, Porta G, Mariotti M. Soybean Meal-Dependent Acute Intestinal Inflammation Delays Osteogenesis in Zebrafish Larvae. International Journal of Molecular Sciences. 2022; 23(13):7480. https://doi.org/10.3390/ijms23137480
Chicago/Turabian StyleCarnovali, Marta, Giuseppe Banfi, Giovanni Porta, and Massimo Mariotti. 2022. "Soybean Meal-Dependent Acute Intestinal Inflammation Delays Osteogenesis in Zebrafish Larvae" International Journal of Molecular Sciences 23, no. 13: 7480. https://doi.org/10.3390/ijms23137480
APA StyleCarnovali, M., Banfi, G., Porta, G., & Mariotti, M. (2022). Soybean Meal-Dependent Acute Intestinal Inflammation Delays Osteogenesis in Zebrafish Larvae. International Journal of Molecular Sciences, 23(13), 7480. https://doi.org/10.3390/ijms23137480